![]()
Prev Page--Paleoecology and Paleoclimatology || Next Page--References
Systematic Paleontology
Scanning Electron Microscopy
Specific details of the micromorphology obtained by SEM for individual fossil taxa will be discussed in the systematic section of this paper. In the following section the results obtained will be summarized for each family.
Gramineae
Distinctive epidermal patterns were observed on the lemmas and paleas of all genera of fossil grasses examined. However, the detail observed on individual specimens varied from extremely poor to excellent.
Within the genus Berriochloa, long cells, prominent hooks, and hairs have been observed. The pattern is predominantly one of long cells. These walls are always sinuous ("rippled") with well-developed lobes. These lobes vary between taxa and, within limits, on individual specimens. The lobes may be predominantly simple, as in B. amphoralis (Pl. 8A), B. conica (Pl. 8B, C), B. minuta (Pl. 9A), and B. primaeva (Pl. 9B); or compound as in B. maxima (Pl. 8E, F) and B. inflata (Pl. 8D). Within the simple types, the tips of the lobes may be expanded and broader than the base, with a constricted neck (Pl. 8B, C). The tips may also be only slightly or not at all expanded and terminate in an obtuse (Pl. 9C) or acute apex (Pl. 9B). If the lobes are compound, the lobules may dichotomize regularly (Pl. 8D) or they may arise irregularly (Pl. 8E, F). The lobes, whether simple or compound, may be longer than the width of the central area of the long cell (Pl. 9E), shorter (Pl. 8E, F), or of about the same width (Pl. 8B, C; 9A).
Hooks (see Metcalfe, 1960) are frequently seen on species of Berriochloa. The hooks are broad at the base and terminate in a curved point (Pl. 2E). Where the hooks occur, the long cells develop massive lobes that buttress the hooks (Pl. 9E; 10A). As many as six to as few as three of these "buttress" lobes have been seen to surround a single book. Hooks frequently grade into longer structures towards the summit of the lemma which might be termed large prickles (Metcalfe, 1960, pl. VI, Fig. 2).
Hairs observed on the lemma and callus of some specimens of Berriochloa (Pl. 3A) are long and terminate in a sharp point. Apparently these hairs were stiff in the living plants since, as preserved on the fossils, they are always perfectly straight.
Features of the palea in Berriochloa are similar to those of the lemma. Long cells, hooks, and hairs were variously observed. Short cells were not observed on any of the species of Berriochloa.
Anthoecia of genus Nassella have a pattern of epidermal cells that differs radically from that of Berriochloa. In Nassella, the basic long cells have been strikingly reduced in length, although they still retain the ripple walls common to many long cells (Pl. 10E). Numerous cells that apparently represent silica and suberin cells (silica-suberin couples) are also present. The silica cells are indistinguishable from the suberin cells except by shape. In fossil Nassella reynoldsii the difference is well marked, the silica cells being nearly ovate, while the suberin cells are crescent shaped (see Kaufman et al., 1970, Fig. 14, for an excellent illustration of a silica-suberin couple in an extant grass). The distinction between the members of the pair also seems fairly obvious in N. fryeleonardii (Pl. 10D), although the silica cell tends to be more hemispherical than ovate and frequently is elongated. When this occurs it may not be possible to distinguish the silica cells from the suberin cells.
Hooks are present on the lemmas of N. amphora, N. fryeleonardii and N. reynoldsii (Pl. 3H; 4A, B, D, E). These are very similar to those on species of Berriochloa and offer no distinction between the two genera. Hairs were not observed on any of the fossil species of Nassella.
The only other grass genus found preserved as fossils was Panicum. The microfeatures were well preserved on both the lemma and palea, and consisted of long cells and papillae. The long cells found preserved on both the lemma and palea were very similar. They are basic long cells with sinuous "ripple" walls and well-developed lobes (Pl. 11A). This pattern is similar to the pattern of long cells in Berriochloa. No short cells were observed, but papillae were present on the surface of the palea (Pl. 4H) and on the lower edges of the lemma near the germination lid (Pl. 4I). The papillae are simple rounded structures and are almost identical with those illustrated by Clark and Gould (1975) in some modern Panicum species. The papillae at the base of the lemma were irregularly arranged but those of the palea were in distinct rows. I have discussed elsewhere the significance of the microfeatures in fossil and living Paniceae (Thomasson, 1978a).
Cyperaceae
Only a single fossil genus with a single species has been examined by scanning electron microscopy during this study. Excellent patterns of epidermal cells were revealed.
The pattern observed on Eleofimbris gen. nov. achenes consists entirely of approximately rectangular cells whose long axis is oriented vertically. The walls of the cells are weakly undulate and the central area of the cells is concave (Pl. 11C). Nodules or buttresses, as described for some Cyperaceae by Schuyler (1971), were not observed on the fossils.
Boraginaceae
Representatives of the five fossil borage genera Biorbia, Cryptantha, Eliasiana, Prolappula, and Prolithospermum were examined.
Nutlets of the genus Biorbia exhibit a pattern of irregularly arranged epidermal cells. These frequently, but not always, have a long axis. The cell walls are sinuous with well-developed lobes which are of a simple type (Pl. 11F). Ridges, when present, may be well developed (Pl. 7D) or obscure (Pl. 7C).
The genus Cryptantha has nutlets which are characterized by a pattern of irregularly lobed epidermal cells. The lobes are well developed and may be simple or compound (Pl. 11E). In addition, the surface of the nutlets may be characterized by protuberances which are rounded at the apex (Pl. 5F). These have been previously referred to in the literature as tubercles (nodules). No pattern was observed in the arrangement of these tubercles on the surface. Frequently these tubercles are absent. Very strongly developed protuberances, present on the margin of the nutlets, are termed wings or auricles (Pl. 5G, H).
No epidermal pattern was observed in the newly described genus Eliasiana. However, the surface of the nutlets was characterized by an abundance of short prickles (aculeae) (Pl. 6F).
Within the genus Prolappula, no epidermal patterns were observed. However tubercles with secondary protuberances, barbs, and broad-based, elongated prickles with secondary pustules were noted (Pl. 7F, G, H). Tubercles were randomly dispersed on the surface of the nutlets. These were always more or less bulbous and had secondary protuberances that were cylindrical in shape. Scattered among the tubercles were broad-based, erect, elongated prickles. These prickles were themselves secondarily beset with protuberances which appear as pustules. At the margin of the nutlets were found barbed hairs (Pl. 7F, G). Small protuberances were observed on the stalk of the barbs.
The final genus studied in the Boraginaceae was Prolithospermum. A well-developed epidermal pattern of randomly arranged epidermal cells was observed. The cells possessed simple lobes which were often abruptly angled (Pl. 11D). Complex pseudoepidermal patterns which I interpret as the result of corrosion (? solution) were also observed on some specimens. Ridges similar to those in Biorbia were sometimes present (Pl. 7B). No tubercles or other protuberances were seen.
Significance of Scanning Electron and Light Microscopy Studies to the Phylogeny of Fossil and Living Gramineae
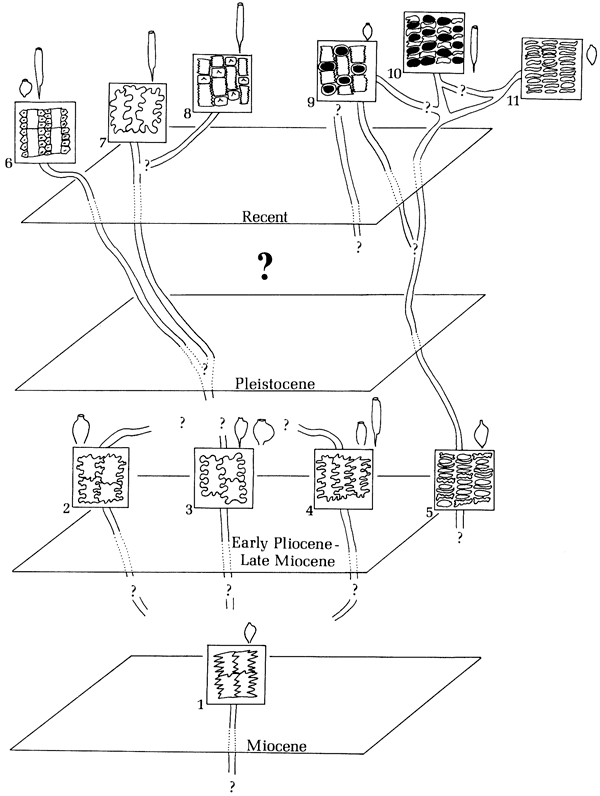
Clarification of the phylogenetic relationships within living groups related to the fossils was also essential if the fossils were to be related to the living taxa. Unfortunately, there was a lack of overall studies of those characteristics of living taxa which are most frequently found preserved on the fossils. Since the Gramineae had been selected for special emphasis in the study, I decided to study in considerable detail with SEM and light microscopy the anthoecia of species of the living genera that had been most closely related in the literature to the fossils, i.e., Piptochaetium, Nassella, Oryzopsis, and Stipa. Results of studies of modern forms are available elsewhere (Thomasson, 1976b, 1978b,c). Results of studies of the fossil forms are presented in the present paper. As a result of the studies, the phylogenetic relationships among the fossil and some living Gramineae, and especially in the Stipeae, have become clearer. Therefore, it is now appropriate to comment on these relationships.
While Elias (1942, 1946) emphasized the relationships of Berriochloa from the High Plains to the genus Piptochaetium, he nevertheless recognized affinities of Berriochloa to Stipa. He (1942) established a new section of the genus Stipa, Hesperostipa, based on the fossils and living species. Stebbins (1947) discussed the fossils and their related modern forms, relating the fossils primarily to the genus Piptochaetium. However, in the same paper he noted that "Stipa comata and S. spartea . . . may well be allopolyploids which have for one parent one of the 'Stipidium' species of the Pliocene flora." Apparently, he later (1975) revised this belief, considering S. comata and S. spartea to be Pleistocene migrants into North America. Williams (1975) has completely excluded the large and complex genus Stipa from consideration in the relationships of the fossils with modern taxa, a proposal which must be rejected as being too simplistic.
The genus Berriochloa has many anthoecial forms which are remarkably like those of the modern genus Piptochaetium. However, the shape of the anthoecia alone does not constitute sufficient evidence to support this assertion without intermediate ancestors. This is true for a number of reasons.
Species of Piptochaetium possess a characteristic grooved palea as well as a diagnostic lemma epidermal pattern. I have examined thousands of specimens of Berriochloa, and have never seen one with a distinctive grooved and keeled palea as found in Piptochaetium. Like Elias (1942), I have seen some Stipa, as well as some Berriochloa, with a medial depression in the palea, but never a groove. The groove and keels in Piptochaetium are not made "in the palea by the mutual pressure of the lemma and endosperm," as thought by Clements (personal communication to Elias, 1934), but rather are genetically determined. Almost equally diagnostic of the genus Piptochaetium is the system of lemma grooves and ridges created by the specialized epidermal cells. Again, none of the fossils examined have this characteristic. In fact the lemma epidermal patterns of most species of Berriochloa are strikingly similar to species of Stipa of the section Hesperostipa Elias, which contains the common North American species S. comata, S. spartea, and S. neomexicana. Finally, I have seen no species of Berriochloa with a blunt callus as found in many Piptochaetium. Even in species of Berriochloa where the callus is fragmentary, the fragments indicate development of a strong elongated callus. Such a callus, it would seem, could have little function if not sharp.
What can we say then about the relationships of the fossils to living genera? Undoubtedly Piptochaetium and Stipa, at least in part, had their origins in fossil ancestors from the High Plains. However the evolutionary route is complex and cannot be summarily explained by a comparison of the shape of the anthoecia. Rather a combination of both macro and microfeatures of the anthoecia must be utilized.
The lemmatal patterns in modern Stipeae seem to be characteristic of phylogenetic relationships (Thomasson, 1978b). The fact that the line of evolution represented by Nassella (Nassella pungens type) has been separate from other groups (i.e., Piptochaetium, many Stipa) for a considerable period of time is clearly indicated by lemmatal patterns of fossil and livimg species of the genus Nassella. Almost as clearly indicated by these studies is the position of the modern species of Stipa (i.e., Stipa robusta type) and Oryzopsis (Oryzopsis racemosa type) having lemma epidermal patterns approaching that of Nassella.
In species of the Stipa robusta type, silica bodies are extremely numerous, nearly one per long cell. In the living Nassella pungens type, silica bodies are entirely absent or very rare. Nevertheless patterns of greatly reduced long cells are clearly distinctive on both types. There appear to be intermediate lemmatal patterns between the Stipa robusta and Nassella pungens types. Species possessing these intermediate patterns include S. viridula and S. richardsonii. The Oryzopsis racemosa type is similar to the Stipa robusta type but the long cells are more developed and the silica cells less numerous.
Evidence for close relationships of species in the Stipa robusta type and at least some species of Oryzopsis is supported by evidence from sources other than lemmatal patterns. Oryzopsis hymenoides readily hybridizes with a number of species having the Stipa robusta type lemmatal epidermis (Johnson, 1945a, 1972; Johnson and Rogler, 1943). This would be expected on the basis of their lemmatal patterns. Combining evidences from hybridization and the study of lemmatal patterns it seems clear that species of Stipa of the Stipa robusta type are more closely related to lines of evolution represented by Nassella and Oryzopsis than to lines of evolution in species of Stipa representing the Stipa comata or Stipa capillata types. A single reported hybridization between Oryzopsis hymenoides and Stipa neomexicana (Weber, 1957) of the Stipa comata type does not seem likely, and in fact the evidence presented by Weber was, according to him, "circumstantial."
