![]()
Prev Page--Paleoenvironments || Next Page--Structural Features, Conclusions
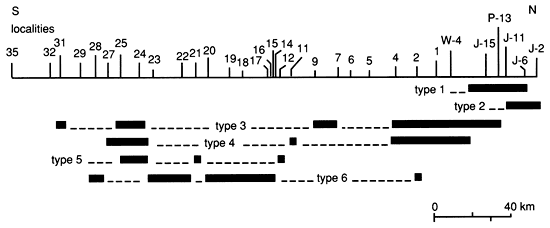
Lateral distribution of paleoenvironments of the lower Americus Limestone Member
Paleoenvironments changed laterally within the lower part of the Americus Limestone Member, as evidenced by changes in thickness of shale, lithologic texture, morphology of boundstones, and fossils. Differentiation of paleoenvironments is largely the result of differences in the paleotopography of the seafloor, which causes differences in depositional conditions. Anticlinal structures in the subsurface are located at the sites of paleotopographic highs, indicating that the paleotopography and the differentiation of paleoenvironments were governed by structural features.
Thickness of shale as an indicator of paleoenvironment
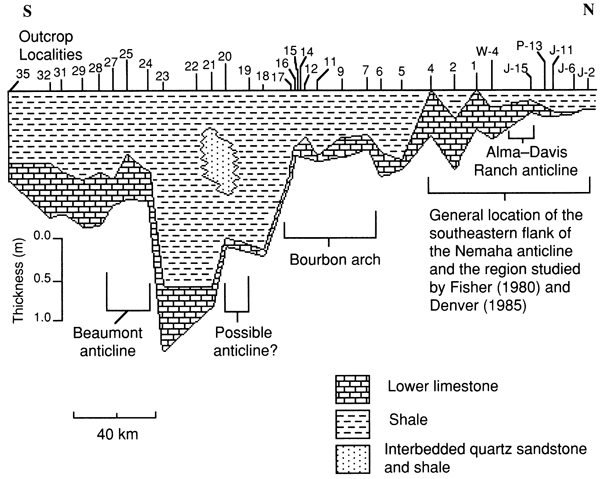
In general, topographically low areas with relatively deep water were the sites of low-energy deposition, which favored the accumulation of thick deposits of terrigenous mud. Topographically high areas, the sites of relatively shallow-water and somewhat higher-energy deposition or subaerial exposure, subsequently received thinner deposits of terrigenous mud. Figure 21 shows in cross section four paleotopographically high areas of seafloor as delineated by relatively thinner intervals of shale. Each area is interpreted as having been the site of relatively shallow-water deposition at some time during transgression. The exaggeration of the shale thickness that results from differential compaction between shale and interbedded quartz sandstone at localities (outcrops) 21 and 22 is negligible because correction for it does not alter the location of thick and thin intervals of shale. It can be argued that thick intervals of shale are delta lobes that prograded with little or no relationship to topography. This is unlikely because, as will be shown, lithologic texture and the morphology of boundstone in the lower limestone bed indicate that areas covered by thin shales were indeed deposited under conditions of relatively higher energy and that areas covered by thick shales were deposited under conditions of relatively low energy. The reasonable explanation for these relationships is that topography controlled depositional patterns.
Figure 21--Cross section of the lower part of the Americus Limestone Member from Jackson County to Cowley County, Kansas, showing sample localities and known locations of structural features. Areas of relatively thin shale are the probable locations of paleotopographically high seafloors [see Jewett and Merriam (1959) and Fisher (1980); see also fig. 43]. Datum is base of limestone above lowermost limestone.
Lithologic texture as an indicator of paleoenvironment
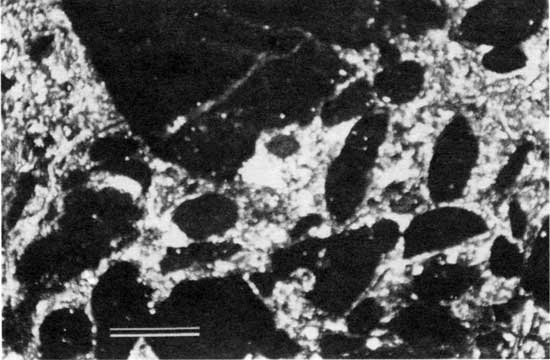
In general, carbonate rocks that contain less mud and are grain supported, such as grainstones and packstones, are believed to have been deposited under higher energy conditions than those that contain more mud and lack grain support, such as wackestones and mudstones (Dunham, 1962; see appendix A). Figure 22 illustrates the lateral distribution of the various rock types of the lower limestone bed, and fig. 23 shows the locations of relatively low- and high-energy deposits as indicated by lithologic texture. As expected, each of the four locations regarded as the probable site of a shallow-water paleoenvironment contains or is closely flanked by rocks with less mud, indicating deposition under relatively high energy. Muddier rocks, on the other hand, occur primarily at localities associated with thick overlying intervals of shale, indicating that these areas had low-energy depositional environments. The conglomeratic lime-foraminifer-ostracode wackestone to packstone (fig. 24), which is limited to localities 29 and 32, may have resulted from erosion of nearby paleotopographically high areas during storms.
