![]()
Prev Page--Geology || Next Page--Paleoecology and Paleoclimatology
Biostratigraphy
Vertebrate Faunas in Ellis County
Vertebrate fossils were discovered in the Tertiary deposits of Ellis County as early as 1875 (Mudge, 1876). Mudge reported the remains of a three-toed horse and indicated that it might be assigned to the Miocene genus Anchiterium. No further significant work on vertebrates of the Tertiary deposits in Ellis County was conducted, however, until Zehr (1974), at the encouragement of R. J. Zakrzewski, studied the fossil vertebrates from deposits on the western edge of Ellis County. His study resulted in the establishment of the Hamburg local fauna that be considered to be an early Clarendonian vertebrate fauna. He assigned an early Pliocene age (now considered late Miocene) to this fauna. Twenty-one taxa of mammals and a number of lower vertebrates were recovered, and he concluded that they indicated "a humid, subtropical climate, abundant deciduous vegetation along the river, grass-covered slopes on the flood plain, with the existence of a nearby savanna."
Several new vertebrate localities were discovered during the course of my fieldwork. Many of the vertebrate remains have been identified both by me and vertebrate paleontologists. These vertebrates provide important evidence as regards the paleoecology and paleoclimatology of the geologic periods during which the fossil plant-bearing strata were deposited.
During late July 1974, several vertebrate fossils (Bemis local fauna) were discovered a few feet above the contact of beds 1 and 2 at site 29 (Fig. 4D). Subsequently, I opened a small excavation and recovered a number of other vertebrates, and later, with the aid of Mr. Harold D. Gross, Hays, Kansas and Dr. R. J. Zakrzewski, expanded the dig. From that time until June 1975, a large number of vertebrate remains and fragments were taken. The Bemis local fauna was subsequently studied by M. E. Tallan (1978) of the Sternberg Memorial Museum. Her identifications of the material are presented in Table 3.
Vertebrate material at site 39 was initially discovered in early July 1975. The fossils (Flinn local fauna) were recovered from the upper 150 cm of bed 2, along with an intimately associated and beautifully preserved flora of fossil plants. Fossil plants recovered at this site are shown on Plate I, graphic section 39. The snake and turtle remains were identified by J. Alan Holman, Museum, Michigan State University, and the other vertebrates were identified by R. J. Zakrzewski, Sternberg Memorial Museum, Fort Hays Kansas State College (Table 4). My experiments revealed that the usual methods of vertebrate recovery, such as screening or washing, recovered the vertebrates but destroyed the anthoecia, nutlets, and endocarps. It is essential that no collection method except handpicking be practiced at this site. Fortunately, the sediments at this site are readily disaggregated by degaging under a binocular microscope.
A few vertebrate fragments have been found near the north end of site 7 in the lowermost exposure of bed 1. Zehr ( 1974) reported, and I have seen, a very large land tortoise (Geochelone) exposed at the contacts of bed 1 and 2 in the north half of the same exposure. Remains from this site were identified by J. Alan Holman (Table 5).
Table 5--Identified fossil vertebrates recovered at site 7.
| JRT Specimen No. |
Specimen | Remarks |
|---|---|---|
| 7-2, 7-3, 7-4 | Large tortoise (Geochelone orthopygia) dermal ossicles | All tortoise fragments collected from same individual |
| 7-5 | Same as above but xiphiplastron | |
| 7-no numbers | Assorted large plastral and carapacial fragments |
At site 6, vertebrates have been discovered at a number of localities (Table 6). Fossils were generally found in densely cemented "mortar beds" and were recovered only with great difficulty. Remains of peccaries were identified with the assistance of R. G. Corner, Museum, University of Nebraska.
Table 6--Identified fossil vertebrates recovered at site 6.
| JRT Specimen No. |
Specimen | Locality | Remarks |
|---|---|---|---|
| 6-1, 6-2 | Large tortoise (? Geochelone sp.) Carapace and plastron | 1. (bed 11 of 12) | Fragments only recovered; main fossil left in place |
| 6-13 | Peccary (Prostennhops graffhammi); inferior P4 that is part of 6-14 | Same as 6-1, 6-2 | Probably all peccary material is from a single individual; all peccary material has been closely compared with the type or referred material of Prostennhops graffhammi at UNSM. (UNSM 76052, 76054, 76071). 6-1, 6-2, 6-13, 6-14, 6-15, 6-17 collected within 18 in. of each other |
| 6-14 | Same as 6-13 but rear two-thirds of right lower jaw including M3 and M2 | Same as 6-1, 6-2 | |
| 6-15 | Same as 6-13 but a median phalanx | Same as 6-1, 6-2 | |
| 6-16 | Same as 6-13 but a right tibia | Same as 6-1, 6-2 | |
| 6-17 | Same as 6-13 but a left tibia | Same as 6-1, 6-2 | |
| 6-no | Large tortoise numbers (? Geochelone) carapacial fragments | 3, upper bed 8 |
Vertebrate fossils at site 41 were discovered during a brief visit in early June 1975. Later visits revealed this site to be of special significance since only one type of vertebrate, large thin-shelled (8-25 mm) land tortoises, was discovered. They were, however, very abundant and on my second visit I discovered five separate individuals exposed in bed 2. In one of these was found the remains of a pelvic girdle (Fig. 5D). If fully explored, this site should prove to be an important new locality for fossil tortoises.