Johnson (1972) has postulated phylogenetic relationships in the Stipeae based on studies of cytology and geographic distribution. He indicated that the affinities of Oryzopsis hymenoides were puzzling, but that its geographic range and hybridization pattern suggest relationships with Stipa. The suggestion of these relationships is clearly supported by lemmatal patterns. A specimen of X Oryzopsis bloomeri examined had a Stipa robusta type lemma epidermis, which is also not unexpected, considering the hybrid nature of this taxon (Oryzopsis hymenoides X Stipa occidentalis).
As noted previously, lemmatal patterns of the fossil species of Berriochloa more closely resemble species of Stipa (e.g., Stipa comata type) than Piptochaetium. Unfortunately the intermediate evolutionary forms of Pliocene or Pleistocene age are not available. Undoubtedly such forms would answer the question about the origin of the groove in Piptochaetium as well as the characteristic lemma epidermal patterns. What can be said with good evidence at this point is that species of Piptochaetium species of Stipa of the Stipa comata type, and possibly others had a common origin in species of the fossil genus Berriochloa. At some time during the late Pliocene or Pleistocene, the evolution of these groups diverged. Their common ancestry is suggested by a basic similarity in lemmatal patterns (i.e., well-developed long cells, no short cells) and shapes of the anthoecia.
On the basis of this evidence I have postulated elsewhere that there appear to be at least two basic lines of evolution indicated by fossils (Thomasson, 1978b). In the Nassella-Oryzopsis-Stipa robusta type line the trend was towards a specialized epidermal pattern in which the long cells are greatly reduced and silica bodies are present or absent. This contrasts with the Berriochloa-Piptochaetium-Stipa comata line in which only long cells (or hooks) are present and are not reduced although they may be highly modified as in Piptochaetium. I have summarized a tentative phylogeny of the fossil and living Stipeae in Figure 8.
Figure 8--A proposed phylogeny of selected fossil and living Stipeae based on the epidermal patterns of the lemma. Small figures outside each box represent the general anthoecia shape of the taxon.
- Berriochloa primaeva
- B. maxima
- B. conica
- B. brevis
- Nassella pohlii
- Piptochaetium, avenaceoides, "Piptochaetium type"
- Stipa comata, "S. contata type"
- S. capillata, "S. capillata type"
- Oryzopsis racemosa, "O. racemosa type"
- Stipa robusta, "S. robusta type"
- Nassella pungens, "N. pungens type"
Systematic Descriptions
More than 24 months were expended in a search for the bulk of the Elias type collections. A significant number of these when found were inadequately labeled or not labeled at all and in some cases types had been lost or destroyed. Considerable time was spent in merely identifying and properly curating the collections. In many cases the collections of specimens are small and fragmentary. I have discussed at length the history of the type collections elsewhere (Thomasson, 1976b). In this publication the collections of Elias are indicated by the prefix E before a UNSM number, i.e., UNSM-E (=Elias) 50. All occurrences, unless otherwise noted, are from Ellis or Rooks (only site 4) counties, Kansas. Major collections of fossils recovered during this study are deposited at FHSM, ISC, UNSM, and ROM. Additional smaller collections are deposited in the Department of Geological and Geophysical Sciences, Princeton University and the Carnegie Museum, Pittsburgh.
Division Magnoliophyta
Class Liliopsida
Family Gramineae
Tribe Stipeae
Genus Berriochloa Elias
Berriochloa Elias, Univ. Kans. Sci. Bull. 33:346. 1932.
Stipidium Elias, Amer. J. Sci. 5th Ser. 29:26. 1935.
Diagnosis--An organ genus of the Gramineae represented by the silicified anthoecia (lemma and palea), rarely with portions of awns attached. Shape of the anthoecia fusiform, terete, oblong, obovate, or obtrullate. Surface of the lemma smooth, tuberculate, hirsute, or a combination of these at low magnification (ca 30x). At high magnification (ca 200+) epidermal patterns of long cells frequently visible. Long cells always longer than wide and with ripple walls; lobes of long cells simple, branched, or secondarily lobed. Hooks, prickles, or both present (see Metcalfe, 1960) surrounded and braced by expanded "buttress" lobes of long cells. No short cells were observed. Lemma narrowed at the summit forming a neck which may or may not expand into a crown. Two small lobes may be present in the front of the anthoecium at the summit. Portions of a quadrangular awn sometimes present. Scar of attachment of awn provided with 3 vascular trace scars, one central and two lateral. Lemma narrowed at the base into a sharp callus which is frequently covered with hairs. A single vascular trace scar present in the oblique lower portions of the callus. Palea enclosed within the lemma and overlapped by its margins. Surface of palea smooth or tuberculate; microfeatures of the palea epidermal cells like those of the lemma. Palea frequently terminating in a well-developed prow, this sometimes obscured by the summit of the lemma.
Discussion--I have reduced the genus Stipidium to a section under Berriochloa. In the section Stipidium I place all fossil species of Berriochloa which have cylindrical anthoecia. The other sections are Berriochloa and Parastipidium as described by Elias (1942). Although an initial macromorphological inspection of the fossils may suggest maintaining the taxonomic status quo, firm evidence can be demonstrated to justify my position.
The genus Berriochloa was originally established by Elias (1942) to include fossil grasses which he questionably referred to the tribe Hordeae. Anthoecia which were subsequently assigned to this genus by him were inflated and usually amphora-shaped. Elias had the fossils oriented upside-down at this time and it was not until later (1935) that he recognized their true orientation. He also established the genus Stipidium for cylindrical grass fossils which he initially assigned to the living genus Stipa. He gave the following very abbreviated generic description: "fossil grass closely related to the living Stipa or speargrass."
Not until 1942 did Elias describe and delimit more specifically Berriochloa and Stipidium. His separation of these genera was highly artificial at this time, and evidence he used in support of this separation is refuted both by the fossil record and by comparative studies of modern genera.
When Elias redefined the genera in 1942, he did it on the shape of the anthoecia. He stated that,
The difference between Stipidium and Berriochloa is only in the shape of the hull. It is cylindrical to oblong in Stipidium, while in Berriochloa it is fusiform, obovate, or resembles the Greek amphora. When Stipidium becomes proportionally shorter, it can still be well differentiated from Berriochloa if it preserves its cylindrical shape, thus looking like a barrel. But when its lower part assumes a gradual slope it becomes like the obovate Berriochloa.
He immediately noted that "some morphologists would be inclined to unite Stipidium and Berriochloa." He then reviewed those factors which he indicated supported his position.
Elias (1942) believed that the modern genera of the Stipeae were diagnostically different in their shape. Thus, all species of Stipa were considered by him to have cylindrical or oblong shaped anthoecia, while species of Piptochaetium and Nassella had fusiform or obovate-shaped anthoecia. Modern studies firmly refute this assumption as supporting the separation of Stipidium and Berriochloa. The genus Piptochaetium was shown by Parodi (1944) to be morphologically distinct from Stipa, Oryzopsis, and Nassella in the possession of a deeply furrowed palea. Although this distinction has long been recognized by South American agrostologists (Parodi, 1944; Rosengurtt, Arrillaga, and Izaguirre, 1970), the same cannot generally be said of their North American counterparts, including Elias. The shape of the anthoecia varies widely in the genus Piptochaetium, from cylindrical in P. avenaceum, P. avenaceoides (both North American), and P. hackelii (South American) to distinctly inflated obovate or nearly spherical in forms such as P. stipoides (Central and South American), P. lejopodum, and P. jubatum (both South American). In addition, there are many intermediate forms such as P. ruprechtianum, P. bicolor (both South American) and P. brevicalyx, P. pringlei, and P. fimbriatum (all North and Central American). More recently, I have shown that shape alone cannot be used to separate the genera of the Stipeae (Thomasson, 1976b, 1978b) and its use to separate the fossil genera Berriochloa and Stipidium is entirely artificial. To Elias's credit he recognized this artificiality, but unfortunately was relying on an interpretation of modern genera which is now proven false.
A recent paper by Williams (1975) contains parts which are quite erroneous and must be selected for comment and firm refutation. Williams (1975) indicates that the results of an analysis of morphological similarity between living and fossil material, "demonstrate that the criteria used by Parodi (1944) to separate the two sections of Piptochaetium successfully separate the fossil genera. Stipidium corresponds to Podopogon while Berriochloa corresponds to Eupiptochaetium." Further he states that, "Two examples of these affinities are first P. (Eupiptochaetium) fimbriatum (H.B.K.) Hitchcock from northern Mexico with Berriochloa minuta Elias from the uppermost Pliocene deposits of Kansas and Texas; and secondly, P. (Podopogon) bicolor (Vahl) E. Desvaux from Uruguay with Stipidium tubus var. laeve Elias of the middle Pliocene." The criteria which Parodi (1944) used were the following:- Podopogon (Rafin.)--Antecio glabro o piloso con pie punzante alargado y arista persistente.
- Eupiptochaetium--Antecio glabro o piloso con pie corto u obtuso y arista caedize o persistente.
I have examined the type collections of Elias, the collections of Leonard at the University of Kansas, and my own collections of perhaps 10,000 specimens of Berriochloa. In addition I have also examined more than 1,000 specimens of Piptochaetium including representatives of all North and Central American species and all but three of their types or type fragments, at least one specimen cited by Parodi (1944) of most species of South American Piptochaetium, and numerous other specimens as well. My research demonstrates that the dichotomy proposed by Williams is totally without basis in fact. There are no known fossil species of Berriochloa with a short blunt callus, which is the only preserved feature on the fossils that might be used to make the separation of the fossils into Parodi's sections. All fossil species of grasses which have been referred to those genera have a sharply pointed callus distinctly like that found in Parodi's Podopogon and many species of Stipa. Not only is Williams' proposal not defensible, but the examples he gives in illustration are in complete contradiction to his contentions. Berriochloa minuta Elias (see UNSM-E63(1) and UNSM-E67) is not comparable to Piptochaetium fimbriatum. Quite the contrary, Berriochloa minuta Elias is almost exactly like the varieties of Piptochaetium bicolor (Vahl) Desv. (see Mercedes, Mourino, Gallinal, Rosengurtt et al. PE 134 (US 1720569), cited by Parodi (1944), p. 256; Rosengurtt B-1122 (ISC 327795), cited by Parodi (1944), p. 253; among others). Berriochloa minuta Elias, like Piptochaetium bicolor (Vahl) Desv. has a very sharp, elongated callus. Stipidium tubus var. laeve Elias is not remotely similar to Piptochaetium bicolor (Vahl) Desv., but rather is comparable to a large number of species of Stipa and to Piptochaetium hackelii (Areck. sp) Parodi (see L. R. Parodi 80 (US 1127761), cited by Parodi (1944), p. 237), in the shape of the anthoecia and callus and little else. It should be noted that Williams has not seen the type collections of Elias of the fossil species mentioned and this may have contributed to his confusion. Finally, Williams fails to mention the genus Stipa in relation to the fossil genus Berriochloa. All fossil Berriochloa lack the distinct palea groove of Piptochaetium [very rarely a weak depression is developed (Pl. 1C)] and thus cannot be effectively separated from Stipa. Any consideration of the relationships of these fossil Stipeae is seriously remiss if the genus Stipa is not considered.
Elias also states that there seems to be an ecological difference between Stipidium and Berriochloa. Facts supporting this conclusion are insufficient at the present time.
Finally, Elias also reported that the position and nature of the labrum wart could be used to separate the genera Berriochloa and Stipidium. Although I have seen such a structure in extant Stipa spartea, I have not seen the distinction on the fossils mentioned by Elias in the collections I have studied. Elias only hesitantly mentioned the feature, since he rarely saw it, and I cannot find any evidence to support its existence as a diagnostic character.
Section Berriochloa
Berriochloa amphoralis Elias
(Pl. 1A, B; Pl. 8A)
Berriochloa glabra (Berry) Elias, Kans. Geol. Surv. Bull. 13:150. 1931. nom. nud.
Berriochloa glabra (Berry) Elias, in part. Sci. Bull.
Univ. Kans. 33: Pl. 29 Fig. 1 only. 1932.
Berriochloa amphoralis Elias, Geol. Soc. Amer. Sp. Paper 41:97. 1942.
Diagnosis--Anthoecia usually amphora-shaped or less commonly almost ovate. Outlines of upper anthoecia convex, never appearing angled. Maximum frontal convexity of the anthoecia usually at the same level or only slightly above that of the maximum rear convexity. Dimensions of anthoecia variable; length 2.0-5.7 mm, depth 1.4-3.2 mm, width 1.5-2.9 mm. Distance from rear of crown to dorsal rib 0.6-1.2 mm. Lemma narrowed towards the summit into a neck which terminates in a distinctive expanded crown to which an awn was apparently attached. Rim of crown smooth, inner diaphragm with three vascular trace scars, the center scar larger than the lateral two. From above, crown-shape obovate, 0.5-1.2 mm long, 0.5-0.9 mm wide. The point where the rear of the lemma enters the callus is designated by the term "heel." In Berriochloa amphoralis the heel of the lemma is well developed, usually abruptly angled at union with the callus. Lemma narrowed towards the base into a strong, elongated callus, only fragments of which have been preserved. Surface of the lemma smooth at low magnifications except for scattered hooks on the shoulder and neck and, in the best preserved specimens, dense hairs present on the lower 1/3-1/2 of the lemma, sometimes also on the margins. At high magnifications, vertical rows of long cells visible. Palea well developed, its margins overlapped by the lemma; summit of the palea terminating in a well-developed prow. Surface of palea weakly to evidently covered with hooks, never smooth.
