![]()
Prev Page--Deposition || Next Page--Gas Production, Summary
Paleoecology of Macroinvertebrate Fossils
Preservation
Most Smoky Hill macro invertebrate fossils are preserved as (1) calcareous skeletal material, (2) molds, (3) borings, (4) combinations of calcareous and organic matter, and (5) chitin.
In the first category are:
- Bivalves
- Inoceramidae (abundant)
- Ostreidae (abundant)
- Radiolitidae (common)
- Bakevelliidae (sparse)
- Pectinidae (sparse)
- Cirripeds
- Scalpellidae (common)
- Stramentidae (common)
- Cephalopods
- Belemnitellidae (sparse)
- Collignoniceratidae (aptychi, rare)
- Crinoids
- Uintacrinidae (sparse)
- Annelids
- Serpulidae (rare)
In the second category are:
- Bivalves
- Lucinidae (rare)
- Inoceramidae (common)
- Cephalopods
- Scaphitidae (common)
- Baculitidae (common)
- Collignoniceratidae? (rare)
- Nautilidae (rare)
The third category includes:
- Sponges
- Clionidae? (sparse)
- Cirripeds
- Acrothoracica (common)
The fourth category includes:
- Cephalopods
- Teuthida (sparse)
The final category includes:
- Crustacea
- Malacostraca (rare)
Among the above-listed fossil groups, inoceramids and oysters are by far the most abundant forms, and both are nearly ubiquitous in measured chalk units of the composite section. Inoceramids are preserved mostly as calcitic valves in which only the prismatic layer remains. In these specimens the inner, nacreous layer is invariably absent, owing to early diagenetic dissolution. Large, platter-shaped inoceramids such as Inoceramus (Platyceramus) platinus are generally preserved intact, with paired valves pressed tightly together and usually with no sediment between the valves. Many valves encrusted by the oyster Pseudoperna congesta are so much compacted that the valve has been deformed around the encrusters and oyster outlines are clearly visible on the inoceramid valve interior. Some of these large inoceramids have suffered breakage during compaction, and valve fragments may be imbricated. Locally, in situ disintegration of these inoceramids has produced lenses of calcirudite, but presence in such lenses of chalk matrix demonstrates that such lenses were not produced by mechanical transport.
Large bowl-shaped inoceramids assigned to Inoceramus (Volviceramus) grandis are almost everywhere deformed and broken by compaction. Many collected specimens remain articulated (Pl. 3, 1) but most of the lower valve occurs as small fragments.
Small, thin-shelled inoceramids such as I. cycloides and I. balticus are invariably much compacted. Collected specimens are mostly molds, although in most parts of the thin prismatic shell layer are preserved.
Smoky Hill oysters, nearly all of which are referred here to Pseudoperna congesta, are represented almost entirely by well-preserved calcitic valves in which original lamellar structure remains intact. Most of these oysters are cemented firmly to exterior surfaces of inoceramid bivalves, but oysters occur also on rudists and on molds of scaphites and baculites. In some beds oysters appear to be isolated within chalk, but upon close inspection of left valves, even these are seen to have attachment scars that testify to the epizoic character of all such oysters. The oysters occur as spat, juveniles, and adults, the latter commonly with thick shells and high side walls where crowded. Valves are commonly articulated, and compaction has pressed the right valves tightly against left ones.
The rudist Durania has a calcitic skeleton in which structural detail is well preserved. Original void spaces within the shell wall have been filled with sparry calcite. Most specimens have suffered compaction, and some were broken apart before burial as attested by attachment of epizoic oysters even to broken surfaces.
Other Smoky Hill bivalves are of more restricted stratigraphic occurrence than the above-described groups. Phelopteria, recorded from just beneath Marker Unit 7 and from the zone of Clioscaphites choteauensis, has calcitic valves that were flattened by compaction. Peetinids occur as very thin, flattened, calcitic valves.
Smoky Hill lepadomorph cirripeds occur as articulated specimens or isolated valves that are invariably calcitic. Most of the specimens are minute and were recorded only by microscopic scanning of inoceramids, oysters, and ammonite molds.
