![]()
Previous--Introduction | Next--Relationships
Terminology of Lophophyllid Structures
Several attempts have been made to standardize the terminology of coral structures (Grabau, 1922; Hill, 1935; Sanford, 1939). Each of these studies is somewhat different from the others, and none has gained general acceptance. The terminology of rugose coral structures that has been proposed in these papers is highly technical and includes a considerable number of precisely defined terms. This precision in the definition of terms aids in the conciseness of descriptions, but the use of little known, complex technical names tends to make understanding of descriptions very difficult for those not specialized in this branch of invertebrate paleontology. A review of several recent papers on American rugose corals indicates no definite agreement as to terminology. Therefore, the somewhat simpler and less concise system of terminology employed by Moore and Jeffords (1941) is adopted. The nomenclature of structures found in lophophyllid corals is here described as an aid to the understanding of the remainder of the paper.
The skeleton secreted by a single individual is known as the corallite. In skeletons formed by one individual only, this is synonymous with the corallum, which is the entire skeletal mass, whether composed of one or many corallites. Rugose corals having a corallum formed by only one corallite have commonly been called "simple," but unless one is familiar with this usage, it may seem to indicate a lack of specialization in the corallite rather than the hard parts of a single individual. The term solitary seems preferable. The initial point at the small end of the corallite is the apex, and it is at the lowermost part of the corallite. The last-formed part is termed the upper portion. The cup-shaped depression at the upper part of the corallite is the calyx. It may contain a raised area, sometimes termed a boss, or a depression, sometimes designated as a pit. The external portion of the corallite is designated the peripheral part and the central or innermost part the axial area. Proximal and distal have been used by some authors in a sense equivalent to peripheral and axial, respectively, but others have designated the lower portion of the corallite as proximal and the upper portion as distal.
The form or shape of solitary corallites may vary from nearly flat discs to long cylinders. Hill (1935, p. 488) has proposed a number of terms based on the rate of expansion of the conical lower part of the corallite, as shown by the apical angle. The shape of the corallite of many rugose corals varies somewhat during growth, and it may vary among individuals. The terms conical and cylindrical, or combinations of these with common adjectives, seem sufficient to characterize the shape, at least in the case of the lophophyllid corals here described.
Study of sectioned material shows that superficial similarity of external features offers extremely little chance for accurate separation of these corals. After internal features of corals from any one locality are known, it is possible to distinguish the lophophyllid corals accurately from other genera and usually to recognize the different species. One cannot make reliable determinations, however, by applying the same criteria to corals from other localities or horizons. Some species of "Zaphrenthis," Canina, "Axophyllum," "Amplexus," Aulophyllum?, and other unidentified genera cannot be separated from lophophyllid corals without knowledge of internal structures, shown either in the calyx or in transverse section. It may be noted that the original spelling of the name commonly written Zaphrentis is as given here (Zaphrenthis) and emendation in dropping the second "h" is incorrect. Typically the corallite in the case of the lophophyllid corals continues to increase in diameter or remains nearly constant as it develops. Some specimens, however, show abrupt constrictions that are followed by expansion like that near the apex. This is known as rejuvenescence.
The outer wall of the corallite was presumed by early workers to be a layer deposited upon the original wall or theca. In well preserved specimens of the Rugosa it has been found that this outer layer, or "epitheca," extends to the highest part of the calyx, showing that it is the first formed, fundamental unit of the wall (Grabau, 1922, p. 4) and not a layer superposed on an inner wall. This single external wall of the corallite is here designated as the theca (Sanford, 1939, p. 305).
The outer surface of the theca may be smooth, or it may show longitudinal grooves and ridges. In the Rugosa these grooves correspond in position to the septa and are called septal grooves. The ridges, known as interseptal ridges, are formed by the bulging out of the polyp wall between the septa. In the Hexacoralla the longitudinal ridges are extensions of the septa through the wall, and the grooves correspondingly indicate interseptal positions.
The nature of the septal grooves and interseptal ridges may be of value in separating types of lophophyllid corals not otherwise separable on external features. For example, each of the Lower Permian species, Malonophyllum kansasense, Lophamplexus eliasi, and Lophophyllidium dunbari, described by Moore and Jeffords (1941) from the Florena shale member of the Beattie limestone at Grand Summit, Cowley County, Kansas, seems to have distinctive features of the septal grooves and ridges.
Transverse ornamentation may consist of fine growth lines or alternate expansions and broad constrictions called wrinkles. Some species have small tubes or rootlets projecting from the theca. These are termed radicles. A number of very different types of corals have these radicles and they seem to be a parallel development that is shown by species belonging to a number of different genera. The radicles may form only during the youthful stages and be restricted to the lower part of the corallite, or they may be formed throughout growth of the polyp.