Nomenclature--The type specimens of this species are a collection now in UNSM as UNSM-E78. Specimen UNSM-E78-L is designated as the lectotype.
Occurrences--(1) In the upper exposed portion of bed 3, at locality 2, site 5. (2) In the limited exposure at site 14. (3) In the lower 25 cm of sandy tuff in bed 2 at site 13. (4) In the lower 60 cm of bed 3 and the lower 25 cm of bed 5 at site 21. (5) In the lower 1/2 of bed 1 at site 22. (6) 50 cm above the contacts of beds 1 and 2, in bed 2 at site 31. (7) Several clusters were recovered in the upper 60 cm of bed 5 at site 42.
Discussion--The specimens of this species are very variable in size and general shape. The largest specimens approach in size those of B. tuberculata and more rarely B. maxima. When this occurs, the smoothness of the lemma is the distinguishing feature. At high magnification the epidermal pattern of B. amphoralis appears distinct from that of B. maxima. In B. maxima, the lobes of the long cells are secondarily lobed or branched (Pl. 8E, F), whereas both B. amphoralis and B. tuberculata have lobes only rarely secondarily lobed or branched (Pl. 8A; 9D). Berriochloa amphoralis can be distinguished from B. tuberculata by the complete absence of hooks or hairs on the medial 2/3 of the lemma surface. When the largest specimens of B. minuta approach the smallest of B. amphoralis, the former are usually distinguished by their lesser depth. Usually the shape is also distinctive, the outline of B. minuta being more angular. Where single specimens of either are collected, it may not be possible to assign a species name.
Berriochloa conica Elias
(Pl. 8B, C)
Berriochloa conica Elias, Geol. Soc. Amer. Sp. Paper 41:95. 1942.
Diagnosis--As given by Elias (1942) with modifications of the dimensions and the addition of new micromorphological data. Length of anthoecia, excluding callus, 2.1-3.7 mm, width 1.1-1.7 mm, depth 1.1-1.6 mm; distance from rear of crown to dorsal rib 0.4-0.7 mm; crown length 0.3-0.4 mm, width 0.3-0.5 mm. Average length to depth ratio of 3 samples 1.9-2.1. Epidermis of lemma at high magnification characterized by long cells with well-developed lobes. Lobes simple and expanded at the apex. Transverse as well as longitudinal walls lobed.
Nomenclature--The types of this species are in UNSM as UNSM-E77. Specimen UNSM-E77-L is designated as the lectotype.
Discussion--This species is closest to B. minuta but differs in the proportionally larger size. See discussion of B. minuta in Elias (1942) and this paper.
Berriochloa dawesensis (Elias) Thomasson, comb. nov.
Stipidium ∝ ∝ Chaney and Elias, Carnegie Inst. Wash.
Publ. 476:78. 1938.
Stipidium dawesense Elias, Geol. Soc. Amer. Sp. Paper 41:79. 1942.
Diagnosis--As given in Elias (1942). Poorly preserved long cells with well-developed (? simple) lobes were observed at high magnification.
Nomenclature---The type specimens of Berriochloa (Stipidium) dawesensis are in UNSM as UNSM-E24. Specimen UNSM-E24-L is designed as the lectotype.
Discussion--The type collection is quite fragmentary and the reader is referred to UNSM-E17 for a well-preserved collection of this species. See also Galusha (1975) for a new interpretation of the stratigraphic occurrence of Berriochloa dawesensis.
Berriochloa glabra (Berry) Elias
Lithospermum fossilium Berry var. glabrum Berry, Proc. U.S. Nat. Mus. 73 (2734): 2; pl. 1, Figs. 11 and 12 only. 1928.
Diagnosis--As given by Elias (1942, pp. 93-94; pl. 12, Figs. 10-13) except that Fig. 13 of Berry (1928b) is now excluded. The anthoecia measurements are modified as follows: length 2.9-6.1 mm; depth 1.5-3.1 mm; width 1.3-2.9 mm. Distance from rear of crown to dorsal rib, 0.7-1.3 mm.
Nomenclature--I have carefully examined all the specimens from the USNM including the specimens shown by Berry in 1928b and those by Elias in 1942. The collections were found by me to be a mixture of species. I now restrict the species B. glabra to those specimens illustrated by Berry (1928b) as Fig. 11 (now USNM 38073) and Fig. 12 (now USNM 22718). Both specimens show clearly the diagnostic character of this species--the distinctly elongated neck. The specimen illustrated by Berry (1928b) in Fig. 13 (now USNM 22719) belongs to an undescribed species of Berriochloa similar to B. amphoralis. I designate as lectotype the specimen (USNM 22718) figured by Berry in 1928b on Plate 1 as Fig. 12.
Occurrences--No representatives of this species were collected during this study.
Discussion--Even though the specimens of this species are quite variable in size, they are readily identified by the elongated neck.
Berriochloa inflata Elias
(Pl. 1C, Pl. 8D)
Berriochloa inflata Elias, Geol. Soc. Amer. Sp. Paper 41:96. 1942.
Diagnosis--As given by Elias (1942) except for the addition of micromorphological characteristics. At high magnification surface of lemma with long cells possessing well-developed lobes; lobes compound, usually dichotomous.
Nomenclature--The type specimens when found were in a vial with a label that read on one side "Berriochloa lemma and palea smooth. Other with tuberculate palea" and on the reverse "center sec. 34, T. 15, R. 27 W. Panicum bed." A portion of the type collection of Berriochloa inflata is at UNSM as UNSM-E14. Specimen UNSM-E14-L is designated as lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--See also Elias (1942). These specimens are very fragmentary and a new collection is needed. The pattern of the epidermal cells of the lemma is unique if it holds for all members of this taxon. The only other Berriochloa with nearly the same shape and a smooth palea is B. obtrullata, but B. obtrullata is much larger.
Berriochloa intermedia (Elias) Thomasson, comb. nov.
Stipidium intermedium Elias, Geol. Soc. Amer. Sp. Paper 41:83. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--The type collection is now at UNSM as UNSM-E1. Specimen UNSM-E1-L is designated as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--Also see Elias (1942). Although I have made a number of trips to the Reamsville area, I have been unable to relocate the type collection site. Local ranchers and farmers indicate that no exposure of rock to their knowledge was ever present at the locality given in Elias (1942) and that locality can probably be considered in error.
Berriochloa maxima Elias
(Pl. 8E, F)
Berriochloa maxima Elias, Geol. Soc. Amer. Sp. Paper 41:98. 1942.
Diagnosis--As given by Elias in 1942, with the addition of micromorphological data. Long cells well developed with irregularly branched lobes.
Nomenclature--The syntypes of B. maxima are Elias specimens collected from the Panicum elegans subzone of the Ash Hollow Formation, Ogallala Group, 3/4 mile north of Wolback, on highway to Spalding, Greeley County, Nebraska (Elias, 1942). Though I have made extensive searches, I have been able to locate only one of the specimens shown by Elias. The specimen (1942, pl. 14, Fig. 3) is a mere fragment. Only this single specimen from UNSM-E23 can be considered a type specimen. Specimens which now best illustrate at least some of the characteristics of this taxon are those in UNSM-El3.
Occurrences--(1) In the lower 20 cm of bed 2 at site 9b (= bed 5 at 9a). (2) In the lower 30 cm of bed 3 at site 5. (3) In the uppermost exposed beds at site 30. (4) In the uppermost exposed beds at site 10. (5) In the lower 100 cm of bed 4 at site 13.
Discussion--The fragmentary nature of all the reference collections in my possession makes the recognition of this species difficult. However, the large size of the anthoecia of this species, for the most part, sets it aside from all other species of Berriochloa. Berriochloa (Stipidium) grandis is perhaps the nearest in size but differs in the larger size of the anthoecium, and in the more distinctly cylindrical shape. Rarely, the largest specimens of B. tuberculata approach the smallest of B. maxima in size and where this is the case it is necessary to use samples rather than individuals to determine the correct identity. Most of the specimens of B. maxima recovered in Ellis County are poorly preserved. Undoubtedly B. maxima and B. tuberculata are closely related, and this factor must be considered when arriving at a "species" identification when the characteristics of each overlap. This taxon is in need of a thorough restudy including collection of well-preserved specimens with strict stratigraphic control.
Berriochloa minuta Elias
(Pl. 1D, E, F; Pl. 9A)
Berriochloa minuta Elias, Geol. Soc. Amer. Sp. Paper 41:96. 1942.
Diagnosis--As given in Elias (1942) but with the following modifications in dimensions and the addition of new information on micromorphological structures. Length of anthoecia 1.3-2.8 mm, width 0.9-1.8 mm, depth 0.9-1.8 mm; distance from rear of crown to dorsal rib 0.3-0.9 mm; crown length 0.3-0.6 mm, width 0.3-0.5 mm; length/depth ratio of 14 samples 1.5-1.8. At high magnifications surface of epidermis with long cells having well-developed lobes. Lobes simple, slightly or evidently expanded at end, apex obtuse.
Nomenclature--The types of this collection are at UNSM as UNSM-E76. UNSM-E63(2) is another collection which was cited in the protologue by Elias (1942).
Occurrences--(1) In the lower 30 cm of bed 2 at locality 2, site 30 as cluster T, 5 July 1975. (2) In the upper 1/4 of bed 2 at site 8 as clusters 3(=C), 6(=F), and 7(=G) 10 August 1975 (inadvertently not shown on Pl. I, graphic section 8). (3) At site 9a (= locality 1a and 1b) approximately 8.0 cm above the contacts of bed 3 and 4 in bed 4 as cluster A, 23 December 1974; at site 9a approximately 90 cm below the contacts of beds 2 and 3 in bed 2. At site 13 in the lower meter of bed 4, locality 1 as cluster A, 4 August 1973. (5) At site 22 in the middle of bed 1 as cluster 1(4), 27 May 1974. (6) In upper bed 1 about 2.2 meters below the contacts of bed 1 and 2 at site 33 as cluster B, 28 December 1974. (7) At site 39 in lower bed 4 at locality 1, as cluster K, 7 July 1975; and in the upper 14' of bed 2 at locality 3 as cluster E, 9 July 1975. (8) At site 42 in the upper meter of bed 6 as clusters K and D, 3 August 1975.
Discussion--Berriochloa minuta is variable in size but separable from closely-related forms. From B. conica, to which it is the most similar, it differs in a smaller average length and consequently smaller length-to-depth ratio. While there is overlapping, most samples are distinct. It may not be possible however to place individual specimens securely in either taxon where this occurs. The smallest specimens of B. amphoralis may be similar to the largest specimens of B. minuta but the depth in B. amphoralis is almost always proportionally greater. Specimens of B. tuberculata which are as small in length as those of B. minuta differ in their hooked or hirsute lemma surface and proportionally greater depth.
Berriochloa obtrullata sp. nov.
(Pl. 1G; Pl. 2A, B)
Diagnosis--Shape of anthoecia obtrullate in frontal view; length 2.2-3.2 mm, depth 1.7-2.4 mm; width 1.6-2.4 mm; distance from rear of crown to dorsal rib 0.9-1.3 mm; well-developed, expanded crown present at apex of lemma; crown length 0.4-0.7 mm, width 0.4-0.6 mm; callus not recovered. Surface of lemma smooth, except for scattered hooks on a well-developed shoulder and neck. Palea smooth in the middle 12, with hooks at the margins; palea terminating in a well-developed prow which is usually enclosed by the margins of the lemma. Awn scar with three vascular trace scars, one large and medial, two lateral and smaller.
Type locality and horizon--In bed 2 which is exposed as a distinct "mortar" bed, locality 3c, at site 30, SE SW sec. 9, T. 12 S., R. 18 W., Ellis Co., Kansas.
Additional Occurrences--(1) Possible specimens of this species were found in poorly exposed strata at site 24.
Discussion--This species is closest to B. amphoralis but differs in the smooth palea and the distinctly obtrullate shape which creates a well-defined, almost platform-like shoulder. It also resembles the species described by Elias as B. inflata which also has a smooth palea but the size of B. inflata as indicated by Elias is smaller than B. obtrullata.
Holotype collection--Holotype, Thomasson SEM No. BP172, 15 August 1974 as cluster A in Sternberg Memorial Museum (FHSM); isotypes Thomasson No. 30.1, 30.2, 30.3, and 30.4 in FHSM, ISC, UNSM, and ROM.
Berriochloa primaeva Elias
(Pl. 9B)
Berriochloa primaeva Elias, Geol. Soc. Amer. Sp. Paper 41:94. 1942.
Diagnosis--As given in Elias (1942) with the addition of micromorphological characters of the lemma. Long cells of lemma at high magnification with well-developed lobes; lobes simple, their tips acute, only very rarely expanded.
Nomenclature--The types of this species are in UNSM as UNSM-E9. Specimen UNSN4-E9-L is designated as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942).
Berriochloa pumila sp. nov.