The jaw of a small rodent, which appears to be that of a ground squirrel, was found in association with a cache of seeds in the upper part of bed 1, of site 33. A small unidentified limb bone was found in the lower, exposed 60 cm of bed 1.
Floral Zones in High Plains Tertiary Deposits
An examination of the history of floral zones in the High Plains Tertiary deposits is necessary to a better understanding of the important changes in use of floral zones proposed in this paper. Because previous workers have used anthoecia, nutlets, and endocarps for zonation in a slightly different way, it is appropriate to investigate these differences and to comment on those aspects of this history which have contributed to recent questions concerning floral zones by this author and others (Webb, 1969; Zehr, 1974; Galusha, 1975; Thomasson, 1976b, 1977).
Elias ( 1931) was the first investigator to use fossil anthoecia, nutlets, and endocarps in a biostratigraphic sense in the High Plains Tertiary deposits. He collected a number of fossil angiosperm anthoecia (Berriochloa glabra), nutlets (Biorbia fossilia), and endocarps (Celtis) and described a "zone of fossil herbs" in the middle portion of the Ogallala Formation. Although Elias described the "zone of fossil herbs" as occurring in beds of a particular lithology, clearly his zone was biologic in nature. The following year, in a systematic report on grasses and other plants from the Tertiary rocks of Kansas and Colorado, Elias (1932) described several species of grasses and borages, mentioning again the "fossil herbs zone." A considerable elaboration and refinement of floral zones was published by him in 1933. Later Elias (Hibbard, 1934) illustrated a zonation specifically related to western Kansas sediments. Two zones were illustrated, a lower zone of Krynitzkia coroniformis followed by an upper zone of Biorbia fossilia. The floral zone of Krynitzkia coroniformis had its upper limit near the top of the Woodhouse clays. In 1935 Elias elaborated on his earlier zonations, concluding that many of the fossil "seeds" could be used as index fossils to determine the age of rocks from which they came. This represents the first time the angiosperm remains from the Ogallala Formation were used to determine ages of rocks enclosing them.
Three years later Chaney and Elias (1938) developed a zonation of all the Tertiary strata above the Arikaree Formation in the High Plains areas of Nebraska and Colorado. Five (possibly six) zones were recognized. Celtis, which had originally been considered an index species of its own floral zone (Elias, 1934, 1935), was no longer considered so, and a zone of Echinochloa sp. nov. was in approximately the same position. Of the zones proposed, four were based on undescribed taxa.
Lugn (1938, 1939) presented charts in which "seed" zones in the High Plains Tertiary deposits were illustrated. Both charts were essentially the result of Elias's previous 10 years research and both tie biostratigraphic floral zones firmly to rock-stratigraphic units. Lugn concluded that, "It seems probable that the use of fossil seed zones may well mark a new epoch in Tertiary stratigraphic investigation for nearly all late Miocene and Pliocene sediments in the Great Plains Region."
The early era of studies culminated in Elias's "Tertiary Prairie Grasses and Other Herbs From the High Plains" published in 1942. This work summarized the conclusions of Elias with regard to the zonation of the Ogallala on the basis of the known "seeds." The zonation presented by Elias was little questioned in the literature until quite recently (Galusha, 1975; Thomasson, 1976b, 1977).
Except for an occasional reference to fossil anthoecia, nutlets, or endocarps recovered incidentally to other geologic studies, no further investigations appeared until Frye, Leonard, and Swineford (1956) studied the Ogallala Formation in northern Kansas. They examined a large number of stratigraphic exposures and presented a detailed summary of the occurrence of fossilized "seeds." They concluded that:
In summary, it must be emphasized that for the field stratigrapher interested in local and regional correlations, fossil seeds are the most useful stratigraphically of all the kinds of organic remains found in the Ogallala sediments.