The different classes of septa have been given several names. The terms primary, secondary, and tertiary septa vary in meaning according to the author using them. The designation of the septa as given by Hill (1935, p. 504-505) seems to have the advantage of both clarity and consistency. The cardinal, counter, and two alar septa are termed the protosepta. A variable number of other septa that are added in pairs in the cardinal and counter quadrants and that closely resemble the protosepta, are designated as metasepta (plate 6, figures 1b, c). The protosepta and metasepta together are termed the major septa. Short septa alternating with the major septa, introduced between the majors, are termed the minor septa (plate 5, figure 2b).
The manner of introduction of the septa in the Rugosa is well known and needs little description here. The first septa introduced are the counter and cardinal septa and indicate the plane about which most rugose corals are bilaterally symmetrical. Subsequently, or more rarely at the same time as the other protosepta are formed, two lateral septa known as the alar septa are introduced. These divided the corallite into four quadrants, termed the counter and the cardinal quadrants, according to whether they are separated by the counter or the cardinal septum (plate 7, figure 2b). The rest of the major septa are inserted alternately in pairs in the counter quadrants and cardinal quadrants throughout growth. In the cardinal quadrants metasepta are inserted on each side of the cardinal septum. Those inserted in the counter quadrants develop on the counter side of the alar septa. The most recently inserted septa tend to be shorter than the earlier ones and to be inclined away from the point of development (plate 2, figure 3b).
The European students of rugose corals (Duerden, 1903, p. 389, 1906, p. 226, 1906a; Carruthers, 1906, p. 356; Faurot, 1909; Smith, 1916, p. 60; Gerth, 1919; Schindewolf, 1930; Hill, 1935, p. 504) also include as protosepta a third pair of septa, which are called counter laterals. Edwards and Haime (1848), Kunth (1869), Gordon (1906), Brown (1909, p. 51), Grabau (1922, p. 10), Weissermel (1927), and Sanford (1939, p. 306) consider only the first four septa as protosepta. Hudson (1935a, p. 3) designates only the cardinal and counter septa as protosepta. In the lophophyllid corals that I have examined, the counter lateral septa are not distinctive, aside from their position and early appearance. The distinct acceleration of the counter quadrants may cause the insertion of the counter lateral septa almost simultaneously with the alar septa. The counter-cardinal plane is an important morphologic feature throughout growth, inasmuch as one of each pair of the metasepta is inserted on each side of this plane. The metasepta of the counter quadrants are inserted adjacent to the alar septa. The counter laterals, on the other hand, have little relation to the further development of the corallite, and so do not seem to be worthy of inclusion with the other distinctive septa as protosepta.
In corallites having distinct septal grooves, it is possible to determine the position of the protosepta from study of the exterior. The cardinal septum is represented by a septal groove running the entire length of the corallite. The insertion of the pairs of metasepta, one on each side, is shown by the pinnate arrangement of their septal grooves. Inasmuch as no metasepta are added adjacent to the counter septum, the metaseptal grooves are parallel to its groove. The alar septa are indicated by the parallel grooves on the cardinal side, with grooves on the counter side converging toward them.
The septal groove pattern of these lophophyllid corals is similar to that described by Vojnovskij-Krieger (1928) for the rugose corals. The grooves do not have a simple pinnate junction with the cardinal septum or the counter side of the alar septa, as shown by Grabau (1922, p. 12) and Sanford (1939, fig. 1). The traces of the alar, cardinal and counter septa extend from the apex to the calyx and are not connected to other grooves. Metasepta inserted in the cardinal quadrants start in the spaces next to the cardinal septum, and grooves are formed composed of the early parts of each of the metasepta of the cardinal quadrants. Similar grooves develop on the counter side of the alar septa (fig. 1a-d). Thus the traces of the septa on the exterior may be divided into four similar quadrants.
Figure 1. Diagrammatic sketches of the septal groove pattern of rugose corals. A, Septal groove pattern in the cardinal quadrants. B, Septal groove pattern in the counter quadrants. C, Septal groove pattern of the cardinal and counter quadrants seen from the side. D, Sketch of the relationships of septal grooves to septa, fossula, and pseudofossulae. The position of the pseudofossulae is indicated by the stippled areas.