(Pl. 2C, D, E; Pl. 9C)
Diagnosis--Sbape of anthoecia obconical to obovate; length excluding callus 1.5-2.8 mm, depth 0.9-1.3 mm, width 0.8-1.3 mm; distance from rear of crown to dorsal rib 0.3-0.5 mm; lemma with an expanded crown at the summit; crown 0.3-0.5 mm long, 0.3-0.5 mm wide; lemma tapering at the base, ending in a sharp-pointed hirsute callus 0.4-1.0 mm long. Surface of lemma at low magnifications with abundant hooks, these frequently absent at the margin of the lemma toward the base. At high magnification surface of lemma with long cells possessing well-developed lobes. Lobes simple, not expanded at the apex. Palea tuberculate, terminating in a weakly-developed prow which is usually concealed by the margins of the lemma.
Type locality and horizon--From 1 meter below the contacts of beds 2 and 3, in bed 2, at site 9a exposed in a roadcut along the east side of federal-aid-secondary highway (FAS) 231 (Fig. 7; Pl. 1, graphic section 9a) sec. SW NW NW 3, T. 12 S., R. 20 W., Ellis Co., Kansas, as cluster B, 1 August 1974.
Additional Occurrences--(1) At approximately the middle of bed 1 at site 8. (2) In the upper 1/2 of bed 2 at site 8 as cluster 7(=G) and 6(=F) 10 August 1974. (3) In the upper 20 cm of the bed 1 (= bed 5 at 9a) at site 9b (=locality 2). (4) In the small exposure at site 14. (5) In the middle of bed 1 at site 22 as cluster 1(2) 27 May 1974. (6) In the upper 1/2 of bed 1 at site 28 as cluster B, 26 July 1974. (7) At site 33, 2.2 meters below the contacts of bed 1 and 2, in bed 1, as cluster B(1) "seeds" 12 June 1975. (8) In the upper 1/4 of bed 2 at locality 3, as cluster E, 9 July 1975 and in the lower 1/4 of bed 4 at locality 1 as cluster D, 7 July 1975, both at site 39.
Discussion--This species is most similar to B. variegata var. dartoni. However it differs in the proportionally smaller size.
Holotype collection--Thomasson SEM No. BP137, 1 August 1974, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 9.12, 9.13, 9.16, and 9.17 in FHSM, ISC, UNSM, and ROM.
Berriochloa sternbergii sp. nov.
Lithospermum fossilium var. aristatum Berry, Proc. U.S. Nat. Mus. 73 (2734):2. In part only, specimens not shown by Berry, 1928.
Stipidium aristatum (Berry) Elias, Geol. Soc. Amer. Sp. Paper 41:85. Only pl. 10, Fig. 5.
Diagnosis--As in Elias (1942) but the dimensions are modified as follows: Length of anthoecia 5.2-6.5 mm; depth 1.1-1.7 mm; width 1.3-1.7 mm. Distance from rear of crown to dorsal rib 0.2-0.3 mm.
Nomenclature--This species is based on specimens referred to by Berry (1928b) under Lithospermum fossilium var. aristatum but not illustrated. The illustration of this new species is pl. 10, Fig. 5 of Elias (1942). Specimens in pl. 10, Fig. 6 are in my opinion too fragmentary to be identified. All specimens are now at USNM. These specimens are most similar to Berriochloa (Stipidium) nebraskensis. The collection of B. nebraskensis is very poor, consisting of one whole specimen and two fragments. Nevertheless, the whole specimen does have a smooth palea, whereas the palea of Berriochloa sternbergii is tuberculate.
Holotype collection--John B. Hatcher, Phillips Co., Kansas, 1884, now in USNM.
Discussion--See section on discussion of Berriochloa aristata. This species is named in honor of Charles H. Sternberg who originally discovered the beds from which these fossils probably come.
Berriochloa tuberculata Elias
(Pl. 2F, G; Pl. 9D)
Berriochloa tuberculata Elias, Geol. Soc. Amer. Sp. Paper 41:98. 1942.
Diagnosis--As in Elias (1942) with the following changes or additions: Anthoecia length excluding callus 1.7-6.6 mm, width 1.2-3.4 mm, depth 1.3-3.4 mm; mean length/depth ratio of 11 measured samples 1.2-1.7 mm. Surface of lemma at low magnification, with hooks or hairs or both. At high magnifications long cells with well-developed lobes are observed. Lobes mostly simple and expanded at the tip, or becoming buttress lobes around hooks. Surface of palea with hooks and hairs.
Nomenclature--The type collection of this species is at UNSM as UNSM-E75. Specimen UNSM-E75-L is designated as the lectotype. Another reference collection is UNSM-E74. This is a collection of B. tuberculata and was illustrated by Elias (1942, pl. 13, Fig. 5).
Occurrences--(1) In the upper 10 cm of bed 3 at site 1. (2) Site 2: lower bed 2 locality 1, 25 cm above base of bed as cluster T, 5 July 1975; a mass occurrence (? coprolite) (Fig. 5A) in the upper 1/3 of bed 2; specimens unrelated to the ? coprolite were common throughout bed 2. (3) Clusters of specimens with excellently preserved hairs and hooks were recovered in the lower 1/2 of bed 2 at locality 1, site 5. (4) In the upper 1/3 of bed 2 at site 8 as cluster 6 (=F) 10 August 1974. (5) Clusters and individuals were common throughout bed 4 at site 9a (= localities 1a and 1b), and in the exposed portion of the equivalent bed at site 9b (= locality 2). (6) In the upper 100 cm of bed 2 at locality 1, site 10. (7) In the small exposure at site 14. (8) In the middle of bed 1 at site 22. (9) In the small exposure at site 24. (10) About 150 cm above the contact of the Tertiary and Cretaceous deposits at site 27. (11) About the middle of bed 1 at site 28. (12) In the silo pit (locality 1) in the upper portions of bed 1 at site 30. (13) In bed 8 at site 32. (14) In the upper 1/3 of bed 1 at site 33. (15) In the lower 1/4 of bed 4 at locality 1, site 39. (16) In the upper 10 cm near the contact of beds 5 and 6 in bed 6 at site 42 as clusters Q and O, 3 August 1975. (17) In the lower 1 meter of bed 1, at site 40, Smith County, Kansas.
Discussion--Specimens of B. tuberculata are extremely variable both in size and surface features. In the smallest specimens they overlap in size with B. amphoralis and B. minuta. From both however they are readily distinguished by the hirsute-hooked surface of Berriochloa tuberculata. The lemma surface of both B. minuta and B. amphoralis is usually smooth although scattered hooks may be present on the shoulder and hairs may be present on the callus. But hairs or hooks were never observed on the medial 2/3 of either B. minuta or B. amphoralis. It is not unusual that the loss of the cuticle and hair bases or processes of fossilization may obscure the true relations of a particular sample. In this case only, tentative affinities can be ascribed, i.e., B. c.f. tuberculata-amphoralis. The depth of the anthoecia in B. tuberculata is usually proportionally greater than in B. minuta but about the same as in B. amphoralis.
Berriochloa variegata (Elias) Thomasson, comb. nov.
Stipidium variegatum Elias, Geol. Soc. Amer. Sp. Paper 41:82. 1942.
Diagnosis--As given by Elias (1942) with the following additions: Depth of anthoecia 0.9-1.4 mm; distance from rear of crown to dorsal rib 0.3-0.6 mm; crown length 0.4-0.7 mm, width 0.4-0.6 mm.
Nomenclature--The types of this species are in the UNSM as UNSM-E8. Specimen UNSM-E8-L is designated as the lectotype.
Occurrences--(1) At site 9b in the upper 25 cm of bed 1. (2) In the middle of bed 1 at site 22. (3) About 1.5 meters above the base of bed 4, at locality 1, site 39.
Discussion--Although the type specimens are very well preserved, they are very few in number. As a result the species is very poorly defined. Unfortunately, I was unable to make new collections at the type locality, a necessity if this important taxon is to be accurately defined. Collections at KANU as KANU 9603 and KANU 9774 do not appear to be this taxon although they were so labeled by me early in this study. The Ellis County fossils are assigned to this taxon only in light of these previous considerations. Also see Elias (1942) and discussion of B. pumila in this paper.
Berriochloa variegata (Elias) Thomasson, var. dartoni (Elias) Thomasson, var. nov.
Stipidium variegatum Elias, modf. dartonii Elias, Geol. Soc. Amer. Sp. Paper 41:83. 1942.
Diagnosis--As given in Elias (1942) with the following additions: Depth of anthoecia 1.1-1.5 mm; distance from rear of crown to dorsal rib 0.4-0.6 mm; crown length 0.5-0.7 mm, width 0.4-0.5 mm; callus length to 1.2 mm.
Nomenclature--The type collection is now at UNSM as UNSM-E6. These specimens were illustrated by Elias (1942, pl. 11, Figs. 6-8). Specimen UNSM-E6-L is designated as the lectotype. I have excluded fragments of an unknown (but not B. variegata var. dartonii) Berriochloa sp.
Occurrences--No representatives of this taxon were collected during this study.
Discussion--See Elias (1942) and discussion of B. pumila in this paper.
Section Stipidium Elias
Berriochloa aristata (Berry) Elias
Lithospermum fossilium var. aristatum Berry, Proc. U.S. Nat. Mus. 73 (2734):2, pl. 1, Fig. 14 only. 1928.
Berriochloa ? aristata (Berry) Elias, Univ. Kans. Sci. Bull. 20(20):349. 1932.
Diagnosis--Anthoecium narrowly obovate; length 8.4 mm; depth ≈ 2.7 mm (broken); width 2.8 mm. Lemma narrows at the summit, terminating in a short neck and expanded crown. Crown viewed from above ovate, 1.1 mm long; 1.0 mm wide. Distance from rear of crown to dorsal rib 1.0 mm. Neck and shoulder finely tuberculate, hairs present on the lower lemma near the callus. Remainder of lemma smooth. Palea well developed, terminating in a prominent prow; surface of palea finely tuberculate. A shallow medial depression runs the length of the palea. Callus well developed, 5.1 mm long, terminating in a broken end.
Nomenclature--See section on nomenclature of Berriochloa sternbergii sp. nov. The single specimen illustrated by Berry (1928b) in Pl. 1, Fig. 14 (now USNM 38074) is designated as the lectotype.
Discussion--These specimens were part of a mixture of two taxa. Berry (1928b) described the mixture as a single species and Elias (1942) continued this practice. I now restrict the name Berriochloa (Stipidium) aristata to the single specimen illustrated by Berry (1928b) and referred to by Elias (1942).
Berriochloa aspera sp. nov.
(Pl. 3B, C, D)
Diagnosis--Anthoecia oblong; length, excluding callus 2.8-4.7 mm long, depth 0.6-1.1 mm, width 0.71.2 mm; distance from rear of crown to dorsal rib 0.1-0.2 mm; apex of lemma with expanded crown; length of crown 0.4-0.6 mm, width 0.5-0.6 mm; callus well developed, greatest length 1.6 mm. Surface of lemma at low and high magnification characterized by very coarse hooks over the entire surface except near the base in the front. Palea tuberculate, terminating in a very strong prow which projects forward from the anthoecia at about a 45° angle.
Type locality and horizon--In the upper 1/4 of bed 1 at the sand pit which is site 22 at NW SW sec. 31, T. 13 S., R. 19 W., Ellis Co., Kansas.
Discussion--This species is closest to B. coloradoensis and some specimens of B. variegata. However it differs from both in the very coarse hooks and the stout, projecting prow of the palea. The nature of the projecting palea is similar to the condition found in the living North American species of Stipa, S. comata, S. spartea, and S. neomexicana. The size of the crown is proportionally larger in this species with the crown length nearly equaling the anthoecia depth.
Holotype collection--Holotype, Thomasson SEM No. BP162, 28 May 1974, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 22.4, 22.5, 22.6, and 22.7, 28 May 1974 in FHSM, ISC, UNSM, and ROM.
Berriochloa brevis (Elias) Thomasson, comb. nov. (Pl. 3E; Pl. 9E)
Stipidium breve Elias, Geol. Soc. Amer. Sp. Paper 41:81. 1942.
Diagnosis--As given by Elias except for modifications of measurements and newly discovered micromorphological information. Width of anthoecia 0.91.0 mm, depth 0.7-1.0 mm; distance from rear of crown to dorsal rib 0.2-0.3 mm; crown length 0.4 mm, width 0.4-0.5 mm. Surface of lemma at high magnification with long cells and hooks. Long cells sinuous with well-developed lobes; lobes generally compound, length of lobes always greater than the width of the central area of the cells; buttress lobes well developed around hooks.
Nomenclature--The type specimens of this species are in the UNSM as UNSM-E73. The specimens were illustrated by Elias (1942, Fig. 1, plate 11). Specimen UNSM-E73-L is designated as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--This species is most similar to Berriochloa nova, but B. nova is also proportionally longer than B. brevis. In addition, B. nova has hooks over the entire surface of the lemma, while B. brevis does not. See also Elias (1942).
Berriochloa coloradoensis (Elias) Thomasson, comb. nov.
Stipidium coloradoense Elias, Geol. Soc. Amer. Sp. Paper 41:80. 1942.
Diagnosis--As in Elias (1942), with the addition of the following micromorphological characters: Lemma with abundant hooks grading into broad-based elongated prickles near the summit. "Shadow" of epidermal cells can be seen indicating long cells with probably compound lobes.