Frye and Leonard (1957, 1959) expanded significantly the known areal and temporal distribution of "seeds" in the Ogallala Formation. They established fossil "floral" zones and used them to correlate the Ogallala Formation in Texas with the type of localities in Nebraska. They recognized three floral zones which they considered stratigraphically equivalent with the Valentine, Ash Hollow, and Kimball Members of the Ogallala Formation of Kansas classification and with the Valentine, Ash Hollow, and Sidney-Kimball Formations of Nebraska. This floral zonation was later maintained, without modifications, by Frye and Leonard (1964) and Frye (1971). Schultz, Schultz, and Martin (1970) indicated that these floral zones agreed in general with faunal evidence and that, "The Valentine floral zone would be of Valentinian provincial age, the Ash Hollow floral zone would be of Clarendonian and Hemphillian ages, and the Kimball floral zone would be of Kimballian age."
Floral Zones in Ellis and Rooks Counties, Kansas
I am convinced, as others before me, that fossil anthoecia, nutlets, endocarps, and achenes from the Tertiary sediments of the High Plains are one of the most important biostratigraphic tools available. Preservation is generally detailed, and many fossils are readily identifiable with a common binocular micro- scope. Their widespread temporal and areal distribution in these deposits is unequaled by any other group of fossils. Even the smallest outcrops frequently yield large numbers of "seeds" in a wide variety of taxa. The frequent association of a number of taxa makes possible more accurate correlations of strata. The sediments in Ellis County contain numerous fossil anthoecia, nutlets, endocarps, and achenes assignable to many taxa. From studies of fossils collected with strict stratigraphic control it is now possible to comment on their biostratigraphic significance. A new biostratigraphic zone is proposed and a stratotype for this zone is designated. Nomenclature for this new biostratigraphic zone follows American Association of Petroleum Geologists (1961) and Hedberg (1972a, b).
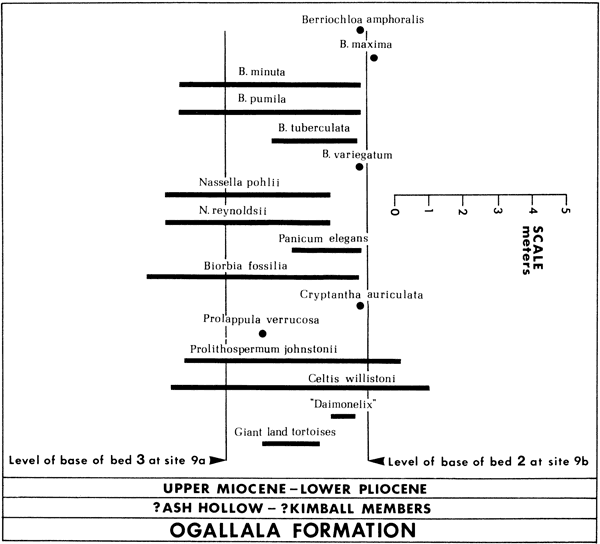
The name of the new unit is the Prolithospermum johnstonii-Nassella pohlii Assemblage Zone. The diagnostic fossils of this zone are Prolithospermum johnstonii Elias and Nassella pohlii sp. nov. Other fossils present in this zone are N. reynoldsii sp. nov., Panicum elegans Elias, Berriochloa minuta Elias, B. tuberculata Elias, B. maxima Elias, B. amphoralis Elias, B. variegatum (Elias) Thomasson, comb. nov., B. pumila sp. nov., Prolappula verrucosa, gen. nov., sp. nov., Cryptantha auriculata (Elias) Segal, Biorbia fossilia (Berry) Cockerell, and Celtis willistonii (Cockerell) Berry. Included in this zone are the occurrences of "Daemonelix" (see discussion of "Daemonelix" in the systematic portion of this study).
The stratotype, or specific sequence of rock strata on which the zone is established, is located at WL SW NW NW Sec. 3, T. 12 S., R. 20 W. Ellis Co., Kansas and includes those strata between 1.8 meters (6 ft.) below the contact of bed 3 at site 9a and 1.0 meters (3 ft.) above the base of bed 2 (= bed 5 at site 9a) at site 9b (Pl. I, graphic sections 9a and 9b; Fig. 6A,B; Fig. 7). Reference sections are located at sites 8 and 39. The new zone is shown pictorially in Figure 6, and the sequence of fossils is shown diagrammatically in Figure 7. At least at the stratotype locality and others nearby, the P. johnstonii-N. pohlii Zone appears to be associated with a distinct suite of lithologic units. At the base is a coarse sandstone (locally gravel); the medial portions are predominantly reddish silts to medium sands (frequently extremely well sorted); and the upper parts are characterized by coarse conglomerates. At least two diatomaceous freshwater limestone deposits (marls) and a volcanic ash bed are included within this zone (Pl. I, graphic sections 9a, 9b; Figs. 6A,B). Fossil charophytes have been described recently from the marl layers (Daily and Thomasson, 1978). A lithologic description of site 9a is given in the Appendix.