Figure 22--Schematic cross section of the lower limestone bed of the Americus Limestone Member showing distribution of lithofacies and locations of structural features. Datum is the base of the limestone above the lowermost limestone of the Americus. [See Jewett and Merriam (1959) and Fisher (1980).]
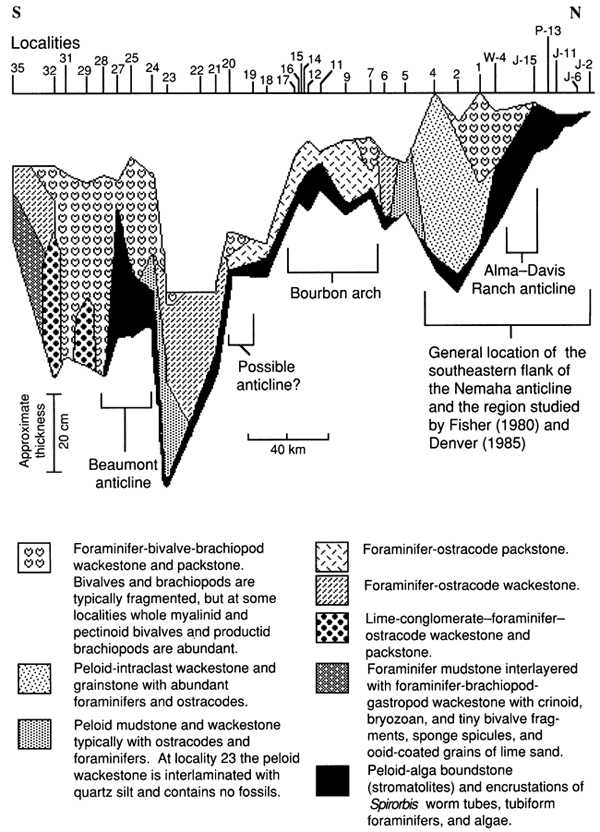
Figure 23--Schematic cross section of the lower limestone bed of the Americus Limestone Member showing distribution of lithofacies resulting from deposition under relatively high- or low-energy conditions and locations of structural features. Datum is the base of the limestone above the lowermost limestone of the Americus. [See Jewett and Merriam (1959) and Fisher (1980).]
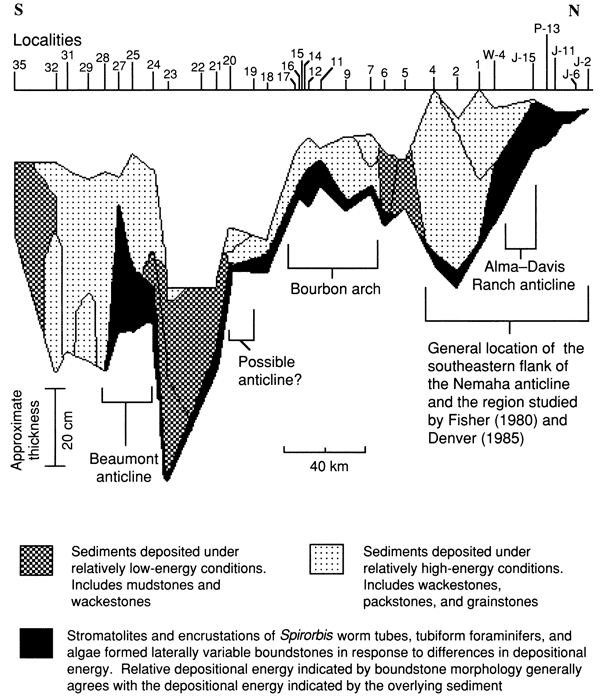
Figure 24--Photomicrograph of conglomeratic foraminifer-ostracode wackestone to packstone from locality 29 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,010.
Morphology of boundstone as an indicator of paleoenvironment
Recognition of paleoenvironmentally controlled features in the peloid-alga boundstones (stromatolites) and associated Spirorbis-foraminifer-alga boundstones provides clues to the conditions that prevailed at the time of deposition. Although the biology of algae and other microorganisms is an important control of morphology of stromatolites (Raaben, 1969; Golubic, 1976; Monty, 1977), such environmental conditions as wave energy, desiccation, and substrate stability have also been shown to affect morphology (Logan, 1961; Gebelein, 1969; Logan et al., 1974; Hardie and Ginsburg, 1977; Hoffman, 1976).