The few known Smoky Hill belemnitellids are all calcitic and are preserved in the round. The recorded specimens of aptychi (Fischer and Fay, 1953) are apparently all calcitic. The single, possible sepioid reported by Miller (1968, p. 56) is calcitic, but is of vertebrate origin (J. D. Stewart, written communication, 1982).
All specimens of Uintacrinus socialis are calcitic and most, if not all, have suffered from effects of compaction and dissolution (Neugebauer, 1978a; Hattin, 1981). The. specimens occur mostly in thin lenses of hard, brittle limestone in which some coccolith-rich matrix occurs.
Data on Smoky Hill serpulids are suspect. Of the three species occurring presumably in the Smoky Hill (Miller, 1968), only Serpula intrica White has locality data that strongly suggest a Smoky Hill occurrence but the record is based on a single specimen, which has been lost. The other two species, S. tenuicarinata and S. semicoalita, are abundant in the Fairport Member of the Carlile Shale. Despite many months of field work and extensive microscopic examination of substrates (inoceramids, oysters) on which serpulids are found commonly, I have not recorded specimens in the Smoky Hill Member. In other chalk units of Kansas serpulids are invariably calcitic and are preserved in the round.
Fossils preserved as molds are more difficult to detect in the field, and with few exceptions are discovered by extensive splitting of the chalk. All molds, whether of cephalopods or bivalves, have been flattened by compaction, although most specimens retain at least a small thickness.
Sparse circular pits in inoceramid bivalves are attributed tentatively to the boring sponge, Cliona. Extensive systems of excavated galleries within valves are very rare. In contrast, acrothoracican borings are common in the Smoky Hill, and are abundant in the shelly zones. Inoceramids were the usual substrates for these cirripeds, but such borings occur also on some Durania specimens. Most are assignable to the genus Rogerella.
Teuthid coleoids are sparse in most Smoky Hill rocks. Well-preserved specimens comprise a broad, flattened gladius and long, cylindrical guard. The skeleton of these cephalopods is more or less calcified, but retains much of the original conchiolin of which Recent teuthids are largely composed.
Except for cirripeds the Crustacea are poorly represented in Smoky Hill rocks. Malacostracans, which preserve the original chitinous exoskeleton, are known from very rare, imperfectly preserved specimens.
Inoceramids
Smoky Hill strata contain few species of benthonic macroinvertebrates, and those represented by body fossils were epifaunal suspension feeders. Inoceramids and ostreids were the principal macroinvertebrates that inhabited the sea floor during deposition of the member. Principal growth forms of inoceramids include large, highly inequivalved, bowl-shaped, thick-shelled forms, represented by Inoceramus (Volviceramus) grandis; giantsized, equivalved, slightly biconvex forms, represented by Inoceramus (Platyceramus) platinus and Inoceramus (Cladoceramus) undulatoplicatus; and small- to medium-sized, thin-shelled, elongate, biconvex forms with prominent growth ridges, represented by Inoceramus stantoni, I. balticus, and I. sp.
The bowl-like lower valve of Inoceramus (Volviceramus) grandis reflects an iceberg-like (Thayer, 1975) adaptation to life on a soft substrate, wherein the growing specimen sank progressively lower into the substrate, apparently displacing sufficient carbonate mud to maintain freeboard for the lower valve. Mostly random orientation of epizoic oysters on the upper valve suggests that the plane of commissure was oriented parallel to the mud-water interface and throughout life remained above that surface. The lower valve is thick, smooth, and coiled in the grypheate manner so common in bivalves that lived on soft substrates (e.g., Carter, 1972, p. 333). The upper valve is thinner and coarsely wrinkled in the central area, the rugosity apparently functioning to strengthen the valve much as in corrugated steel roofing.
The platter-like inoceramids, which reached maximum diameters approaching 2 m in Inoceramus (Platyceramus) platinus, reflect a snowshoe-like adaptation (Thayer, 1975) to life on soft carbonate mud. The large bivalves lay flat on the sea floor, with commissural plane oriented horizontally. None of the thousands of specimens I examined shows indication of vertical posture or partial burial. Together with I. (Volviceramus) grandis these bivalves were the principal "islands" for colonization by epizoans during much of Smoky Hill deposition (Fig. 55).