Figure 6--Stratotype section of Prolithospermum johnstonii-Nassella pohlii Assemblage Zone. A. View looking east at site 9a. a-a: vertical extent of strata in new zone. B. View looking northeast at site 9a. Site 9b is in background.

Figure 7--Chart showing the stratigraphic position of fossil angiosperm anthoecia, nutlets, and endocarps at the stratotype section of the Prolithospermum johnstonii-Nassella pohlii Assemblage Zone. Level of base of bed 2 at site 9b is identical with the level of the base of bed 5 at site 9a. P. johnstonii-N. pohlii zone includes sediments 1.8 m below the base of bed 3 at site 9a through 1 m above the base of bed 2 (= bed 5 at site 9a) at site 9b.
The history of biostratigraphy in the Ogallala has been characterized by attempts to associate the floral zones with established lithostratigraphic units, when these rock units themselves were the subject of serious debates. The floral zones were frequently established on the basis of widely separated, often fragmentary collections of "seeds," and then correlations across great distances were made between rock-stratigraphic units on the basis of these fossils. Eventually, the floral zones were equated with the rock-stratigraphic units.
Because of questions concerning the various interpretations of "seed" occurrences, I have reexamined the collections of Elias and Leonard. Additionally I have made extensive new collections to "test" the established floral zones. I found no evidence to confirm the established zones and, as will be shown, fossils from Ellis County deposits occur in such a manner as to contradict significant parts of the established zones.
A comparison of the floral zones of the Ogallala of Elias (1942), Frye and Leonard (1959), and the newly established P. johnstonii-N. pohlii Zone illustrates that past interpretations of the "seeds" in Ash Hollow or Kimball floral zones are no longer tenable. In Ellis County deposits, Prolithospermum johnstonii, considered to be the index fossil of the Kimball floral zone (Elias, 1942; Frye and Leonard, 1959), significantly overlaps in range a number of fossils diagnostic of the Ash Hollow floral zones. Frye and Leonard (1959) do not illustrate Cryptantha (Krynitzkia) auriculata, but indicate (p. 25) that this taxon is "restricted to the lower third of the Ash Hollow floral zone." Elias also considered this taxon as diagnostic of lower Ash Hollow deposits and in fact established a Cryptantha (Krynitzkia) auriculata subzone. Clearly, the position of Cryptantha auriculata with respect to Prolithospermum johnstonii in Ellis County deposits (Fig. 7) contradicts these previous interpretations.
Likewise, the ranges of Panicum elegans and Berriochloa minuta in the Ellis County section do not agree with the conclusions of previous workers. Thus, the diagnostic fossils and their ranges in each zone are no longer representative of the known facts. Therefore the Ash Hollow and Kimball floral zones as previously described must be abandoned. The present studies confirm that the Kimball and Ash Hollow floral zones do not exist, and the "seeds" that have been found appear to combine the "seeds" of both zones with several significant range extensions. It might be argued that the designation Ash Hollow or Kimball should be retained for any resulting single zone, but there is no compelling reason to do so. The retention of either designation would obscure the essential biostratigraphic nature of the zone being established. In addition, attempts to bring nomenclatorial stability to geology would suffer.
While the present studies may contradict the findings of previous authors, we may not undervalue the painstaking and laborious efforts of those who have helped to lay the foundations of our present knowledge of the High Plains Tertiary deposits. Widespread reconnaissance studies such as those conducted by Frye and Leonard have been and will continue to be as essential as the present detailed study if we are to eventually learn the intricacies of the Tertiary deposits. Their studies for example have dramatically extended the known geographic range of "seed" floras, including fossil anthoecia and endocarps recently reported for the first time from deposits in New Mexico (A. B. Leonard, University of Kansas, personal communication, July, 1976). One of the most important sites in my study area, site 9, was initially visited as a result of consulting a study by Frye, Leonard, and Swineford (1956).
For all these reasons, I have found it necessary to establish the new zone. If fossil anthoecia, nutlets, endocarps, and achenes are to have maximum significance in the interpretation of late Tertiary deposits, they must be collected not only in widespread reconnaissance studies but in large numbers and with strict stratigraphic control in restricted areas. Hopefully, this study will be a stimulus to such future studies.
Galusha (1975) has called to question the credibility of some of the lower zones of Elias (1942). Since the sediments involved are much older than those in Ellis County, I cannot comment with any expertise on the findings of Galusha. A restudy of the fossils and their ranges in the involved deposits is needed.
Prev Page--Geology || Next Page--Paleoecology and Paleoclimatology
Kansas Geological Survey, Geology
Placed on web May 1, 2009; originally published September 1979.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/218/04_bios.html