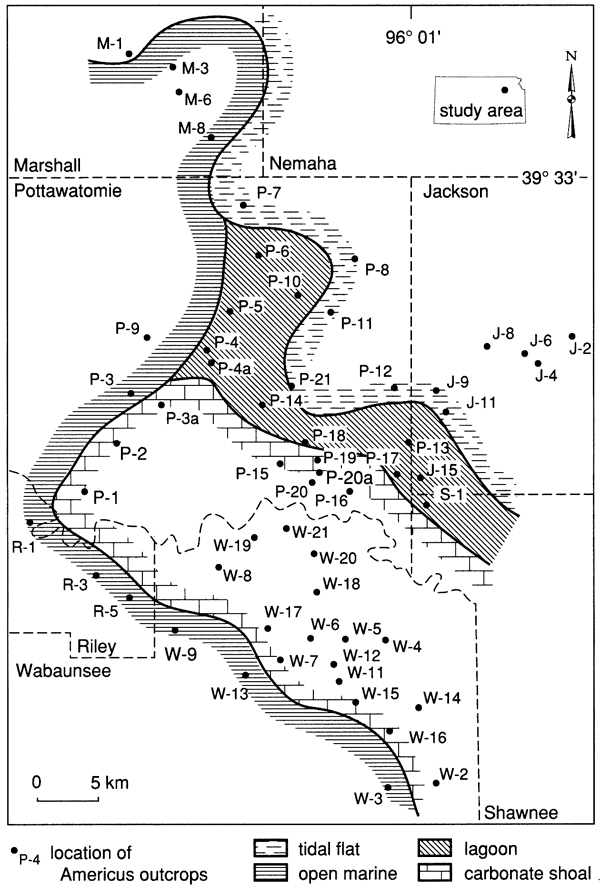
Boundstones in the Americus Limestone Member occur as far north as locality P-7 (Fisher, 1980) in central Pottawatomie County and, as rare clasts, as far south as locality 31 in northern Elk County, Kansas. Lack of boundstones to the north is evidently the result of subaerial conditions (Fisher, 1980), which were unfavorable to the growth of algae (Hoffman, 1976), encrusting tubiform foraminifers, and Spirorbis worms. The lack of intact peloid-alga boundstones south of locality 28 may be the result of more nearly normal marine conditions to the south, indicated by the greater abundance of marine fossils and the occurrence of such typically marine features as ooids and grapestone. This southern environment may have favored metazoans, which prevented the development of stromatolites by grazing and by disrupting the substrate (Garrett, 1970; Awramik, 1971; Monty, 1973; Hoffman, 1976; Gebelein, 1976).
Boundstones in the Americus Limestone Member occur in six paleoenvironmentally significant morphologic types (forms): (1) discrete plates of Spirorbis-foraminifer-alga boundstone; (2) laterally discontinuous alga boundstone with extensive fenestrae; (3) rip-up clasts of boundstone; (4) thick layers of peloid-alga boundstone with extensive encrustations; (5) high-domed peloid-alga boundstone; and (6) thin, flat-layered to low-domed peloid-alga boundstone. The distribution of the types of boundstone morphology of the lower bed of the Americus is provided in fig. 25.
Figure 25--Distribution of types of boundstone morphology of the lower limestone bed of the Americus Limestone Member.
Type 1 morphology
Type 1 morphology consists of discrete plates of Spirorbis-foraminifer-alga boundstone (fig. 26), which Fisher (1980) described as pustular crusts of Ottonosia. These plate-shaped masses, typically 3-20 cm (1.2-7.9 in.) across and 5 cm (2 in.) thick, contain abundant Spirorbis worm tubes, tubiform foraminifers, algae, and less abundant fragments of brachiopods. This morphology differs from encrustations of Spirorbis-foraminifer-alga boundstone found elsewhere in the Americus because it occurs as discrete plates independent of peloid-alga boundstone, is vertically cracked in its top surface, and contains pervasive pore spaces lined with multiple generations of pendant PF2 cement and filled with terrigenous mudstone (figs. 27 and 28). Type 1 boundstone is characteristic of the lower limestone of the Americus at its northern limits.
Figure 26--Top surface of hand sample of type 1 boundstone (discrete plates of Spirorbis-foraminifer-alga boundstone) from locality P-11. Slab (right) shows typical thickness, and arrow indicates the up direction. Scale bar is 2 cm. KUMIP 227,011.

Figure 27--Photomicrograph of pendant PF2 cement (arrow A) within type 1 boundstone from locality P-11 (plane-polarized light). Arrow B points to a Spirorbis worm tube. Scale bar is 1 mm. KUMIP 227,012.
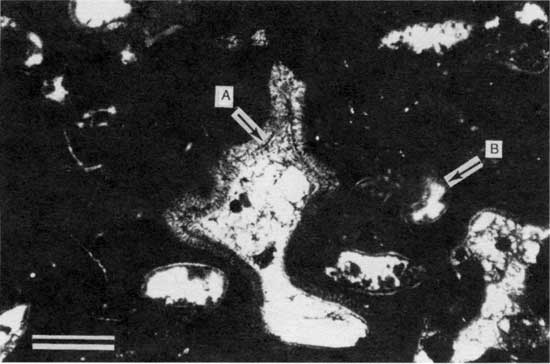
Figure 28--Photomicrograph showing close-up of pendant cement shown in fig. 27 (plane-polarized light). Scale bar is 1 mm.