Nomenclature--The type collection of this species, which is poor, is at UNSM as UNSM-E11. These specimens were illustrated by Elias (1942, pl. 8, Figs. 3, 4). Specimen UNSM-E11-L is designated as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--This collection is very poor. Another collection is necessary to determine whether this is a "good" species or only fragments of a previously described one.
Berriochloa communis (Elias) Thomasson, comb. nov.
Stipidium commune Elias, Geol. Soc. Amer. Sp. Paper 41:80. 1942.
Diagnosis--As given in Elias (1942), but modified as follows: length of anthoecia 2.3-5.5 mm, depth 0.6-1.2 mm, width 0.7-1.3 mm; distance from rear of crown to dorsal rib 0.1-0.4 mm; crown length 0.3-0.6 mm, width 0.3-0.6 mm; callus length 0.6-1.8 mm. The lemma and palea at high magnification show a "shadow" pattern of long cells. Lobes appear to compound, but this is uncertain. Well-developed hooks present on both lemma and palea.
Nomenclature--The type specimens are in the UNSM as UNSM-E72. Specimen UNSM-E72-L is designated as the lectotype. Two other collections of B. commune that were studied are the following: (1) A collection which is now UNSM-E33. These specimens were illustrated in part, by Elias (1942, pl. 7, Fig. 4). (2) Another collection is now UNSM-E29. These specimens were also illustrated by Elias (1942, pl. 9, Figs. 3-7), as "autotypes." Since both these collections of specimens were referred to by Elias in his protologue, they might be regarded as paratypes.
Occurrences--No representatives of this species were collected during this study.
Discussion--New collections of this species with strict stratigraphic controls are needed since it occurs at some of the most significant vertebrate quarries in the High Plains. A fossil mentioned earlier by Chaney and Elias (1938) as Stipidium ∝ is Berriochloa communis.
Berriochloa elongata (Elias) Thomasson, comb. nov.
Stipidium elongatum Elias, Geol. Soc. Amer. Sp. Paper 41:84. 1942.
Diagnosis--As given in Elias (1942) but dimensions modified as follows: Length of anthoecia 4.8-5.4 mm, width 0.7-1.4 mm, depth 0.6-1.4 mm; distance from rear of crown to dorsal rib 0.1-0.3 mm; crown length 0.4-1.0 mm, width 0.5-1.0 mm; callus length 1.1-1.5 mm.
Nomenclature--The types of Berriochloa elongata are at UNSM as UNSM-E25. These were illustrated by Elias (1942, pl. 10, Fig. 1). Specimen UNSM-E25-L is designated as the lectotype.
Discussion--As given in Elias (1942). This species differs only slightly from Berriochloa nebraskensis but the collection of B. nebraskensis is of such small size and poor quality that a differentiation is doubtful.
Berriochloa grandis (Elias) Thomasson, comb. nov.
(Pl. 9F)
Stipidium grande Elias, Geol. Soc. Amer. Sp. Paper 41:86. 1942.
Diagnosis--As given by Elias (1942). Lemma surface at high magnification with long cells possessing well-developed lobes; lobes simple or compound, always longer than the width of the central area of the cells.
Nomenclature--The type collection is at UNSM as UNSM-E10. These specimens were illustrated by Elias (1942, pl. 14, Fig. 5). Specimen UNSM-E10-L is designated as the lectotype. A paratype collection is also in UNSM as UNSM-E71. This is the "smaller variety" of Elias as comparison of the collection with Elias's illustrations shows (Elias, 1942, pl. 14, Fig. 6).
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942).
Berriochloa kansasensis (Elias) Thomasson, comb. nov.
Stipa kansasensis Elias, Univ. Kans. Sci. Bull. 20:344. 1932.
Stipidium kansasense Elias, Geol. Soc. Amer. Sp. Paper 41:81. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--The type collection is very fragmentary, but a comparison with the illustrations confirms these are the "cotypes" designated by Elias. The collection is now in UNSM as UNSM-E70.
Occurrences--No representatives of this species were collected during this study.
Discussion--Also see Elias (1942). This collection is extremely poor.
Berriochloa minima (Elias) Thomasson, comb. nov.
Stipidium minimum Elias, Geol. Soc. Amer. Sp. Paper 41:77. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--The type collection of this species is at UNSM as UNSM-E69. These specimens were illustrated as syntypes by Elias (1942, pl. 6, Figs. 1-4). I have designated UNSM-E69-L as the lectotype and the remainder may be considered paratypes.
Discussion--See Elias, 1942.
Berriochloa naruhea sp. nov.
(Pl. 3F, G; Pl. 10A)
Diagnosis--Shape of anthoecia oblong; length, excluding callus, 2.7-5.5 mm, depth 0.8-1.3 mm; distance from rear of crown to dorsal rib 0.2-0.4 mm; lemma terminating in an expanded crown; crown length, 0.4-0.6 mm, width 0.4-0.5 mm; fragments of sharp callus to 1.1 mm; awn fragments square in cross section, 0.3-0.4 mm thick; fragments recovered bad a maximum length of 1.0 mm, whole awns were apparently much longer. Surface of lemma at low magnification with hooks usually covering the entire surface but sometimes restricted to as little as upper 3/4, the base and margins of the lemma in front often smooth. High magnification reveals sinuous long cells with well-developed lobes; lobes simple or slightly compound, tips of lobes obtuse, not usually expanded, buttress lobes strongly developed around hooks. Palea well developed, terminating in an evident prow; surface of palea with hooks and at high magnification epidermis like that of the lemma.
Type locality and horizon--In the upper 10 cm of bed 1 at site 4, NE NE sec. 26, T. 10 S., R. 18 W. Rooks Co., Kansas.
Discussion--This species is very similar to Berriochloa communis. However, the width and depth are proportionally greater in B. naruhea. In B. naruhea the lemma is more completely covered with hooks which are also coarser than in B. communis. The specific epithet of B. naruhea is a contrived name, of no scientific significance.
Holotype collection--Holotype, Thomasson SEM No. BP157, 28 May 1973, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 4.10, 4.11, 4.12, and 4.13, FHSM, ISC, UNSM, and ROM.
Berriochloa nebraskensis (Elias) Thomasson, comb. nov.
Stipidium nebraskense Elias, Geol. Soc. Amer. Sp. Paper 41:85. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--The type collection is now at UNSM as UNSM-E5. I have selected UNSM-E5-L as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942). Also see discussion of Berriochloa elongata in this paper.
Berriochloa nova (Elias) Thomasson, comb. nov.
Stipidium novum Elias, Geol. Soc. Amer. Sp. Paper 41:87. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--The type specimens of Berriochloa nova are now at UNSM as UNSM-E2. These specimens were illustrated by Elias (1942, pl. 8, Fig. 1). I have designated UNSM-E2-L as the lectotype. This collection was mixed, containing some specimens of Berriochloa primaeva. I have excluded the latter.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942). Also see discussion of Berriochloa brevis in this paper.
Berriochloa tuba (Elias) Thomasson, comb. nov. (Pl. 10B)
Stipidium tubus Elias, Geol. Soc. Amer. Sp. Paper 41:84. 1942.
Stipidium tubus Elias var. laeve Elias, Geol. Soc. Amer. Sp. Paper 41:84. 1942. nom. nud.
Diagnosis--As given by Elias (1942).
Nomenclature--The type specimens are now at UNSM as UNSM-E67. Comparison of the specimens with the illustrations of Elias (1942, pl. 10, Fig. 4) confirms these as the type specimens of Berriochloa tuba. I have designated UNSM-E67-L as the lectotype. The variety designated by Elias as Stipidium laeve var. laeve is placed in synonymy with B. tuba. The specimens of this taxon are at UNSM as UNSM-E68.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942).
Section Parastipidium Elias
Berriochloa hirsuta (Elias) Thomasson, comb. nov.
Stipidium hirsutum Elias, Geol. Soc. Amer. Sp. Paper 41:89. 1942.
Diagnosis--As given in Elias (1942).
Nomenclature--There are two collections of fossils at UNSM which by comparison with the illustrations of Elias (1942) are syntypes. Collection UNSM-E4 are those specimens designated by Elias which come from Pliohippus draw and are illustrated on pl. 6, Fig. 5A (Elias, 1942). The second designated by Elias as syntypes is UNSM-E27. These are the specimens illustrated on pl. 6, Fig. 5 (Elias, 1942). 1 have designated UNSM-E27-L as the lectotype. The remainder of both collections should be considered paratypes.
Occurrences--No representatives of this species were collected during this study.
Discussion--See also Elias (1942). Both UNSM-E4 and UNSM-E27 were originally found mixed. Neither collection is very large and new collections of this species are needed to further define its characteristics.
Berriochloa schereri Elias var. marslandensis (Elias) Thomasson, comb. nov.
Stipidium schereri Elias mut. marslandense Elias, Geol. Soc. Amer. Sp. Paper 41:89. 1942.
Diagnosis--As given by Elias (1942).
Nomenclature--The type collection of this species is now at UNSM as UNSM-E28. I have designated UNSM-E28-L as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942).
Berriochloa ventricosa (Elias) Thomasson, comb. nov.
Stipidium ventricosum Elias, Geol. Soc. Amer. Sp. Paper 41:90. 1942.
Diagnosis--As given by Elias (1942).
Nomenclature--The type collection of this species is at UNSM as UNSM-E15. An investigation of the collection revealed that it was the mixed collection shown by Elias (1942, plate 7, Fig. 1). The original collection contained specimens of B. brevis and B. communis as well as the syntypes of B. ventricosa. I have excluded the specimens of the former two species from the types of B. ventricosa and I have also designated UNSM-E15-L as the lectotype.
Occurrences--No representatives of this species were collected during this study.
Discussion--See Elias (1942).
Genus Nassella Desvaux
Diagnosis--The genus Nassella is characterized by small, short, broad, often prolate, indurated anthoecia. The margins of the lemma completely overlap and enclose the greatly reduced palea (not seen in fossil Nassella). The short awn may be centrically or eccentrically attached and is either readily deciduous, persistent, or only tardily deciduous. The callus is short and usually blunt. Micromorphologically all species of Nassella are characterized by the pattern of epidermal cells on the lemma (Thomasson, 1978b). The basic "long" cells are reduced nearly to the appearance of a short cell but are distinguished by ripple walls. Specialized short cells may or may not be present. Most frequently these are represented as silica-suberin couples or silica cells. Hairs and hooks may also be present on the lemma surface.
Discussion--For a detailed discussion of the history of the genus Nassella see Thomasson (1976b).
| Key to the fossil species of Nassella | ||
|---|---|---|
| 1a. Surface of lemma smooth or with hooks restricted to the overlapping margin of the lemma. | ||
| 2a. Hooks completely absent from the lemma | N. pohlii | |
| 2b. Hooks restricted to margins of overlapping lemma | N. fryeleonardii | |
| 1b. Surface of lemma conspicuously covered with hooks. | ||
| 3a. Anthoecium pyriform, 1.3-1.6 mm long; lemma surface completely covered with hooks | N. amphora | |
| 3b. Anthoecium obovate, slightly laterally compressed, 1.6-2.1 mm long; lemma surface with hooks restricted to the medial 1/2-1/3 | N. reynoldsii | |
Nassella amphora Elias
(Pl. 3H; Pl. 10C)
Nassella amphora Elias, Geol. Soc. Amer. Sp. Paper 41:101 (1942).
Diagnosis--Little can be added to Elias's original description (Elias, 1942). However, scanning electron microscopy resolves more clearly hooks and faint vertical rows of short and long cells which confirm its correct assignment in this genus.
Nomenclature--The type specimens of this species are at UNSM as UNSM-E66. I designate UNSM-E66-L as the lectotype.
Occurrences--One specimen, possibly referable to this species, was collected in bed 9 at site 6.
Discussion--The pyriform nature of the anthoecia, their size, and the lemma surface completely covered with hooks distinguish this species from all other fossil Nassella species. It appears to be intermediate between the fossil Nassella species from Ellis County and the extant species of Nassella characterized by N. trichotoma.
Nassella fryeleonardii sp. nov.
(Pl. 4A, B; Pl. 10D)
Diagnosis--Anthoecia (palea absent) short, obovate, and slightly laterally compressed; length excluding the callus but including the neck 1.9-2.1 mm; width 1.0-1.2 mm; depth 1.2-1.3 mm; depth from the rear of the crown to the dorsal rib 0.5 mm. Greatest diameter and depth measured at about 3/4 above the base. Surface of anthoecia at low magnification (ca. 30x) smooth except for distinct hooks at the overlapping margins of the lemma. Short cells and greatly reduced long cells visible at high magnification. The neck is attenuated and with an apical scar to which an awn was probably attached. The callus is short (0.1 mm) and blunt.
Type locality and horizon--From the upper part of bed 1, SW SE SE sec. 29, T. 13 S., R. 19 W., Ellis Co., Kansas. This horizon is the "mortar" bed (impure tuff) below the Bemis Local Fauna.
Discussion--Anthoecia are very similar to those of N. pohlii except for the presence of hooks on the overlapping lemma margins. Only three specimens were found, but the fact that none of the hundreds of anthoecia of N. pohlii have distinctive hooks indicates that this is a new species.
This species is named in honor of John C. Frye and A. Byron Leonard. Their joint studies of the Ogallala Formation throughout the High Plains have contributed to a better understanding of its geology and paleontology.
Holotype collection--Holotype, Thomasson SEM No. BP12, 7 August 1974, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 29.56 in FHSM.
Nassella pohlii sp. nov.
