![]()
Prev Page--Introduction || Next Page--Distribution
Vertical succession of paleoenvironments
The Hamlin Shale and Americus Limestone members contain a vertical succession of facies indicative of deposition during eight stages of inundation by an epeiric sea. The following discussion refers to fig. 3, which shows a generalized stratigraphic section of the upper Hamlin and the Americus, the corresponding stages of inundation, and the associated paleoenvironments. (See appendix F for a description of the lithologic units of the Hamlin shale and the Americus limestone in Kansas.)
Figure 3--Generalized vertical section of the upper part of the Hamlin Shale and the Americus Limestone members showing typical rock types and fossils, weathering profile, and paleoenvironments and correlative stages of inundation by epeiric sea.
Stage 1: Paleoenvironments of the Hamlin shale before inundation
The Hamlin Shale Member contains features that indicate deposition under hypersaline, brackish, freshwater, and terrestrial conditions that preceded inundation by the sea responsible for deposition of the Americus Limestone Member. Laterally discontinuous stromatolites with interlaminated gypsum suggest deposition under hypersaline restricted conditions (Kendall and Skipwith, 1969). The low faunal diversity in laterally discontinuous, ostracode-containing lime mudstones and wackestones further indicates deposition in a restricted environment, possibly a result of fluctuating salinity or temperature. The light color of the typically orange, white, tan, light-gray, and brown shales and terrigenous mudstones may indicate oxidizing terrestrial conditions. Boxwork structures (Read, 1974), fenestral porosity (Shinn, 1968; Grover and Read, 1978), pervasive skew and horizontal joint planes (Freytet and Plaziat, 1982; Goldstein, 1986), and polygonal mudcracks are interpreted as the effects of desiccation in supratidal to subaerial environments.
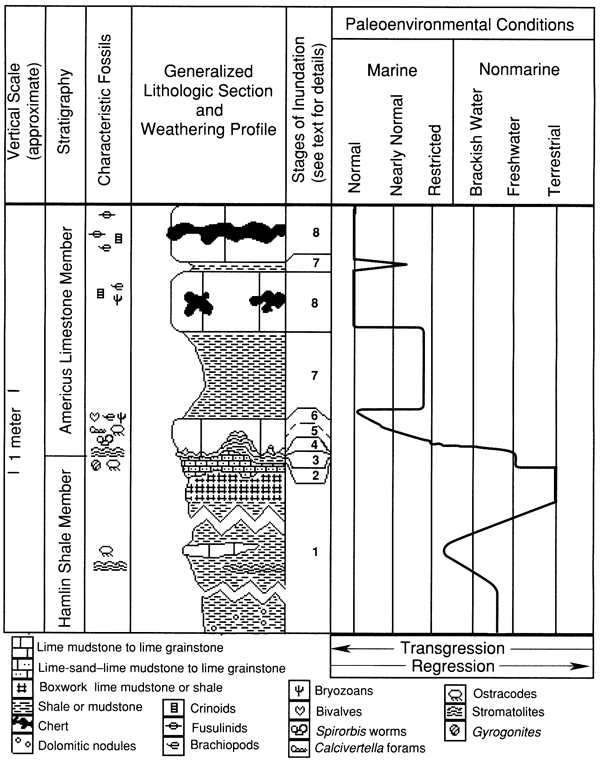
A distinctive orange lime-sand rock unit at the top of the Hamlin Shale Member is evidently the result of deposition by freshwater flowing over an erosional surface. This lithology differs between localities from an orange shale or terrigenous mudstone with a few sand-size lime grains to a conglomeratic lime-sand grainstone with medium sand sized to pebble-size lithoclasts; however, the rock is typically a lime-sand packstone or wackestone (fig. 4). Its thickness varies from 0 to 0.3 m (0-1 ft). Ostracodes and small, well-rounded bivalve fragments are typical; crossbedding is generally observed where the unit is thick. This rock contains no fossils or cements with definite marine affinities. Up to 9 cm (4 in.) of relief on the basal surface of the lime-sand bed suggests erosion, possibly by streams flowing in small channels. The relatively large size of the lime grains indicates sediment transport by moderately strong currents. The lime grains are allochthonous lithoclasts of lime mudstone and are well rounded, suggesting that the sediment was transported across an appreciable distance. Gyrogonites, the fruiting bodies of charophyte algae, are typically present and suggest deposition associated with a freshwater environment based on analogy with modern charophytes (Wray, 1977, pp. 110-111). Coal stringers, plant fragments, and petrified logs have been noted at some localities (Bernasek, 1967; Fisher, 1980), providing additional evidence of deposition in or near freshwater. These features suggest that the Hamlin was being eroded by streams just before inundation by an epeiric sea.