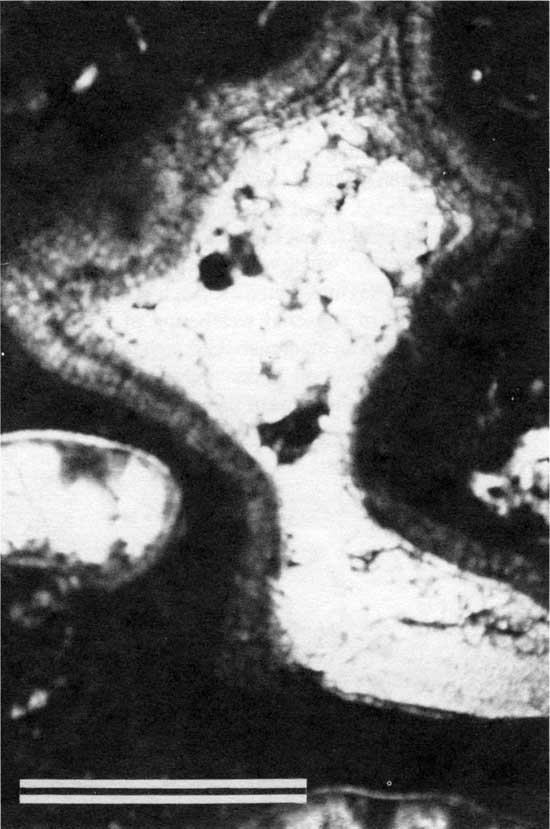
Abundant Spirorbis worm tubes indicate shallow-water restricted marine conditions (Abe, 1943). The vertical cracks in the tops of the plates suggest desiccation resulting from subaerial exposure. The pendant habit and fibrous crystals of cement suggest precipitation in a marine phreatic environment (Longman, 1980). These features indicate that deposition occurred on a paleotopographically high seafloor that was occasionally subaerially exposed. Fisher (1980) interpreted this type as having formed in a tidal flat (fig. 29). Brachiopod fragments also indicate proximity to normal marine paleoenvironments. The mud that covered and infiltrated this boundstone probably was deposited at a later stage, when somewhat deeper marine water covered the paleotopographic high.
Figure 29--Fisher's (1980) paleoenvironmental facies of the lower Americus limestone bed in north-central Kansas. [From Denver (1985), after Fisher (1980).]
Type 2 morphology
Type 2 morphology consists of laterally discontinuous alga boundstone with extensive fenestrae (fig. 30). Denver (1985, p. 24) described such boundstones as "laminoid, domal, ... largely of limonitic mud, silt, and scattered peloids ... [and] characterized by horizontal and vertical fenestrae up to 2 cm long that are lined with clear, equant spar but otherwise open." Type 2 boundstones occur in laterally discontinuous patches, 15 cm (5.9 in.) wide and 2-3 cm (0.8-1.2 in.) high. Associated fossils, including Spirorbis tubes, are rare. Denver (1985) noted that this morphology is rare and that it is generally surrounded by shale. Type 2 boundstones occur in the same general geographic area as type 1 boundstones, that is, at the northern limits of the lower limestone bed.
Figure 30--Polished slab of type 2 boundstone (isolated boundstone with extensive fenestral porosity). Scale bar is 2 cm. KUMIP 214,825.
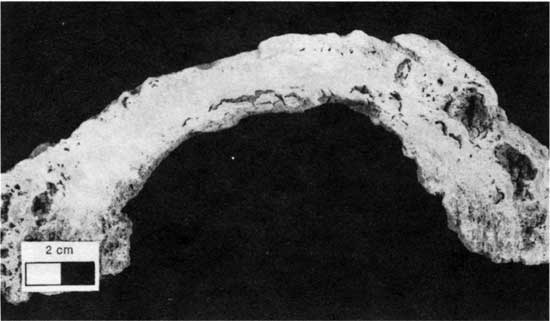
Denver (1985) found that the type 2 morphology occurs only in the area that Fisher (1980) identified as a tidal flat facies and concluded, based on pervasive fenestral porosity and desiccation cracks, that the rocks formed in an upper intertidal environment which was frequently subaerially exposed. Denver (1985) interpreted the patchy lateral distribution as the result of subaerial weathering of a polygonal pattern developed in an algal mat. Although domal features can result from deposition under high-energy conditions (Hoffman, 1976), the fine size of the incorporated sediment suggests that this morphology did not develop in a high-energy environment. In this case the domal features may have formed from arching caused by gas trapped beneath the algal mat.
Type 3 morphology
Type 3 morphology is characterized by rip-up clasts of boundstone. These rocks consist of small intraclasts of both peloid-alga boundstone and Spirorbis-foraminifer-alga boundstone and large clasts of peloid-alga boundstone. The smaller clasts are generally no larger than 7 mm (0.3 in.) across and are irregular (fig. 31). Larger clasts are 1-2 cm (0.4-0.8 in.) thick and up to 20 cm (8 in.) wide (fig. 32). The clasts are generally in a matrix of foraminifer-ostracode packstone or wackestone and are typically associated with thick domal boundstone (fig. 33). At locality 25 the clasts seem to be concentrated in a 0.4-m-deep (1.3-ft-deep) trough developed between deposits of boundstone. The clasts generally are not coated, but some larger ones have been encrusted with both Spirorbis-foraminifer-alga boundstone and peloid-alga boundstone (fig. 34). Fisher (1980) noted extremely thick encrustations of Spirorbis-foraminifer-alga boundstone around clasts of peloid-alga boundstone, which he referred to as reef rock. These reef rocks, which are actually encrustations on large stabilized oncolites, are typically 20 cm (8 in.) thick but may be "40 cm high and nearly a meter across" (Fisher, 1980, p. 44). Clasts of boundstone are numerous at localities 1, 2, 4, 9, 24, and 25, but they also occur at localities 7, 11, 22, and 31.