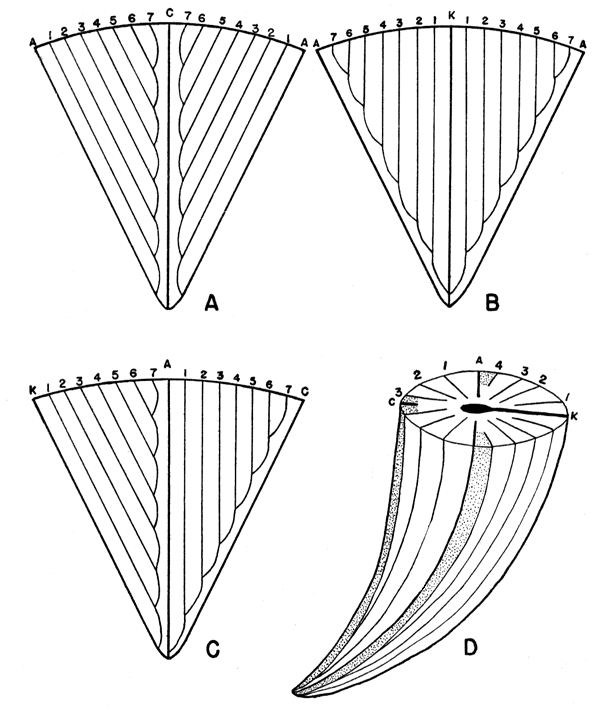
The metasepta may be introduced equally in each of the quadrants, but many species are characterized by a more rapid introduction of the septa in the counter quadrant or in the cardinal quadrant. This advance in rate of increase of septa in one pair of quadrants is called acceleration (plate 7, figure 2b). Minor septa may be inserted between the major septa either equally or unequally in the four quadrants. They usually are present only in the later stages of development.
Long septa are those that reach almost to the axis. Short septa extend only a slight distance from the periphery. The minor septa are nearly always much shorter than the major septa. In some forms the major septa withdraw from the axis, and the septa of such types have been called amplexoid septa. Hill (1935, p. 502) points out that this term was proposed by Carruthers (1910, p. 525) for septa that are fully developed only on the upper surfaces of the tabulae. A preferable term for axially shortened major septa, therefore, is brevisepta.
Depressions in the floor of the calyx, as shown in the corallite, are termed fossulae. These are formed about one or more of the septa that is shortened or aborted. In the lophophyllid corals it is the cardinal septum that is shortened, forming a cardinal fossula (plate 7, figures 1b, c). The spaces left on the counter side of the alar septa, due to the poorly developed last septa and to their tendency to lean towards the counter septum, are called alar pseudofossulae (plate 7, figures 2b, c). They differ from a fossula in that they are produced by a retardation in development and leaning of the septa, rather than by a structural shortening of a septum and down-sagging of the base of the polyp in these regions. Similar pseudofossulae on either side of the cardinal septum are grouped with the cardinal fossula (fig. 1d).
The main transverse skeletal elements of the corallite that are not limited by the septa are called tabulae. They are formed by the polyp in lifting itself upward in the calyx, abandoning the underlying part of the corallite. Tabulae may have a great number of different forms and characters. Complete tabulae (Hill, 1935, p. 511; not Sanford, 1939, p. 309) are those that extend across the axial area, without intersecting other tabulae or an axial column, and that are joined peripherally either to the wall or to dissepiments. Other tabulae are incomplete.
Small plates arched convexly upwards between the septa and peripheral in position are known as dissepiments. The nature and distribution of dissepiments indicate that the polyp could lift itself up in the calyx gradually rather than all at one time as in the formation of tabulae.
In transverse sections dissepiments and some tabulae are indicated by curved bars between the septa (fig. 2). Dissepiments develop only between the septa, and accordingly bars on opposite sides of a single septum are not parts of a single dissepiment. Tabulae extend across nonseptate parts of a coral, and where they intersect the septa they are typically not confined by the septa, as seen in transverse sections. Thus they appear as oblique bars crossing several septa. The tabulae may be slightly lower or higher on different sides of a septum. In transverse section they may show only as an occasional band between septa (plate 3, figure 1b), or slightly offset (plate 2, figure 3b). They also may curve towards the periphery adjacent to a septum (plate 5, figure 2b) and in so doing may seem not to be continuous on opposite sides of the septum.
Figure 2. Sections of Caninia sp. showing tabulae and dissepiments in transverse and longitudinal section. The dissepiments occur in a narrow peripheral zone, and slightly arched tabulae cross the interior of the corallite.