Figure 4--Photomicrograph of lime-sand packstone to grainstone of the upper Hamlin Shale Member from locality 6 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,000.
Stage 2: Initial inundation by the epeiric sea
At many localities an orange to gray shale or terrigenous mudstone, generally less than 1 cm (0.4 in.) thick, was deposited directly over the orange lime-sand packstone or wackestone of stage 1. This shale or terrigenous mudstone resulted from deposition of suspended fine sediments from relatively quiet water, indicating a significant decrease from depositional energies of the underlying lime-sand rocks. The paleoenvironmental significance of this shale or terrigenous mudstone is problematic. It may have resulted from flooding by freshwater streams at the end of stage 1, or it may have developed during initial inundation by seawater that terminated the stream activity of stage 1. The inundating waters were probably saline, but the orange color of the shale or terrigenous mudstone suggests affinity with the freshwater paleoenvironment of the underlying orange lime-sand rocks. It is therefore reasonable to assume that the water at this stage was brackish or fluctuated between marine water and freshwater. Fluctuations between saline and freshwater conditions would have produced an environment unfavorable to most marine and freshwater metazoans and thus would have favored growth of stromatolites (stage 3).
Stage 3: Transition from brackish to restricted marine conditions
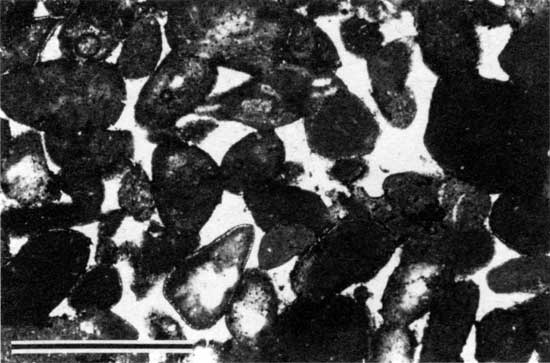
The base of the Americus Limestone Member is characterized by a stromatolite layer that probably formed during the transition from brackish-water to restricted marine conditions. Stromatolites are the result of sediment binding (Black, 1933; Logan, 1961; Logan et al., 1964; Gebelein, 1969) and possibly precipitation of calcium carbonate (Dalrymple, 1966; Monty, 1967) by microbial communities dominated by blue-green algae (Golubic, 1976). The stromatolites of the Americus generally occur as laminations of micritic peloids and are classified as peloid-alga boundstones (Dunham, 1962; see appendix A). The peloids are irregularly shaped to oval and are generally coarse silt to fine sand sized (fig. 5). In thin section these peloids appear to float in PE32 calcite cement (Folk, 1965; see appendix B). Internal laminations are generally 1-6 mm (0.04-0.2 in.) thick. Cement-filled horizontal fenestrae are common (fig. 6). Typically, no grains other than peloids are incorporated in the boundstone layers. Exceptions are rare mollusk fragments, lime-sand grains, and palynomorphs (fig. 7).
Figure 5--Photomicrograph of peloid-alga boundstone from locality 7 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,001.
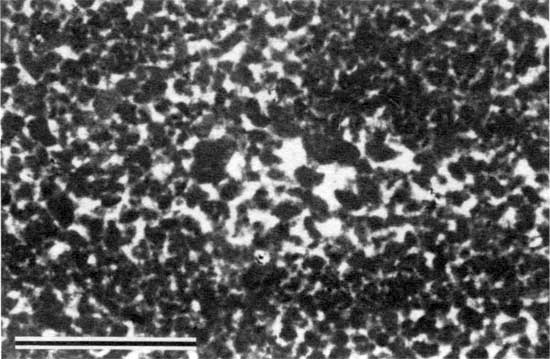
Figure 6--Photomicrograph of fenestral porosity (arrow) in peloid-alga boundstone from locality 17 (plane-polarized light). Scale bar is 100 µm. KUMIP 227,002.