Figure 31--Photomicrograph of type 3 boundstone [rip-up clasts (arrows) of peloid-alga boundstone] from locality 2 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,013.
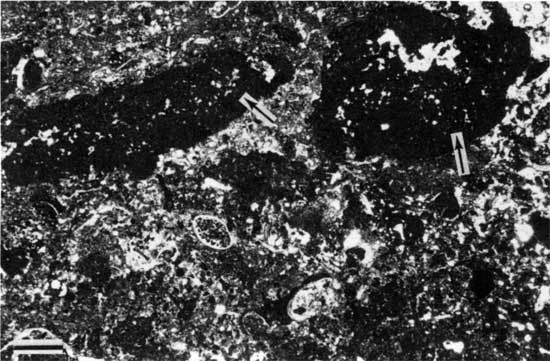
Figure 32--Polished slab showing type 3 boundstone (rip-up clasts of peloid-alga boundstone) from locality 9. Scale bar is 2 cm. KUMIP 227,014.

Figure 33--polished slab showing rip-up clasts of type 3 boundstone (arrows) and proximity to domal (type 5) and thick (type 4) boundstone from locality 24. Scale bar is 2 cm. KUMIP 227,015.
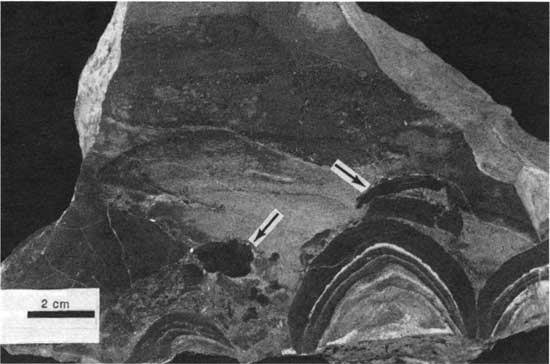
Figure 34--Photomicrograph of rip-up clasts (type 3 boundstone) with organic coatings from locality 2 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,016.
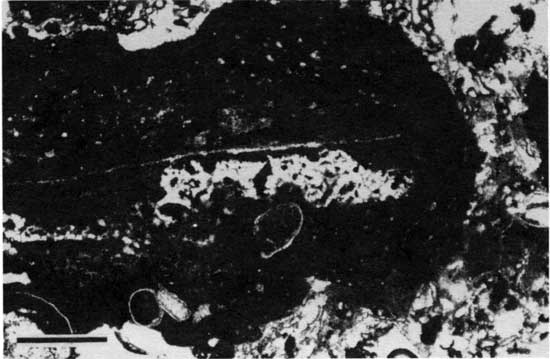
In the north rip-up clasts of boundstone are limited to the vicinity of Fisher's (1980) carbonate-shoal and lagoonal facies. Relatively thin overlying shale at locations 9, 24, and 25 (see fig. 22) suggests deposition in relatively shallow water. The clasts are interpreted as the result of storm erosion in areas of shallow water adjacent to areas of relatively deep water. Rolling during subsequent storms allowed some clasts to acquire oncolitic coats of boundstone. The abundance of oncolites in Fisher's (1980) lagoonal facies suggests that part of the environment was a swash zone rather than a protected lagoonal environment. Boundstones that were not ripped up, however, must have been protected from high-energy waves, developing behind a protective barrier, on a substrate below the wave base, or in wide, shallow areas that damped waves.
Type 4 morphology
Type 4 morphology is characterized by relatively thick layers of peloid-alga boundstone and extensive encrustation with Spirorbis-foraminifer-alga boundstone. This morphology includes Denver's (1985) smooth and laminoid forms. Type 4 boundstones are generally 5-20 cm (2-8 in.) thick and contain coalescing columnal, hemispherical, and horizontal laminations of peloid-alga boundstone. Stromatolites with this morphology typically display extensive ragged pores filled with foraminifers and ostracodes from overlying sediment and are lined with encrustations of Spirorbis-foraminifer-alga boundstone (fig. 35). Laminations in the lower 5 cm (2 in.) are typically disrupted and brecciated and commonly have alveolar structures (figs. 12 and 13).
Figure 35--Polished slab showing type 4 boundstone (thick, porous, encrusted boundstone with disrupted laminations) from locality 25. Scale bar is 2 cm. KUMIP 227,017.