(Pl. 4C; Pl. 10E)
Diagnosis--Anthoecia (palea not found preserved) short, laterally compressed, and obovate. Length of the anthoecia, excluding the short blunt callus, but including the neck, 1.4-2.1 mm; width 0.7-1.1 mm; depth 0.7-1.3 mm; depth from the rear of the crown to the dorsal rib 0.3-0.6 mm. Greatest diameter and depth measured at about 3/4 above the base. Surface of anthoecia minutely striate or smooth at low magnification (ca. 30x). Surface viewed by scanning electron microscopy characterized by vertical rows of long and short cells; long cells greatly shortened, possessing ripple walls; short cells crescent-shaped or oval, alternating with long cells and appearing possibly to be silica-suberin couples. Lemma margins overlap either right or left in usually equal frequencies. The distinct neck slender, not expanded into a crown at the apex, possessing a scar to which an awn was apparently attached; callus short (0.1 mm) and blunt.
Type locality and horizon--From approximately the middle of bed 2, at locality 4, site 39 NE SE SW sec. 4, T. 12 S., R. 20 W., Ellis Co., Kansas.
Additional occurrences--(1) A single specimen was collected in the uppermost 24 inches of bed 1 at locality 1, site 28. (2) Specimens were recovered from lower bed 4 at locality 1, and upper bed 2, site 39. (3) Numerous specimens in several clusters were collected throughout bed 2 at site 8. (4) At site 9a a few specimens were recovered in the upper 4 feet of bed 2 and immediately above the marl layer in bed 4, both at locality 1a.
Discussion--Nassella pohlii differs from all other fossil species of Nassella by the complete absence of any hooks on the surface of the lemma. Among fossil Nassella species, it approaches most closely in shape N. reynoldsii and N. fryeleonardii. A number of living Nassella species are equally smooth. Among these are N. chilensis, N. meyeniana, and N. juncea. Still others are smooth except for the presence of hairs on the callus or lemma surface. Included here are such species as N. humilis, N. johnstonii, N. pungens, and N. pubiflora. However, these living forms lack the distinct neck of the fossils, and possess a readily deciduous awn. Probably N. pohlii is most closely related to the group of Nassella species characterized by Nassella trichotoma. Additional similar species of Stipa that probably should be transferred to this group of Nassella include S. nidulans, S. pseudopampagrandensis, S. sanluisensis, and S. pampeana.
This fossil species is named in honor of Dr. Richard W. Pohl, Department of Botany, Iowa State University, who first introduced the author to the study of fossil grasses, and whose support and encouragement during the course of this study have been invaluable. His contributions to modern agrostology are well known.
Holotype collection--Holotype, Thomasson SEM No. BP54, 11 July 1975 as cluster B, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 39.51, 39.52, 39.53, and 39.54, FHSM, ISC, UNSM, and ROM.
Nassella reynoldsii sp. nov.
(Pl. 4D, E; Pl. 10F)
Diagnosis--Anthoecia (palea not found preserved) short, laterally compressed, obovate. Length 1.6-2.0 mm; width 0.8-0.9 mm; depth 0.8-1.1 mm; depth from the rear of the crown to the dorsal rib 0.3-0.5 mm; greatest diameter, width and depth are measured at approximately 3/4 the length above the base. Direction of overlap of lemma either right or left with equal frequency. Surface of the anthoecium with hooks restricted to a distinct band around the medial 1/2-2/3, the lower and upper 1/4-1/6 smooth at low magnifications (30x). At high magnification the lemma surface is marked by vertical rows of cells, alternating with hooks. Long cells were not observed because of the presence of the fossilized cuticle, but at least some specimens clearly indicated the presence of silica-suberin couples. Neck attenuated, usually terminating in a small scar, rarely with a portion of awn attached. Awn fragments, when preserved, as long as 0.1 mm and square in cross section with minute antrorse trichomes on the edges. No stems or leaves have been discovered.
Type locality and horizon--In the lower exposed portion of bed 1, approximately 1 meter below a volcanic ash bed, at site 21, SW SW SW sec. 29, T. 13 S., R. 19 W., Ellis Co., Kansas. This site slightly NE of a small dry pond.
Additional occurrences--(1) Abundant specimens throughout bed 2 at site 8. (2) Two specimens were recovered in bed 9 at site 6 and tentatively assigned here. (3) A single specimen, readily referable to this species, was collected in the upper 50 cm of bed 1 at site 28.
Discussion--Anthoecia of this species are similar in size and shape to fossil Nassella pohlii, but differ in the obvious presence of hooks. Nassella reynoldsii differs from N. amphora in three respects. The lemma in N. amphora is nearly always completely covered with hooks, only infrequently is the lower 1/6 smooth. In N. reynoldsii the hooks are always restricted to the medial 2/3. The shapes are also distinct between the species. The anthoecia of N. amphora are distinctly pyriform (width usually equals depth), whereas those of N. reynoldsii are almost always deeper than wide and as a result the anthoecia appear slightly laterally compressed. Finally, the lengths of the anthoecia are noticeably different. Those of N. amphora are 1.3-1.6 mm, with an average of about 1.5 mm, whereas those of N. reynoldsii are 1.6-2.1 mm long with an average of approximately 1.8 mm.
N. reynoldsii is undoubtedly closely related to both N. pohlii and N. fryeleonardii, and somewhat more distantly related to N. amphora. Among living species, N. reynoldsii seem most closely related to the Nassella trichotoma group.
This species is named in honor of Dr. Howard C. Reynolds, Division of Biological Sciences, Fort Hays Kansas State University whose enthusiasm in teaching taxonomy while the author was an undergraduate provided the stimulus for the author to pursue agrostology as a career.
Holotype collection--Holotype, Thomasson SEM No. BP104, 16 August 1973, in Sternberg Memorial Museum, FHSM; isotypes, Thomasson 21.4, 21.10, 21.11, and 21.12, in FHSM, ISC, UNSM, and ROM.
Tribe Paniceae
The Paniceae is one of the largest tribes of the Gramineae. According to Hsu (1965), who recently studied the classification of Panicum and its allies it consists of 81 genera and more than 1300 species Within the tribe, a number of characteristics unite the genera; those of the spikelet are of considerable significance. According to Hsu:
The tribe Paniceae is characterized by two-flowered spikelets with an indurate upper floret.... The upper floret shows various degrees of definite markings and rugosity in certain groups. . . . The surface view of the epidermis of the upper lemma (fertile lemma) would provide a good criterion for delimitation of the genera of the Paniceae.
It is fortunate that the fossils which have been found and referred to this tribe consist of the silicified anthoecia (lemma and palea).
Genus Panicum L.
Diagnosis--Only those characteristics of the florets that are normally preserved as fossils are considered here. Fertile lemma and palea indurate, either smooth or rugose; lemma margins involute and clasping the palea. Apex of the lemma obtuse, cuspidate, or with tuft of hairs. Anthoecia dorsally compressed, oval to obovate in outline. Lemma with an oblong to square germination lid at the base of the lemma. Scar of attachment oval to crescent-shaped, with three vascular trace scars.
Discussion--Hsu (1965) reported in his review of Panicum that more than 500 species comprising no less than 26 sections may be placed in this genus. Some authors (Blake, 1958; Rominger, 1962; Gould, 1974), however, have proposed transferring many of these to other genera and this may reduce the number of species in Panicum to less than 75 (Clark and Gould, 1975). For a detailed analysis and review of the genus Panicum and its allies the reader is referred to Hsu (1965).
Panicum elegans Elias
(Pl. 4F, G, H, I; Pl. 11A)
Panicum elegans Elias, Geol. Surv. Kans. Bull. 18:110. 1931. nom.nud.
Panicum elegans Elias, Univ. Kans. Sci. Bull. 20:342. 1932.
Panicum elegans Elias mut. nebraskense Elias, Geol. Soc. Amer. Sp. Paper 41:102. 1942.
Diagnosis--Anthoecia dorsally compressed, often appearing plano-convex due to the inflation of the lemma. A prominent palea is exposed between the margins of the lemma. Anthoecia 2.2-3.0 mm long; 1.2-2.0 mm wide; depth 1.0-1.7 mm. The surface of the lemma perfectly smooth at low magnification, long cells visible at high magnification, arranged in the "Ms-type" pattern (see Hsu, 1965); apex of lemma minutely cuspidate, the medial cusp as long as 0.1 mm; short cells absent. The palea surface appears smooth at low (ca. 30x) magnification, but at high magnification the distal and proximal portions are characterized by simple silicate papillae in regular rows. Two keels, which are usually conspicuous, run longitudinally the length of the palea. Upper 12 of palea frequently swollen outwards. Base of anthoecium with a crescent-shaped scar of attachment 0.60.7 mm long and 0.2-0.3 mm wide at its widest point. Three prominent circular vascular trace scars visible in many specimens, the medial scar larger than the two lateral scars. Well-developed germination lid present at the base of the lemma on the dorsal surface.
Nomenclature--Fragments which apparently are those of the holotype were located in collections at the University of Nebraska. In a vial containing the fragments was a label, "Panicum elegans Elias holotype figured in 1932, pl. 28, Figs. 2 and 4." This vial is now in UNSM as UNSM-E3, but the fragments are too small to be recognizable as the label indicates. It must be assumed that the holotype is destroyed. Another collection discovered (UNSM-E64) contains the label, "syntypes of Panicum elegans Elias, Wallace Co., Kansas, Ogallala abt. 50'from the base 1929." This seems to be the collection from which Elias drew the holotype; unfortunately, all the remaining specimens are also very fragmentary. I therefore designate as a neotype the specimen Thomasson No. 33.1 now at FHSM.
Occurrences--(1) In the upper 6 inches of bed 3, at locality 1, site 1. (2) In the lower 12 of bed 3 at locality 2, site 5. (3) In the lower 12 of bed 1 at locality 1, site 7. (4) In the upper 80 cm of bed 4 at locality 1a, site 9a. (5) In the approximate middle of bed 1, site 22. (6) 40 cm above contact of beds 1 and 2, at site 31. (7) In the upper 24 cm of bed 1 at locality 3a, site 30. Note: Graphic section 30 is a composite "stacked" section and does not necessarily correspond to the exact locality of this occurrence. (8) 2.2 meters below the contact of beds 1 and 2, in bed 2, at site 33. (9) In the lower middle part of bed 4 at locality 1, site 39.
Discussion--See Thomasson, 1978a.
Panicum chasei (Elias) Thomasson, comb. nov.
Setaria chasea Elias, Geol. Soc. Amer. Sp. Paper 41: 103. 1942.
Panicum eliasii Leonard, Univ. Kans. Sci. Bull. 38 (20):1398. 1958.
Diagnosis--As given in Elias (1942). At high magnification the lemma epidermis is characterized by long cells with well-developed simple lobes.
Nomenclature--The syntypes of this species are fragmentary specimens in collections at the University of Nebraska as UNSM-E63(1). These were specimens figured by Elias (1942, Pl. 16, Fig. 10).
Discussion--The fragmentary nature of the type collections shows macromorphologically only the presence of cusps at the apex of the lemma, and the rugose nature of both the lemma and palea. I have examined by light microscopy a fragment of the lemma which shows the lemmatal pattern to be of the "Ms-type." In the absence of any other characteristics, this evidence is used to refer this species to the genus Panicum. Little can be added to the description given for P. eliasii by Leonard (1958) and his description should be referred to for further information. His "vertical diameter" is equivalent in my references to lemma depth. Leonard listed a maximum length of slightly over 3.0 mm, but I found one to be 3.5 mm. Elias gave no quantitative measurements of this taxon, probably because of the fragmentary nature of the types. The Leonard collections studied are KANU 9975 (ABL, A. B. Leonard, 752), the holotype; and, KANU 9976 (ABL, A. B. Leonard, 752), paratypes.
Occurrences--No specimens referable to this species were discovered during the course of this study.
Discussion--This specimen is transferred to Panicum on the basis of the micromorphological characteristics of the lemma discussed in the description.
Genus Graminophyllum Conwentz
Diagnosis--An organ genus of the Gramineae including leaves and epidermal fragments. Long cells and short cells present alternating in rows; guard cells around each stoma enlarged at either end (dumbbell-shaped), narrowed in the middle; two subsidiary cells present, parallel to the guard cells and stomata.
Discussion--See Thomasson, 1976b for a historical sketch of the genus Graminophyllum.
Graminophyllum kansasense sp. nov.
(Pl. 11B)
Diagnosis--Fragments of leaf epidermis; long cells with sinuous walls in rows, alternating with short cells; short cells present as silica-suberin couples, silica cell circular, suberin cell crescent-shaped. Stomata in rows, alternating with long cells; guard cells of each stoma thickened at each end and narrowed in the middle (dumbbell-shaped), subsidiary cells adjacent to and parallel with the guard cells, low dome-shaped.
Type locality and horizon--From the upper 1/2 of bed 2, at site 2, SW NE SW sec. 4, T. 12 S., R. 20 W., Ellis County, Kansas.
Discussion--These fragments were isolated from the mass occurrence of Berriochloa tuberculata illustrated in Fig. 5A. There is no doubt that these are fragments of a grass leaf as the characteristics of the stomata and short cells are those ascribed to members of the Gramineae (Metcalfe, 1960; Litke, 19,68). On the basis of the short cells paired in silica-suberin couples, this is probably a member of the grass subfamily Pooideae.
Holotype collection--Thomasson No. 2.1, 25 May 1973, Sternberg Memorial Museum (FHSM).