Figure 55--Exposure of middle part of Smoky Hill Member at Locality 23, showing large, articulated specimen of Inoceramus (Platyceramus) platinus encrusted by crowded specimens of Pseudoperna congesta. Hammer is 28 cm long.

Thin-shelled, biconvex, apparently equivalved inoceramids such as Inoceramus balticus were neither heavy enough to sink deeply into the sediment, nor broad enough to rest wholly upon the carbonate-mud surface. Instead, these bivalves lay partially buried in the soft mud and their life attitude was stabilized by concentric growth ridges that characterized both valves. No evidence suggests that these forms were byssate or that they were burrowers. All specimens lie in the chalk with plane of commissure oriented parallel to the substrate surface. I conclude that such inoceramids were free-living, epifaunal nestlers that in essence "floated" like coracles on the soft carbonate mud in the manner reported for unionids by Eager (1974). All of these inoceramids were apparently true filter feeders whose gills strained food from water passing through a slightly gaping commissure.
Radiolitids
The rudist Durania maxima was also a sessile, free-lying (West, 1977) epifaunal bivalve. Specimens occur as isolated valves, or in clusters of a few individuals, in at least four stratigraphic intervals of the member. The right valve is inversely conical, and stood erect when young, but was usually recumbent in the large, adult stage. Specimens reached heights as great as a meter (3-4 ft) according to Logan (1898, p. 494), although those examined by Miller (1968) and in my collection are much smaller. According to Miller (1968, p. 38) no left valves have been recorded. This suggests that the upper valves were very thin and have been destroyed by diagenetic dissolution. It has been suggested that in such forms the mantle extended across the broad rim of the valve and contained intracellular zooxanthellae (Kauffman and Sohl, 1974), as in the modern bivalve Tridacna. Absence of endolithic algae in all Smoky Hill skeletal remains examined in thin sections suggests deposition below the photic zone, and casts doubt on this interpretation. The conical shape of D. maxima indicates that specimens lay partly buried in the bottom, propped in the upright or recumbent position by surrounding mud. The Sternberg Memorial Museum contains a large specimen (13820-2) that clearly lay oblique to bedding, about half buried in the mud. This position is demonstrated by the occurrence of encrusting oysters, the distribution of which marks a clearly defined mud-water interface, with oysters occurring only above that plane. Another specimen in that collection (4093) shows evidence of initial growth, rotation in the mud, and final upward growth at an angle to the initial growth stage. Many collected specimens of Durania have been flattened laterally by compaction, which shows that they had been toppled to a prone position before burial, and one specimen in my collection was broken and encrusted by oysters before burial (Fig. 56). Durania, like the inoceramids, served as substrate for at least three kinds of epizoans. Logan (1898, p. 495) stated that the "type specimens are adult forms united by their entire lengths, one of the specimens having three young ones attached to it near its upper extremity by their lower valves."
Figure 56--Specimen of Durania maxima from lower part of Smoky Hill Member in SW Sec. 21, T. 15 S., R. 26 W., Gove County, Kansas, showing encrustation by Pseudoperna congesta on exterior and interior surfaces, as well as on a broken surface (far left). KU108439, X 112.

My field data suggest that environmental conditions suitable for growth occurred mainly during episodes that fostered development of shelly beds, described above, which represent times of better-circulated bottom waters. Coates and others (in press) have suggested that occurrences of Durania mark marginal Tethyan transgressive pulses, and that one such pulse is represented by the Clioscaphites choteauensis Zone; however, to date I have not recorded a single rudist from that zone in Kansas. Furthermore, the richest accumulations in Kansas occur in the lower part of the Smoky Hill Member, which Coates and others (in press) relate to an episode of regression. The generalization that Durania occurrences reflect transgressive pulses from the Tethyan belt is therefore unsubstantiated.