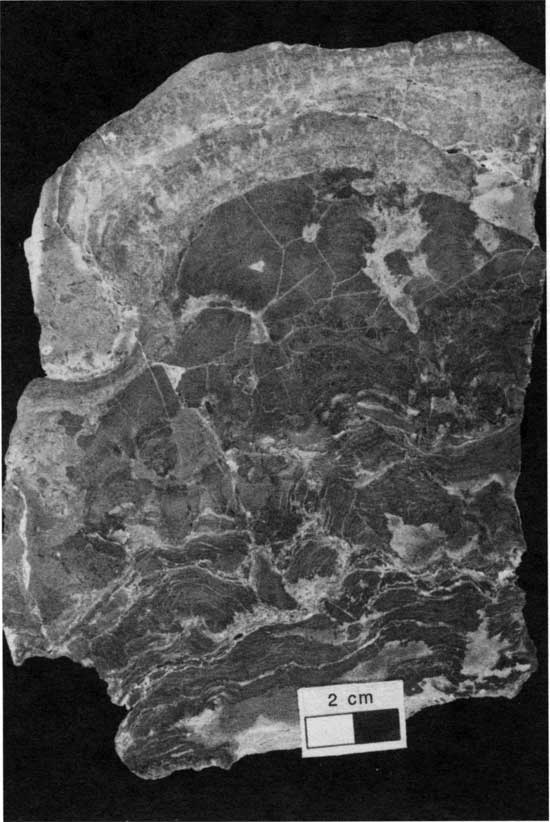
The upper surfaces of the peloid-alga boundstone commonly display two generations of encrustations. The first generation is a Spirorbis-foraminifer-alga boundstone, up to 11 cm (4.3 in.) thick, containing common tiny bivalves (figs. 15 and 16) and having a truncated upper surface. The second generation of encrustation is a coalescing digitate foraminifer-alga boundstone with bryozoans incorporated in the upper portions (fig. 20). This second generation is typically bored (fig. 15), but its upper surface is not truncated.
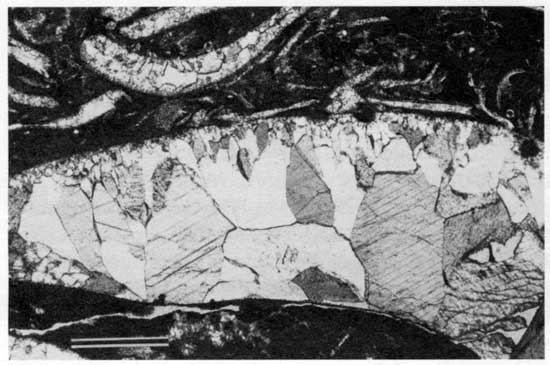
Type 4 morphology occurs at several locations in Fisher's (1980) carbonate-shoal facies and elsewhere at localities 11, 24, 25, and 27. At all locations type 4 boundstones are associated with a dramatic thinning of the overlying shale, suggesting deposition on a substrate of relatively high paleotopography. The coalescing columnar and hemispheroidal laminations and the relatively high relief of the boundstone suggest deposition in a relatively high-energy environment (Logan, 1961; Gebelein, 1969; Hoffman, 1976; Playford and Cockbain, 1976). Alveolar structures and associated disrupted layers indicate a period of subaerial exposure that enabled vegetation to develop. These features can be explained by a lowering of sea level that exposed isolated areas of high paleotopography. The upper layers of peloid-alga boundstone contain no alveolar features, indicating that subaerial exposure was temporary. Under subsequent deeper-water conditions the paleotopographically high seafloors were evidently ideal habitats for encrusting foraminifers and Spirorbis worms.
Type 5 morphology
Type 5 morphology, a variant of Denver's (1985) smooth and laminoid morphologies, is characterized by layers of peloid-alga boundstone arched into relatively high domes (fig. 36). The height of the domes typically is 4-7 cm (1.63 in.), and the width ranges from 5 cm to 15 cm (2-6 in.). The domes typically have an inner core of orange shale or terrigenous mudstone. Encrustations of Spirorbis-foraminifer-alga boundstone are generally less than 1 cm (0.4 in.) thick. At location 21 postdepositional compaction has partially crushed the domes. Type 5 morphology occurs at localities 12, 21, 24, and 25.
Figure 36--Large domes of peloid-alga boundstone typical of type 5 boundstone from locality 12. Scale bar is 2 cm. KUMIP 227,018.
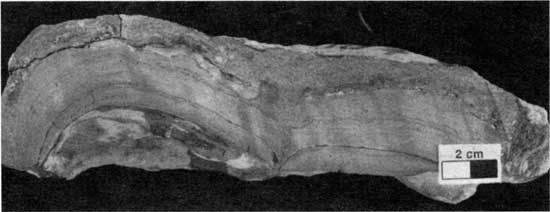
As with morphology types 3 and 4, the high-relief domal features of type 5 morphology probably indicate deposition in moderately high-energy environments (Logan, 1961; Gebelein, 1969; Hoffman, 1976). Figure 21 suggests that localities 12, 24, and 25 were probably the sites of relatively high-energy deposition on a seafloor with relatively high paleotopography.
Type 6 morphology
Type 6 morphology is characterized by low-domed to thin, flat layers of peloid-alga boundstone. These boundstones would have been classified by Denver (1985) as either smooth or laminoid. The layers generally range in thickness from 0.6 cm to 3.5 cm (0.2-1.4 in.) and commonly occur as laterally discontinuous plates, 12-30 cm (4.7-12 in.) wide, with the edges curled under. Low domal features, if present, generally have a height of 1.5 cm (0.6 in.) and are typically oblong in plan view with a maximum length of 12 cm (4.7 in.). Encrustations of Spirorbis-foraminifer-alga boundstone on the upper surface of the peloid-alga boundstone are patchy and 0.4-1 cm (0.2-0.4 in.) thick. Type 6 morphology occurs at localities 2, 15, 16, 17, 18, 19, 20, 22, 23, and 28.