Family Cyperaceae
? Tribe Scirpeae Nees
Genus Eleofimbris gen. nov.
Diagnosis--An organ genus of the Cyperaceae represented by the hulls of achenes. Shape of the achene usually trigonous with one face broader than the other two, or plano-convex. Achenes minutely tessellate with vertically oriented rectangular depressions, narrowed towards the summit, terminating in an expanded style base scar above a slightly constricted neck. The achenes also narrow towards the base, ending in an unexpanded, generally trigonous scar.
Type species--Eleofimbris svensonii sp. nov.
Discussion--The name Eleofimbris is proposed for the achenes of a member of the Cyperaceae that have been found twice in the late Miocene-early Pliocene rocks exposed in Ellis County, Kansas. Since the fossils can be assigned without doubt to the Cyperaceae, this genus is an organ genus. The fossils are most similar to Scirpus, Eleocharis, and Fimbristylis. In living forms, the genus Scirpus is separated from Eleocharis and Fimbristylis by the absence of a jointed pistil, i.e., the ovary and the style are continuous with no line of articulation or constriction separating the achene from the style (Koyama, 1961). Eleocharis and Fimbristylis have jointed pistils and a distinct line of articulation or constriction is present. In addition, both genera usually have enlarged style bases (Svenson, 1957). The genus Eleocharis differs from Fimbristylis in a number of morphological characters (Svenson, 1957; Koyama, 1961; Kral, 1971) and cytology (Koyama, 1961). Eleofimbris fossils possess a triangular scar at the apex of the achene which is expanded above a slightly constricted neck. I interpret this as the point at which the style base was articulated with the ovary; hence, the fossils had a jointed pistil and Scirpus can be eliminated from consideration in the assignment of the fossils to a genus. Unfortunately, none of the characters which effectively separate Eleocharis and Fimbristylis are preserved on the fossils, and they cannot be assigned with certainty to either of these living genera. The organ genus Cyperacites established by Schimper (1870) included rhizomes, stems, leaves, flowers, and fruits of the Cyperaceae. The single fruit described closely resembled that of a species of Cyperus. Pax (1906) described a fossil achene of a member of the Cyperaceae which resembled the living genus Kobresia and proposed for it the organ genus Cyperocarpus. Although the Kansas fossils might be assigned to Cyperacites, which has priority over Cyperocarpus, I have not done so for two reasons. First, the Kansas fossils are clearly related to a much different group within the Cyperaceae than the fruit originally described under Cyperacites. Applying the name Cyperacites to them would imply a much closer relationship than is indicated by the fossils. Second, the stratigraphic relationship of the Kansas fossils with those of Europe is unknown. For the above reasons I have felt it necessary to adopt the new genus, Eleofimbris.
Eleofimbris svensonii sp. nov.
(Pl. 5A, B; Pl. 11C)
Diagnosis--Achenes inequilaterally trigonous with one face broader than the other two, or plano-convex with one face broader and flattened. The length of the achenes, including style base, 0.8-1.1 mm; width 0.5-0.8 mm across the broadest face; surface smooth to minutely reticulate under low magnification (ca. 30x). When viewed under the scanning electron microscope, a tessellate pattern of epidermal cells with undulating cell walls can be observed. Style base scar triangular and approximately 0.1-0.2 mm wide. The achenes narrow towards the base, but the basal width is always greater than that of the style base scar, varying between 0.2-0.25 mm. No seeds or remnants of seeds have been recovered in any of the achenes.
Type locality and horizon--From approximately the middle of bed 3 at locality 2, site 5, NW NW sec. 17, T. 12 S., R. 18 W., Ellis Co., Kansas.
Other occurrences--From the lower part of bed 1 at locality 1 (silage pit), site 30, CSL SE SW sec. 9, T. 12 S., R. 18 W., Ellis Co., Kansas.
Discussion--Fossilized achenes of representatives of the Cyperaceae are not uncommon in Tertiary strata of the eastern hemisphere. Species of the genera Carex, Cladium, Dulichium, and Scirpus have been reported from Russia ( Dorofeev, 1958, 1969) and Eleocharis (Heleocharis) is known from numerous localities in Eurasia (Lancucka-Srodoniowa, 1966). Reports of cyperaceous achenes from the Tertiary strata of the western hemisphere are much less common. Achenes and a spike of Carex and Scirpus have been reported from Canada (Penhallow, 1908). Elias (1941) reported a Carex-like member of the Cyperaceae from Nebraska, but be gave no description, and I was unable to locate any specimens in his collections.
Only the outer walls of the achenes are preserved, but scanning electron microscopy shows that features of these walls are preserved in considerable detail. This is especially significant since Schuyler (1971) has shown, in a scanning electron microscopy study of the achene epidermis in species of Scirpus and related genera, that the epidermal cells provide evidence for determining taxonomic relationships. Although detailed comparisons were not attempted with Eleofimbris, such a study might reveal more closely the relationships of the fossils to modern species.
As previously noted, the fossils cannot be assigned to living genus. However, this does not preclude one from comparing the fossils with modern species which most resemble them. I have done this both through the examination of herbarium specimens and by comparisons with the illustrations and descriptions found in studies of Eleocharis (Svenson, 1929, 1932, 1934, 1937, 1939), Fimbristylis (Svenson, 1957; Kral, 1971), and the Cyperaceae in general (Clarke, 1909). The fossils are most similar to living species of Eleocharis of the Series Palustriformes. Closest among these to the fossils are Eleocharis palmeri and E. parishii, which are chiefly North American, and E. montevidense, which is principally South American.
The species is named in honor of H. K. Svenson, whose monographic studies in the genus Eleocharis have contributed greatly to a better understanding of this genus.
Holotype collection--Holotype, Thomasson SEM No. BP165, 31 May 1973 in Sternberg Memorial Museum (FHSM), Hays, Kansas; isotypes, Thomasson, No. 5.8, 5.9, 5.10, and 5.11, in FHSM, ISC, UNSM, and ROM.
Class Magnoliopsida
Family Ulmaceae
Genus Celtis L.
Celtis willistonii (Cockerell) Berry
Tithymalus willistonii Cockerell, Torreya 9:119. 1909.
Celtis besseyi Barbour, Univ. Nebr. State Mus. Bull. 8. 1:89. 1925.
Celtis willistonii (Cockerell) Berry, Amer. Mus. Novitates No. 298:2. 1928.
Diagnosis--As given by Segal (1966b).
Nomenclature--Berry (1928a) indicates that the type specimens of this species are in the Museum of the University of Colorado. I have not confirmed this.
Occurrences--(1) In the lower half of bed 4 at site 1. (2) In the prominent "mortar" bed at site 3. (3) At site 5 in bed 2 at localities 1 and 2. (4) In the lower 1 m of bed 9 at locality 3, site 6. (5) In beds 1 and 5 at site 7 (lower occurrence not shown on Pl. 1, graphic section 7). (6) About the middle of bed 2 at site 8. (7) In the upper half of bed 2 at site 9. (8) In the lower 20 cm of bed 2 at locality 1 site 10. (9) About the middle of bed 1 at site 11. (10) In the lower 1 m of bed 4, at site 13. (11) In the upper 1 m of bed 1 at site 21. (12) In the uppermost exposed mortar beds at site 30. (13) In beds 1 and 8 at site 32. (14) In the lower 50 cm of bed 4 at site 39. (15) Immediately below the tortoise in Fig. 5D in bed 2 at site 41.
Discussion--See Segal (1966) and Thomasson (1977). Reports of the endocarps of Celtis species from the Tertiary of North America are common (Hatcher, 1902; Wanless, 1923; Chaney, 1925; Elias, 1942; Clark, Beerbower, and Kietzke, 1967, among others). They have also been reported from the Tertiary of Europe (Knobloch, 1969).
Family Boraginaceae
Genus Cryptantha Lehm.
Discussion--See Segal (1964).
Cryptantha coroniformis (Elias) Segal
(Pl. 5C, D, E, F; Pl. 11E)
Krynitzkia (Cryptantha) coroniformis Elias, Univ. Kans. Sci. Bull. 20:356. 1932.
Cryptantha coroniformis (Elias) Segal, Trans. Kans. Acad. Sci. 67:203. 1966.
Diagnosis--As given by Elias (1932) and Segal (1966a) with the following additions. Nutlet length 1.9-2.4 mm, depth (height) 1.0-1.4 mm, width 1.2-1.7 mm. At high magnifications epidermis characterized by irregular cells possessing well-developed lobes; lobes simple or compound, tips expanded, obtuse.
Occurrences--(1) In bed 1 at site 1. (2) In upper bed 2 at site 47. Occurrences (1) and (2) both Ellis County, Kansas. (3) Approximately 3 meters below the top of the exposed beds at site 47, at the religious monument on the SE edge of Wray, Yuma County, Colorado.
Nomenclature--The types of this species are in UNSM as UNSM-E7.
Discussion--See Elias (1932, 1942) and Segal 1966a).
Cryptantha auriculata (Elias) Segal
(Pl. 5G, H)
Krynitzkia auriculata Elias, Univ. Kans. Sci. Bull. 20: 358. 1932.
Cryptantha auriculata (Elias) Segal, Trans. Kans. Acad. Sci. 67:203. 1966.
Diagnosis--As given in Elias (1932) and Segal 1966a).
Nomenclature--The types, or at least part of them, are at KANU as "Cryptantha auriculata, fragments of types, Maxim K. Elias 181." The handwriting on the label in the vial appears to be that of Dr. Elias and these are undoubtedly part of the types of C. auriculata.
Occurrence--(1) In the upper 1 meter of bed 6 at site 42.
Discussion--See Elias (1932) and Segal (1966a). This species was found only once high in the P. johnstonii-N. pohlii Zone. Nearly all nutlets are halves, perhaps indicating rodent damage.
Cryptantha parvicarpa sp. nov.
(Pl. 6A, B)
Diagnosis--Nutlets triangular-ovate, acute at the apex; ventral scar of attachment triangular, extending the length of the nutlet; base rounded; nutlet length 1.6-1.9 mm, depth (height) 1.0-1.2 mm; width 1.11.5 mm; surface of nutlet tuberculate.
Type locality and horizon--In bed 2 at site 9c, in the NW NW sec. 3, T. 12 S., R. 20 W., Ellis Co., Kansas.
Discussion--This species of Cryptantha differs from all other species in the small size of the nutlet. The stratum from which the nutlets come lies below the Hamburg local fauna (Zehr, 1974).
Holotype collection--Holotype, Thomasson SEM No. BP179, 31 July 1974, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 9.14, 9.15, in FHSM, ISC.
Eliasiana gen. nov.
Diagnosis--An organ genus represented only by nutlets; nutlets asymmetrically inflated, ovoid in shape; two prominent keels present, a main ventral keel and a less prominent lateral keel; lateral keel turns either right or left resulting in left or right nutlets; surface of nutlet reticulate and minutely aculeate. Scar of attachment funnel-shaped, narrowing into a small deep hole in the center; rim of scar flared.
Discussion--This genus appears to combine the characteristics of both Biorbia and Prolithospermum. It differs from Biorbia in the possession of a funnel-shaped scar and from Prolithospermum in that the base of the alar projection is not obviously gibbous. It is quite similar to some species of the living Anchusa. The genus is named in honor of Maxim K. Elias, the father of modern paleoagrostology, who pioneered the studies of fossil anthoecia, nutlets, achenes, and endocarps from the High Plains Tertiary deposits.
Type species--Eliasiana aculeata sp. nov.
Eliasiana aculeata sp. nov.
(Pl. 6C, D, E, F)
Diagnosis--Nutlets ovoid, asymmetrically inflated, length 1.9-2.5 mm, width 1.7-2.0 mm, depth 1.5-1.9 mm; base swollen, tapers to an obtuse apex; alar projection (ventral keel) prominent, arises at a funnel-shaped scar, extends over the apex and then turns right or left forming a prominent lateral ridge which connects with the funnel-shaped scar; surface of nutlet rugose with reticulate ridges, at high magnifications aculeate; scar of attachment funnel-shaped, neck of scar not constricted nor distinctly gibbous at the base.
Type locality and occurrence--In bed 2 at site 47, SW SE sec. 6, T. 12 S., R. 20 W., Ellis Co., Kansas.
Discussion--This species is similar to Biorbia fossilia, Prolithospermum johnstonii, and P. grossii. However, from Biorbia fossilia it differs in the distinct funnel-shaped scar; from Prolithospermum johnstonii and P. grossii it differs in the lack of a distinct gibbous protuberance near the scar of attachment at the base of the alar projection (ventral keel). It appears to combine the characteristics of species of both Biorbia and Prolithospermum. This taxon was not found in association with either Biorbia fossilia or Prolithospermum spp. and may represent an evolutionary intermediate between these and the nearest living taxon. In this regard Eliasiana aculeata sp. nov. is very similar to and probably a progenitor of species closely related to Anchusa sempervirens. This living species has a funnel-shaped scar with a flared rim, and surface features which relate it immediately with the fossil Eliasiana aculeata sp. nov. E. aculeata possibly represents the ancestor of taxa that migrated to Europe from North America but became extinct in North America.
Holotype collection--Holotype, Thomasson SEM No. BP82, 22 December 1975, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 47.1, 47.2, 47.3, and 47.4 in FHSM, ISC, UNSM, and ROM.