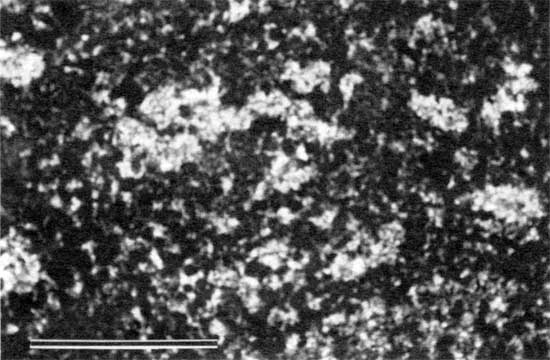
Figure 7--Photomicrograph of palynomorph (arrow) in peloid-alga boundstone from locality 12 (plane-polarized light). Scale bar is 100 µm. KUMIP 227,003.
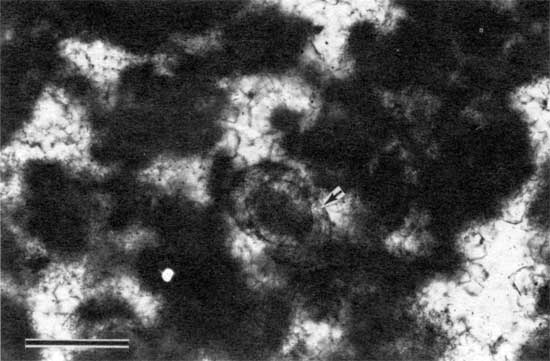
Traces of algal filaments were noted in many samples (figs. 8 and 9). These traces occur in dense micritic laminations and appear in thin section as subtle horizontal threadlike features. The filaments, never longer than 0.5 mm (0.02 in.), have bright micritic hollow centers, about 0.02 mm (0.0008 in.) thick, and dark micritic walls, about 0.01 mm (0.0004 in.) thick. These dimensions compare favorably with those of Scytonema, a modern stromatolite-producing alga (Bathurst, 1975; Golubic, 1976), but are slightly larger than those of the modern Oscillatoriales, including Schizothrix (Golubic, 1976). The dimensions of the filaments are also about the same as those of some species of Girvanella (Riding, 1975; Wray, 1977); however, the irregular and indistinct outline of the filament traces in the stromatolites of the Americus are unlike those generally observed in Girvanella.
Figure 8--Photomicrograph of algal filaments (arrow) in peloid-alga boundstone from locality 7 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,001.
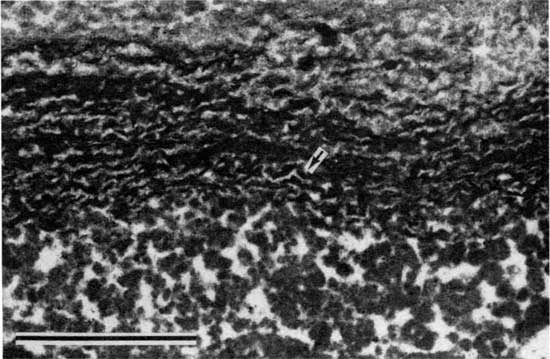
Figure 9--Photomicrograph showing in greater detail the same algal filaments as those in fig. 8 (plane-polarized light). Scale bar is 100 µm.
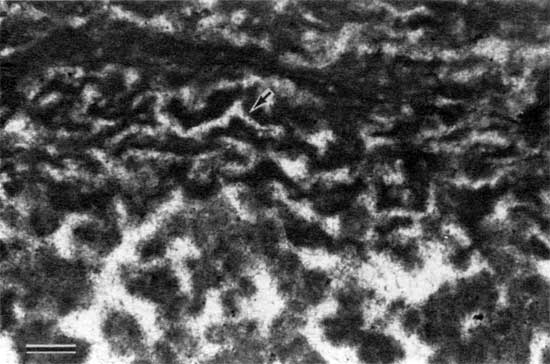
Rare pseudomorphs of evaporites occur within the algal laminates (figs. 10 and 11). At some localities the lower laminations of the stromatolite are disrupted and contain alveolar structures (figs. 12 and 13).