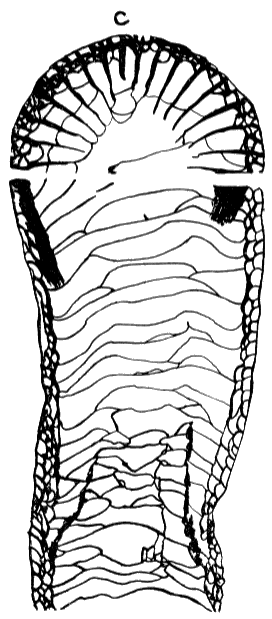
Several recent descriptions of rugose corals reveal a tendency to interpret as dissepiments some of these intercepts of irregular tabulae in transverse sections. Inasmuch as tabulae and dissepiments are distinct structures and important features in the description of rugose corals, uniform interpretation and nomenclature of these structures is considered necessary.
The details of the axial structures seem very significant in correct generic placement of rugose corals. Detailed classifications of the axial structures have been proposed by Grabau (1922) and by Hill (1935), but there is little similarity in the terminology. These and others who have described axial structures of the Rugosa use the terms columella and pseudocolumella, with contradictory meaning, to describe varying types of axial structures. The classification proposed by Hill is perhaps best adapted for use in describing axial structures in corals similar to Lonsdaleia or Clisiophyllum. Grabau describes a number of different kinds of axial structures, especially the solid, rodlike type, and gives descriptive names to each type. It is difficult or impossible to describe all of the characteristic types of axial structures by separate names without the use of many unfamiliar technical terms. Inasmuch as the names now used in description of axial structures have no standardized meaning or are unnecessarily technical, Moore and Jeffords (1941) have termed any axial structure of a corallite an axial column, regardless of its appearance or manner of formation. The features of the column of each of these forms must then be described in detail. The significance of many features seen in axial columns of the corals studied is not clearly known.
Short longitudinal plates in the axial region are known as lamellae. A lamella in the counter cardinal plane is termed the median lamella and others lateral in position are called radiating lamellae.
Transverse sections of some rugose corals show the presence of an inner wall. This wall is concentric with the exterior of the corallite and may be formed in several ways (Grabau, 1922). When present in lophophillid corals, this is most often produced by deposition of stereoplasm around the septa and other skeletal elements. In some forms ("Sinophyllum" Grabau, 1928) the inner wall is reported to be formed by the sudden bending of the axial ends of the septa so as to join with each other. A consistant spacing of a change in the arching of tabulae may also produce an inner wall. The swollen axial ends of the septa supplemented by deposition of stereoplasm about this thickened part forms definite walls in other lophophyllid corals.
Greek (1936) and others compare coral species by means of transverse sections at similar diameters. The size of the corallite in many genera, especially in Caninia, is rather variable. This method of comparison, therefore, seems of little value for comparing many of the Rugosa. Study of lophophyllid coral structures suggests comparison of equivalent stages in development regardless of size. Differences in equivalent periods of growth and in extent of development can then be determined easily.
The peculiar features of some rugose corals as shown by Lang (1923) are common to many widely varying types. One genetic line may show any number of these different tendencies superimposed on the original genetic structural features. The astraeoid trend (1) is defined as the development from solitary to colonial forms having loosely bundled corallites and finally to closely packed types. The caninioid trend (2) is marked by change in shape of the corallite from discord through conical to cylindrical. A cyathophylloid trend (3) is the tendency of a corallite to go from its primary bilateral symmetry to radial symmetry. The amplexoid trend (4) is distinguished by a tendency of the septa to become shorter (breviseptal) and to withdraw from the axis. The lonsdaleoid trend (5) is expressed by a shortening of the septa at their peripheral ends, accompanied by the presence of a peripheral dissepimental zone. The cystiphylloid trend (6) is the uplifting of the polyp in the calyx very gradually by dissepiments instead of periodically by tabulae. The aulophylloid trend (7) is the tendency for an axial column to become more highly developed and complex as a genetic line progresses. The petraeoid trend (8) is represented by the development of secondary thickening of the skeletal elements in a definite manner.
The conspicuous development of like trends in several distinct generic lines may produce very similar forms, known as homeomorphs. These are forms that show a similarity of structures but a different ancestry. Hudson (1935) considers the very similar genera Rhopalolasma Hudson (1935) and Tachylasma Grabau (1922) to be merely homeomorphic. Likewise Weissermel (1897), Schindewolf (1940), and Moore and Jeffords (1941) have shown that several different types of rugose corals have a prominent development of the amplexoid trend, or breviseptal phase, in maturity. Several of these corals that were grouped formerly in the long ranging genus "Amplexus" have now been separated into genera that are useful stratigraphic markers. As yet, little work has been done on the significance and extent of homeomorphism in the external and internal elements of the Rugosa.
Previous--Introduction | Next--Relationships
Kansas Geological Survey, Geology
Placed on web September 2005; originally published November 30, 1942.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/41_5/03_term.html