At localities 18, 19, 22, and 23 the boundstone is characterized by relatively flat layers with microcolumns and tiny troughs (fig. 37). These flat layers are 2 cm (0.8 in.) thick and are dissected by tiny troughs, 4 mm (0.2 in.) wide and 1.5 cm (0.6 in.) deep, that form microcolumns 2 cm (0.8 in.) wide. In plan view the trough pattern is roughly polygonal. The troughs are generally filled with foraminifer-ostracode packstone to wackestone and often contain tiny gastropods. Laminae of peloid-alga boundstone are generally turned down at the edges of the microcolumns. The trough and microcolumnar structures are believed to have resulted from grazing by tiny metazoans along the edges of desiccation cracks developed in a thin, flat to undulated mat of algally bound peloids. Tiny cracks would have provided refuge for such small metazoans as ostracodes and gastropods, which may have widened the cracks by browsing. Subsequent algal activity formed additional boundstone layers that either turned down at the edge of the trough or, more rarely, bridged the trough to connect adjacent microcolumns. The laterally discontinuous plates with curled-under edges probably resulted from subaerial modifications of larger desiccation polygons early in the inundation.
Figure 37--Microcolumns and troughs developed in the low-relief layers of type 6 boundstone from locality 22. Scale bar is 2 cm. KUMIP 227,019.
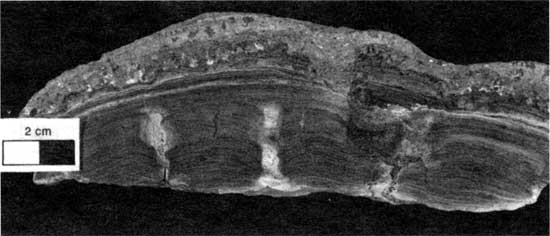
The low relief of this morphology suggests deposition in a low-energy environment (Logan, 1961; Gebelein; 1969; Hoffman, 1976; Playford and Cockbain, 1976). Figure 21 shows that localities with type 6 morphology have relatively thick intervals of shale and thus probably had deeper-water low-energy environments at maximum transgression. This morphology evidently developed in the wide low-lying areas that were the initial sites of inundation. With continued transgression these initial deposits were soon below the wave base and thus did not develop features indicative of a high-energy environment. Shale probably first accumulated in these low-lying areas and evidently prevented extensive encrustations of Spirorbis, foraminifers, and algae.
Distribution of fossils and other constituents as indicators of paleoenvironment
Lateral distribution and relative abundance of fossils and other constituents provide additional information on the paleoenvironment of the lowermost bed of the Americus Limestone Member. Figure 38 shows that such normal marine fossils as calcareous sponge spicules, echinoid spines, bryozoan fragments, crinoid fragments, and, to some extent, gastropods are more abundant to the south. Fragments of brachiopods occur at many locations, but unfragmented Linoproductus, Derbyia bennetti, and dictyoclostid brachiopods are abundant only at locality 28 on the flank of the southernmost paleotopographically high seafloor. Ooids, ooid-coated grains, and grapestones (fig. 39)--features typical of agitated seafloors under normal marine conditions (Bathurst, 1975)--occur at only a few southern outcrops. This evidence suggests that normal marine influences were greater in the south than in the north. Subaerial exposure at paleotopographically high locations is indicated by pendant PB5 cement (fig. 40) within molds of bivalves from locality 25 and alveolar structures at localities 24 and 25.
Figure 38--Distribution and relative abundance of fossils and other rock constituents at each locality.
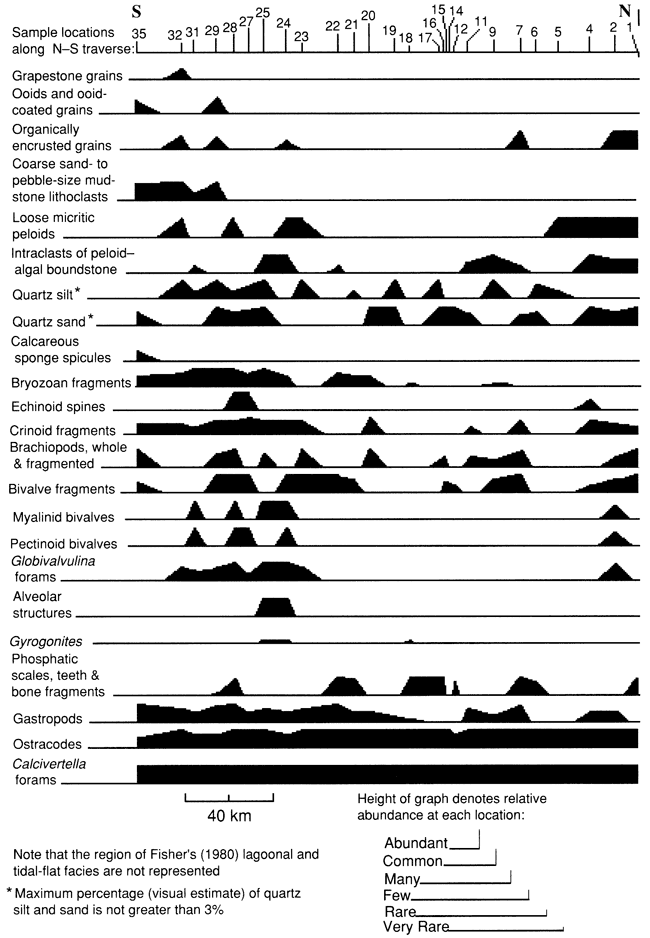
Figure 39--Photomicrograph of ooid-coated foraminifer (A) and ostracode (B) from locality 29 (plane-polarized light). Scale bar is 100 µm. KUMIP 227,020.