Figure 10--Photomicrograph of pseudomorphs of evaporites (arrow) in peloid-alga boundstone from locality 18 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,004.
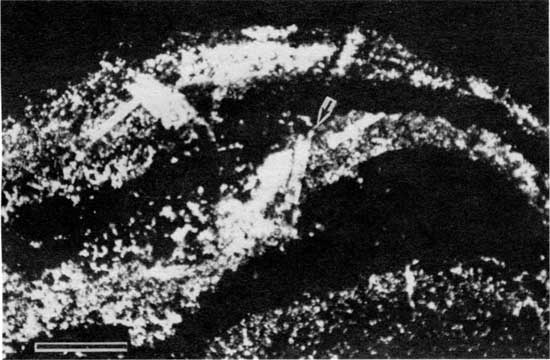
Figure 11--Photomicrograph at higher magnification of the same pseudomorphs of evaporites shown in fig. 10. Scale bar is 1 mm.
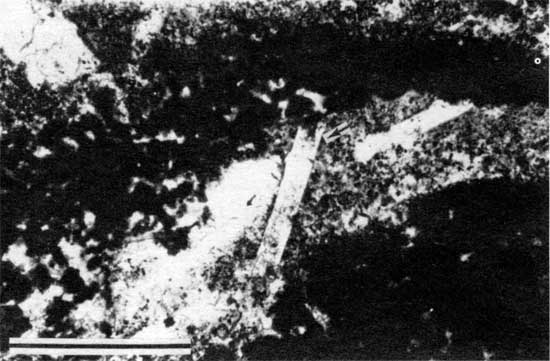
Figure 12--Photomicrograph of disrupted laminations and alveolar structures (arrows) in peloid-alga boundstone from locality 25 (plane-polarized light). Scale bar is 1 mm. KUMIP 227,005.
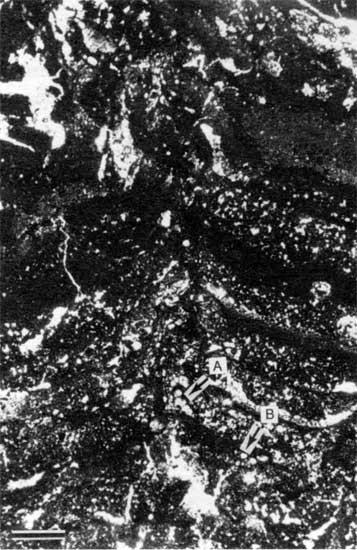
Figure 13--Photomicrograph of alveolar structure (A) from fig. 12 (plane-polarized light). Scale bar is 100 µm.
The rarity of incorporated fossil fragments indicates that the depositional conditions were too harsh for the existence of most metazoans. Such harsh conditions may have been the result of fluctuating salinity or temperature, desiccation, or anoxic or otherwise poisonous conditions caused by the stromatolites themselves (Monty, 1973). Conditions of fluctuating salinity are suggested by the stratigraphic position of the stromatolites between nonmarine and marine deposits and the presence of rare pseudomorphs of evaporites. The fenestral porosity suggests early diagenesis in shallow-water to subaerial environments (Shinn, 1968; Grover and Read, 1978) and may have resulted from the oxidation of algal matter (Monty and Hardie, 1976). Alveolar structures, a type of root trace (Esteban and Klappa, 1983), suggest that the stromatolitic layer was affected by a brief period of subaerial exposure, probably the result of a minor fluctuation in sea level. The thickness and morphology of the stromatolite layer varies laterally from locality to locality, a result of laterally different paleoenvironmental conditions. This lateral variation is described in detail in later sections.