Genus Prolithospermum Elias
Prolithospermum Elias, Geol. Soc. Amer. Sp. Paper 41:105. 1942.
Diagnosis--As given in Elias (1942) with the following additions on micromorphology. Epidermal cells of nutlets with sinuous walls; cells irregularly shaped and arranged.
Type species--Prolithospermum johnstonii Elias.
Discussion--See discussion of Eliasiana.
| Key to the Species of Prolithospermum | |
|---|---|
| 1. Nutlet surface smooth; lateral ridge absent | P. johnstonii |
| 1. Nutlet surface reticulate; prominent lateral ridge present | P. grossii |
Prolithospermum johnstonii Elias
(Pl. 6G, H; Pl. 7A; Pl. 11D)
Prolithospermum johnstonii Elias, Geol. Soc. Amer. Sp. Paper 41:105. 1942.
Diagnosis--As given in Elias with the addition of new dimensions and features of micromorphology. Nutlets 2.5-3.0 mm long, depth (height) of nutlets 1.9-2.5 mm, width 1.8-2.4 mm. At low magnification, nutlets smooth; at high magnification, with a pattern of irregularly lobed and arranged epidermal cells. Lobes of epidermal cells simple, somewhat angular.
Nomenclature--Since the holotype and all its duplicates are either destroyed or missing, I am designating a neotype. The neotype is Thomasson No. 6.18, 12 August 1975, now at FHSM.
Occurrence--(1) In bed 1, site 2 as clusters J and R, 5 July 1975 and unlettered collection 9 July 1975. (2) A number of collections in the lower 100 cm of bed 9, locality 3 at site 6. Neotype comes from this collection. (3) At site 9a in bed 2, 1.2 m below the contact of beds 1 and 2; in lower bed 2 (= bed 5 at site 9a) of site 9b about 35 cm above the contacts of beds 1 and 2. (4) At site 42 in upper bed 6 as clusters E, F, and G, 2 August 1975.
Discussion--Elias (1942, p. 122) believed this species to be a transitional form between Biorbia fossilia and Lithospermum linearifolium. I have found occurrences of Prolithospermum johnstonii well below those of Biorbia fossilia and I do not accept his interpretation. It appears to me that if species of the genus Prolithospermum can be related to living species, then they are best related to forms such as the European Anchusa sempervirens L. possibly through or along with Eliasiana aculeata sp. nov. See also discussion of Prolithospermum grossii, sp. nov. and Eliasiana aculeata, gen. nov., sp. nov.
Neotype collection--Neotype, Thomasson No. 6. 18, 12 August 1975, at FHSM.
Prolithospermum grossii, sp. nov.
(Pl. 7B)
Diagnosis--Nutlets asymmetric, either right or left, shape ovate; alar projection = ventral keel) distinct, gibbous at the base at the site of the attachment of the nutlet to the gynobase; lateral ridge also present; scar of attachment funnel-shaped; length of nutlets 2.8-3.1 mm, depth (height) 2.0-2.4 mm, width 2.0-2.3 mm; surface of nutlet reticulate at low magnifications; at high magnification with an epidermal pattern of irregular cells with complex lobes (this may be a pseudoepidermal pattern due to solution or some other unknown factor).
Type locality and horizon--In bed 1 at site 2, SW NE SW sec. 4, T. 12 S., R. 20 W., Ellis Co., Kansas, as cluster R, 5 July 1975.
Discussion--This species is most similar to P. johnstonii except for the presence of a distinct lateral ridge in addition to the ventral keel and the reticulate nature of the nutlet surface. In these characteristics P. grossii also closely resembles B. fossilia but the gibbous nature of the base of the ventral keel and the tubular nature of the scar of attachment clearly refers it to Prolithospermum.
The species is named in honor of Mr. Harold Dean Gross, Hays, Kansas who accompanied and assisted the author during many collecting trips, including the one on which this taxon was discovered.
Holotype collection--Holotype, Thomasson SEM No. BP178, 5 July 1976, in Sternberg Memorial Museum (FHSM), Hays, Kansas; isotype, Thomasson 2.4 and 2.5 in FHSM and ISC.
Genus Biorbia Elias
Biorbia Elias, Univ. Kans. Sci. Bull. 20:350. 1932.
Diagnosis--As given by Elias (1932) and emended by Segal (1966c) with the addition of new information on micromorphology. Surface of nutlets at high magnification with irregularly lobed and arranged epidermal cells; lobes simple.
Type species--Lithospermum fossilium var. rugosum Berry.
Discussion--See Elias (1932), Segal (1966c), and Thomasson (1977).
Biorbia fossilia (Berry) Cockerell
(Pl. 7C, D, E; Pl. 11F)
Lithospermum fossilium var. rugosum Berry, U.S. Nat. Mus. Proc. 73:1. 1928.
Celtis microendocarpica Brooks, Carnegie Mus. Ann. 18:299. 1928.
Biorbia rugosa (Berry) Elias, 1931, State Geol. Surv. Kans., Bull. 32:150. 1931, nom. nud.
Biorbia rugosa (Berry) Elias, Univ. Kans. Sci. Bull. 20:350. 1932, comb. nov.
Biorbia fossilia (Berry) Cockerell, Torreya 33:15. 1933, comb. nov.
Biorbia fossilia (Berry) Elias, Geol. Soc. Amer., Spec. Paper 41:106. 1942, comb. nov.
Biorbia fossilia (Berry) Cockerell var. parvula Segal, Univ. Kans. Sci. Bull. 46:499. 1966.
Biorbia levis Segal, Univ. Kans. Sci. Bull. 46:499. 1966.
Biorbia levis Segal var. minima Segal, Univ. Kans. Sci. Bull. 46:500. 1966.
Biorbia microendocarpica (Brooks) Leonard, Am. Mid. Natl. 22:491. 1977.
Diagnosis--As given by Elias (1932) with the following modifications and additions. Dimensions of nutlets extremely variable; length 1.9-3.4 mm, depth 1.3-2.5 mm, width 1.4-2.5 mm; surface of nutlet at low magnifications reticulate to smooth, at high magnifications with epidermal cells with sinuous walls. Lobes of cell walls simple.
Nomenclature--The types of Biorbia fossilia are a collection of J. B. Hatcher from Phillips Co., Kansas now at USNM.
Occurrences--(1) Site 1 at locality 1, throughout bed 4. (2) Site 2, throughout bed 2 as clusters G and CC. (3) Site 4 in bed 1 at locality 2, bed 1 at locality 4 and in bed 3 at locality 5. (4) Bed 9, localities 1 and 3 at site 6. (5) Site 7, lower 30 cm of bed 1, as cluster B. (6) Site 9, locality 1, in the middle of bed 2 and in the upper meter of bed 4. (7) Site 10, bed 2 at locality 1 and bed 1 at locality 2. (8) Site 11, the lower 10 cm of bed 3 as cluster B. (9) Site 14 in the small exposure present. (10) Site 16, bed 1. (11) Site 21, in the lower meter of bed 3 at locality 1a as cluster A. (12) Site 22, in the middle of bed 1 at locality 1. (13) Site 24, in the small exposure present. (14) Site 31, in the upper portion of bed 2 as cluster A. (15) Site 32, in beds 1 and 8. (16) Site 33, in the upper 1 meter of bed 1. (17) Site 38, in the middle of bed 3. (18) Site 30, at all localities, throughout beds 1, 2, and 6. (19) Site 39, in the upper 20 cm of bed 2 at locality 3 and throughout the lower 2 m of bed 4 at locality 1. (20) Site 40 (Smith County, Kansas) in the middle of bed 1 at locality 1. (21) Site 41, in the lower portion of bed 3. (22) Site 44 (Garden County, Nebraska), in the exposed beds at locality 1. (23) Site 46 (Gove County, Kansas), bed 8 at locality 1, bed 6 at locality 3, bed 7 at locality 4 and beds 7 and 8 at locality 5.
Discussion--Biorbia fossilia is an extremely variable taxon. I have not found it possible, with the possible exception of Biorbia papillosa Leonard, to assign collections to one or another "species" of previous authors with any degree of confidence. See Thomasson (1977) for a detailed discussion of this problem.
Prolappula gen. nov.
Diagnosis--An organ genus of the Boraginaceae represented by nutlets. Nutlets oblong-triangular when viewed from the ventral side and triangular in cross section. Nutlets with three faces, the dorsal face (disk) broadest with two marginal ribs to which barbed hairs are attached. The two lateral faces (sides) taper to a ventral keel. Ventral keel running the entire length of the nutlet, apparently representing the scar of former attachment of the nutlet to the gynophore. Two small lateral keels diverge at right angles to the ventral keel at the base of the nutlet. Base of nutlet slightly rounded to almost truncate. Surface of nutlet (sides, disk, and base) minutely tuberculate.
Type species--Prolappula verrucosa sp. nov.
Discussion--The name Prolappula is proposed for the nutlets of a member of the Boraginaceae which have been found once in late Miocene-early Pliocene rocks exposed in Ellis County, Kansas. Living genera which have nutlets similar to this genus are Lappula, Hackelia, Trianschaniella, and Eritrichium. However, the ventral keel, representing the attachment of the nutlet to the gynobase of the parent plant, is present the entire length of the nutlet only in Lappula. In Hackelia, Trianschaniella, and Eritrichium, the scar is medial. The actual scar on the nutlet through which the vascular trace passes is termed the cicatrice (Popov, 1953). Species of Lappula have a basal cicatrice, whereas species of Hackelia, Trianchaniella, and Eritrichium have a medial cicatrice. The cicatrice on Prolappula verrucosa is basal. Another important distinction among these genera is the shape of the gynophore. However, neither the gynophore nor any other reproductive or vegetative parts are preserved on the fossils. Therefore, even though these fossils are undoubtedly closely related to Lappula, I have chosen to place them in the new genus Prolappula.
Prolappula verrucosa sp. nov.
(Pl. 7F, G, H)
Diagnosis--Nutlets oblong-triangular in ventral view; triangular in cross section. Nutlets slightly distorted, but dimensions as follows: length 1.5-1.8 mm, depth 1.0-1.1 mm, width 0.8 mm. Nutlets three-faced; one face (disk) broader than the other two. Simple or compound barbed hairs 0.2-0.3 mm long present on the slightly thickened margins of the disk. Lateral faces taper to a ventral keel which runs the entire length of the nutlet; at the base lateral keels arise at right angles to the ventral keel. Base of nutlet slightly rounded to almost truncate. Surface of nutlets tuberculate on all faces at low magnifications (ca 30x); high magnification reveals abundant bulbous tubercles from which secondary protuberances arise. Scattered, broad-based, elongated prickles with secondary protuberances also present (Pl. 7H). No vegetative material has been recovered.
Type locality and horizon--From the lower 20 inches of bed 4, of site 9a, east wall of northernmost roadcut along federal-aid-secondary (FAS) highway 231, WL SW NW NW sec. 3, T. 12 S., R. 20 W., Ellis Co., Kansas (Fig. 6).
Discussion--There are no previous reports of fossils similar to these from any geologic strata. Among living forms the fossils are most similar on the basis of gross morphology to species of Lappula, particularly L. occidentalis (S. Wats.) Greene and L. redowskii (Hornem.) Greene. However, the fossils differ from these in the base of the nutlet being almost truncate, and in the possession of two short lateral keels at the base of the ventral keel. In microscopic character, Prolappula verrucosa nutlets are very similar to L. redowskii and L. texana (Schule) Britt. Prolappula verrucosa differs from the latter two species in the possession of tubercles which have many secondary cylindrical protuberances as opposed to few. In addition, the protuberances of the elongated prickles in P. verrucosa are cylindrical, whereas those of Lappula redowskii and Lappula texana are distinctly pustule-shaped.
Holotype collection--Holotype, Thomasson SEM No. BP18, 23 December 1974, in Sternberg Memorial Museum (FHSM); isotypes, Thomasson No. 9.10, 9.11, 9.18, and 9.19 in FHSM, ISC, UNSM, and ROM.
"Daemonelix"
"Daemonelix" is an obsolete generic name that was applied by Barbour (1892, 1897) to a varied assortment of fossil burrows, caliche stringers, and plant roots. The structures originally described were eventually proven to be the large corkscrew burrows of large fossorial beavers, Paleocastor spp., and in fact skeletons have been found in some burrows. Schultz (1942) has given an excellent review of the history of the "Daemonelix" problem in which are contained copious references.
Among the remains identified as "Daemonelix" by Barbour are some which are undoubtedly of a vegetable origin. Barbour (1897) described and illustrated two such groups of aggregations of "Daemonelix" which he termed cakes, fingers, and balls. I have found identical structures at numerous localities in Ellis County (Fig. 5C). Usually they occur in well-sorted silts. Altogether these structures appear much like algal mats or balls which I have seen quite frequently along modern streams, where they are found in flooded depressions on floodplains. However, close examination of the remains reveals the presence of secondary branches on many of the fibers. These cakes, balls, and fingers of "Daemonelix" are undoubtedly masses of roots and in fact many individual roots appear very similar to seminal roots of many germinating grasses. The origin of the masses themselves is not known. Possibly they are fecal ejections of some sort or burrow "fillings." Their restricted occurrence in Ellis County deposits appears to indicate a specific depositional environment, possibly on the margins of streams or on the floodplains.
Prev Page--Paleoecology and Paleoclimatology || Next Page--References
Kansas Geological Survey, Geology
Placed on web May 1, 2009; originally published September 1979.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/218/06_syst.html