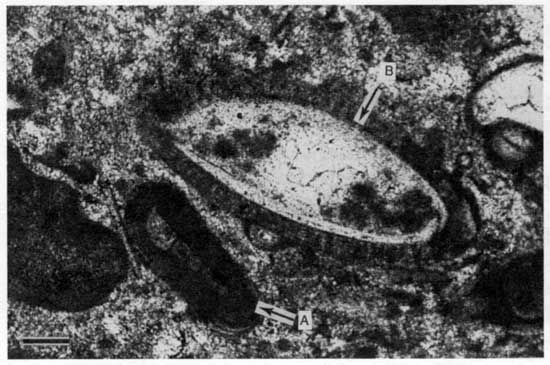
Figure 40--Photomicrograph of bivalve molds from locality 25 displaying pendant PB5 cement (cross-polarized light). Scale bar is 1 mm. KUMIP 227,021.
The paleoenvironmental significance of the distributions of other constituents shown in fig. 38 is more problematic. Gyrogonites is present at localities 18, 24, and 25 but is quite rare. It may have been derived from the underlying freshwater lime sand of the Hamlin Shale Member. Indeed, the lower limestone bed at localities 24 and 25 displays a basal surface with irregularities that suggest erosion, and at these localities Gyrogonites occurs with ripped up fragments of boundstone. If not allogenic, Gyrogonites may have originated from charophytes living in freshwater during periods of subaerial exposure on paleotopographically high areas.
The environmental significance of the distribution of silt- and sand-size quartz grains and tiny fragments of bone, teeth, and scales typically found immediately above the stromatolite layer is not known. Some of the scales are from acanthodian fish, which are believed to have lived in both marine and freshwater environments (Schultze, 1985). Calcivertella foraminifers, ostracodes, and tiny high-spired gastropods are present at nearly all locations and occur in greater abundance than any other fossil in the lowermost bed of the Americus. This abundant low-diversity fauna suggests deposition in an overall restricted environment. Kaesler et al. (1990) examined the distribution patterns of ostracode species to determine their paleoenvironmental significance.
The distribution of Globivalvulina foraminifers and pectinoid and myalinid bivalves provides insight into their environmental preferences. Globivalvulina foraminifers (fig. 41) are more abundant toward the south, suggesting that these foraminifers preferred more nearly normal marine conditions. Furthermore, Globivalvulina and pectinoid and myalinid bivalves (fig. 42) are bimodally distributed, with clusters located between localities 1-4 and localities 23-32. These were locations of relatively high depositional energies on paleotopographically high seafloors, suggesting that these organisms preferred such habitats. The distribution may be similar to that of the modern Pinna-Pinctada community in Shark Bay, Australia, which is characteristic of sublittoral sills covered by no more than 2 m (7 ft) of water and periodically exposed during the summer (Read, 1974, p. 15).
Figure 41--Photomicrograph of the foraminifer Globivalvulina (arrow) from locality 29 (plane-polarized light). Scale bar is 100 µm. KUMIP 227,020.
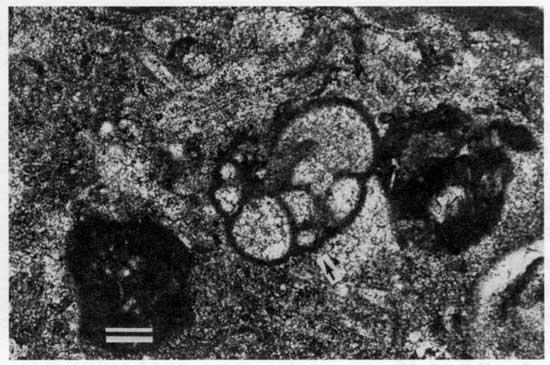
Figure 42--Myalinid (A) and pectinoid (B) bivalves on the weathered surface of the lowermost limestone bed of the Americus Limestone Member from locality 31. Scale bar is 2 cm. KUMIP 227,022.

Prev Page--Paleoenvironments || Next Page--Structural Features, Conclusions
Kansas Geological Survey, Geology
ISBN: 1-58806-107-8
Placed on web Sept. 8, 2011; originally published 1992.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/Sub13/04_dist.html