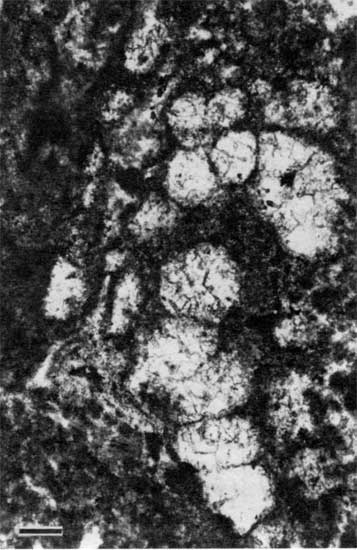
Stage 4: Restricted marine conditionsWhen formation of peloid-alga boundstone ceased, the exposed surface became encrusted with Spirorbis, foraminifers, and algae under shallow-water restricted marine conditions. These encrustations formed a boundstone ranging in thickness from <1 cm (<0.4 in.) to 19 cm (7.5 in.). In thin section these boundstone encrustations appear as ragged, porous, dense, micritic crusts containing abundant to common Spirorbis worm tubes and tangled tubiform features (fig. 14). Tiny bivalves (figs. 15 and 16) are enmeshed within the thick encrustations at localities 25 and 27. The taxonomic affinity of the tubiform features is problematic because of poor preservation. The micritic tube walls are only slightly darker and the tube centers only slightly brighter than the surrounding micrite. The tubes are nonbranching and have an outside diameter of 0.03-0.05 mm (0.001-0.002 in.). The diameter and morphology of the tubes conform to those of Girvanella (Riding, 1975; Wray, 1977) and tubiform foraminifers and may be the remains of either or of both (Henbest, 1963; Toomey et al., 1977).
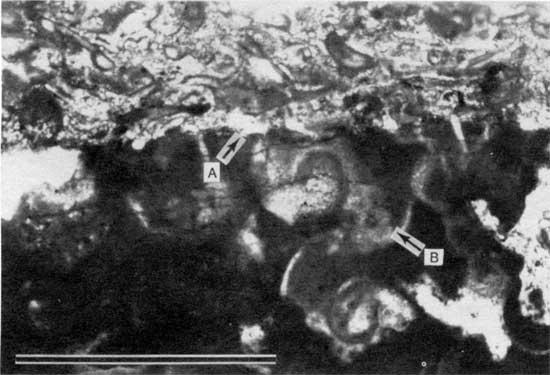
Figure 14--Photomicrograph of encrustation of Spirorbis-foraminifer-alga boundstone on peloid-alga boundstone from locality 11 (plane-polarized light). Arrow A points to Spirorbis worm tube; arrow B points to a tubiform encruster, probably a foraminifer or an alga. Scale bar is 1 mm. KUMIP 227,006.
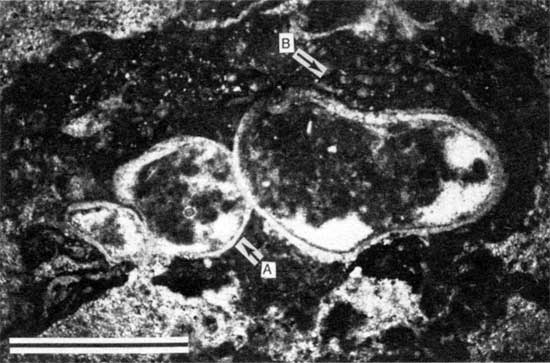
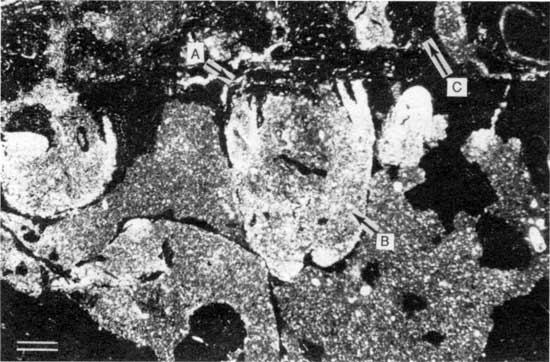
Figure 15--Polished slab from locality 27 showing thick encrustation of Spirorbis-foraminifer-alga boundstone (bracket) on peloid-alga boundstone. Arrow A points to bores in foraminifer-alga boundstone; arrow B points to bivalve within Spirorbis-foraminifer-alga boundstone; arrow C points to disrupted peloid-alga boundstone. Scale bar is 2 cm. KUMIP 227,007.

Figure 16--Photomicrograph of truncated bivalves within Spirorbis-foraminifer-alga boundstone from locality 25 (plane-polarized light). Arrow A points to truncated surface; arrow B points to truncated bivalve; arrow C points to digitate foraminifer-alga growths on truncated surface. Scale bar is 1 mm. KUMIP 227,008.
The abundance of Spirorbis worm tubes suggests deposition in shallow restricted marine water. Modern Spirorbis worms typically inhabit the littoral zone and thrive in restricted tidal pools. They can tolerate brief periods of subaerial exposure and are unaffected by temperature fluctuations from extremely warm to subfreezing (Abe, 1943). Toomey and Cys (1977) described the occurrence of Spirorbis in stromatolites from what they interpreted as restricted marine to brackish water deposits, and Toomey and Cys (1979) regarded similar foraminifer-alga boundstones as a pioneer-stage community that formed in shallow marine water. Water depth may have been the factor responsible for restricted conditions. A covering of relatively shallow marine water probably would have lacked strong circulatory currents, and solar heating would have elevated the temperature and salinity. Salinity of such a shallow body of water may also have fluctuated as a result of dilution by precipitation. The truncated upper surface of the Spirorbis-foraminifer-alga encrustations (figs. 16 and 17) may have resulted from high-energy conditions during storms or from bioerosion during a later, more nearly normal marine stage.
Figure 17--Photomicrograph of Spirorbis-foraminifer-alga boundstone showing typically truncated upper surface from locality 18 (plane-polarized light). Boundstone is overlain by a foraminifer-ostracode wackestone to packstone. Arrow A points to truncated surface; arrow B points to a Spirorbis worm tube. Scale bar is 1 mm. KUMIP 227,004.
Stage 5: Transition from restricted marine to nearly normal marine conditions
Stage 5 is characterized by foraminifer- and ostracode-rich lime wackestones and packstones and by peloid-intraclast wackestones to grainstones. These deposits formed in a restricted marine environment that became nearly normal marine with time. The fauna was dominated by Calcivertella foraminifers, ostracodes, and tiny high-spired gastropods (figs. 18 and 19). Calcivertella are normally encrusters, but in the Americus Limestone Member they are generally unattached. Their typically curved to circular base of attachment indicates encrustation on perishable objects, probably aquatic vegetation. The great abundance of free Calcivertella suggests that this vegetation was abundant. Spirorbis is generally absent in this stage; however, a foraminifer-alga boundstone formed at some locations on the truncated surface of the underlying Spirorbis-foraminifer-alga boundstone. This subsequent encrustation was digitate, lacked worms, and incorporated bryozoans toward the top (fig. 20). Features indicative of relatively high depositional energies developed at localities interpreted as paleotopographic highs. These high-energy features include rip-up clasts of peloid-alga boundstone, algae-coated grains (oncolites), and packstone or grainstone depositional textures.
Figure 18--Photomicrograph of ostracodes and Calcivertella from locality 18 (plane-polarized light). Arrow points to a Calcivertella within an ostracode. Scale bar is 1 mm. KUMIP 227,004.
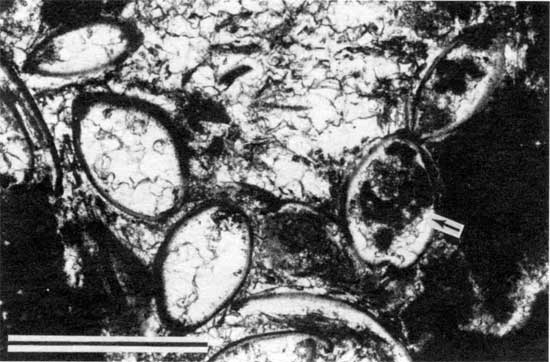
Figure 19--Photomicrograph of an ostracode and Calcivertella from locality 11 (plane-polarized light). Note curved base of attachment (arrow) on the foraminifer. Scale bar is 100 µm. KUMIP 227,006.
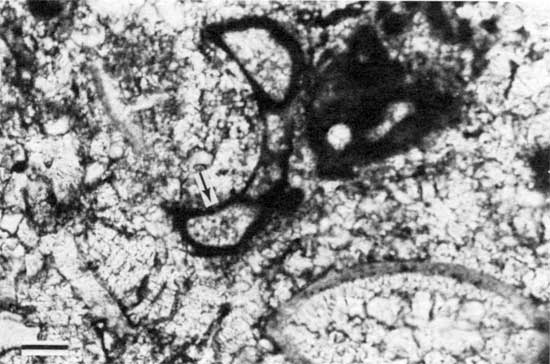
Figure 20--Polished slab showing coalescing digitate foraminifer-alga boundstone (bracketed) containing bryozoans, developed on truncated surface (arrow) cutting Spirorbis-foraminifer-alga boundstone and peloid-alga-boundstone from locality 27. The light-gray zone directly over the foraminifer-alga boundstone is a diagenetic effect and does not contain boundstone. Scale bar is 2 cm. KUMIP 227,009.

The low diversity but great abundance of fossils in this stage suggests deposition in a restricted marine environment. High-energy features, presumably the result of waves and storms in areas of shallow water, indicate that, adjacent to shallow areas, the water had become deep enough to allow the generation of waves that dissipated their energy on shoals. The occurrence of such normal marine fossils as brachiopods, bryozoans, and crinoid fragments, although rare, suggests that the environment was becoming more nearly normal marine.
Stage 6: Nearly normal to normal marine conditions
Fossils and features indicative of marine conditions are prevalent near the top of the lower limestone of the Americus Limestone Member. These include echinoid spines, crinoids, bryozoans, brachiopods, sponge spicules, pectinoid and myalinid bivalves, grapestone, and ooids. The foraminifers Globivalvulina, Tetrataxis, and Syzaria occur at this stage. Borings with a diameter of 3 mm (0.1 in.) in the foraminifer-alga boundstone of stage 5 probably occurred during this stage.
The high-diversity fauna with marine affinities indicates deposition under normal marine conditions. Many of these organisms relied on currents for delivery of food and dispersal of larvae, indicating that the water was deep enough to allow circulatory currents to develop. The continued occurrence of Calcivertella, however, suggests partial persistence of the conditions of stage 5.
Stage 7: Turbid marine conditions
Deposition in turbid marine waters resulted in deposits up to 2.5 m (8.2 ft) thick of black, brown, and gray shales. These shales consist of a lower shale situated between the lower and upper limestone beds and a thinner upper shale within the upper limestone bed of the Americus Limestone Member. At some localities thin beds of micaceous quartz sand are interbedded with the lower shale. The fine grain size indicates that the terrigenous material was probably derived from a distant source and accumulated in the deeper, quieter portions of the inundated shelf. Sparse fragments of crinoids, brachiopods, and bivalves reported by Fisher (1980) in the lower shale suggest deposition under restricted marine conditions. Perhaps extensive colonization by normal marine organisms was restricted by the turbid water and slightly lower salinity that may have accompanied the influx of terrigenous sediment. More nearly normal marine conditions evidently were associated with the development of the upper shale, as suggested by the prevalence of fusulinids at some localities.
Stage 8: Normal marine conditions
The upper limestones of the Americus Limestone Member contain a diverse fauna that indicates normal marine conditions. This fauna includes abundant fusulinids, crinoids, brachiopods, bryozoans, and bellerophontid gastropods. [See Harbaugh and Demirmen (1964) and Fisher (1980) for discussions on paleoenvironmental trends within the upper limestone beds of the Americus. Also see appendix G, outcrop 32.]
Prev Page--Introduction || Next Page--Distribution
Kansas Geological Survey, Geology
ISBN: 1-58806-107-8
Placed on web Sept. 8, 2011; originally published 1992.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/Sub13/03_paleo.html