Ostreids
Ostreid bivalves, mostly assignable to Pseudoperna congesta, are the principal epizoans on Smoky Hill inoceramids and rudists. In my collection scarcely any specimen of Inoceramus (Volviceramus) grandis, I. (Platyceramus) platinus, or I. (Cladoceramus) undulatoplicatus lacks at least partial encrustation by these small oysters. In fact, many articulated inoceramids are encrusted almost completely on the exterior surfaces of both valves. On I. (Platyceramus) platinus the oysters may be largely spat, juveniles, or adults, or may represent as many as four generations of oysters including spat through large adult forms. Oysters of second and third generations may be stacked on first-generation oysters. Where incompletely encrusted, these inoceramids may host uncrowded specimens of nearly circular or oval outline; but where the valves are heavily encrusted the oysters are crowded together, have irregular outlines that reflect competition for space, and may have high marginal walls formed by upward growth when lateral growth was impeded by impingement against neighboring oysters. On many large specimens of I. (P.) platinus the marginal oysters manifest strongly preferred orientation, with longest beak-to-commissure axes arranged perpendicular to inoceramid valve margins (Fig. 57), and with general parallelism to their immediate neighbors. Such oyster arrangement apparently resulted from marginal oysters elongating their shells so as to take advantage of newly created substrate. On other I. (P.) platinus a majority of encrusting oysters may show preferred orientation, not just those at the inoceramid valve margin, a condition suggesting rheotropism, i.e., growth in the direction of approaching currents so as to maximize feeding efficiency (Bottjer and others, 1978). Oyster clusters with radial orientation at inoceramid valve margins, as well as those displaying two or more generations, apparently signify growth on a living substrate. Inoceramids encrusted only by spat or small juveniles were probably moribund or dead at the time of oyster colonization, and were buried before the colonizing oysters could mature. Oyster spat are arranged commonly along depressions between inoceramid growth ridges, as if the oyster larvae selected sheltered places for settlement, but occurrence of spat also on crests of growth ridges or extensively across flat inoceramid valves demonstrates that shelter was not a major requirement for the larvae.
Figure 57--Fragment of I. (Platyceramus) platinus encrusted by crowded specimens of Pseudoperna congesta, which show radial orientation and commissureward elongation of specimens closest to inoceramid valve margin. Lower part of Smoky Hill Member in SW Sec. 21, T. 15 S., R. 26 W., Gove County, Kansas. KU108432, X 112.

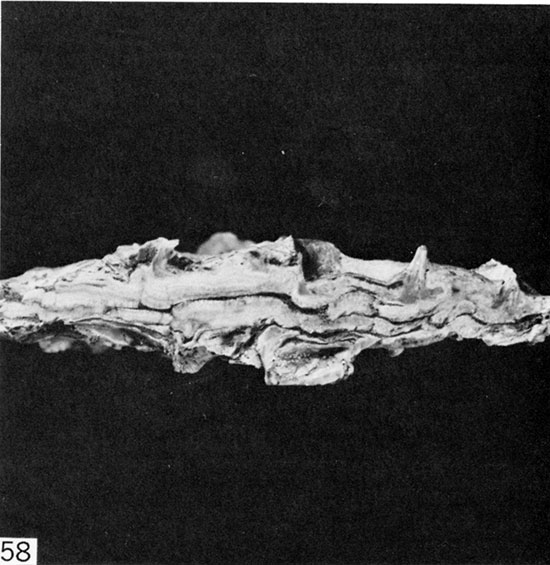
Most perplexing is occurrence of thick encrustations of crowded oyster specimens on external surfaces of both valves in articulated Inoceramus (Platyceramus) platinus (Fig. 58). Possible explanations include (1) the host was oriented vertically, either partly buried in the sea floor or suspended byssally from floating objects such as logs; (2) the host lay flat on the sea floor, oysters colonized the upper valve, and the inoceramid was then overturned by large predatory organisms in search of food (Hattin, 1965, p. 21); (3) same as (2) but inoceramids were overturned by waves or currents; or (4) the host lay flat on the sea floor and was colonized simultaneously on both upper and lower valves. The first hypothesis is rejected because (a) not a single specimen of I. (P.) platinus is preserved in the erect position, (b) the valves preserve no evidence of byssal attachment (Erle Kauffman, oral communication, 1977), and (c) logs, which are common in Greenhorn and Fairport strata of Kansas, and were thus readily preservable in chalky beds, are exceedingly rare in Smoky Hill strata. The second hypothesis is rejected because (a) evidence of predation (e.g., bite marks, extensive shell breakage, debris in coprolites) is sparse, (b) in some beds all articulated I. (P.) platinus are encrusted on both valves and the likelihood that all of these were overturned by predators is highly improbable, (c) chalk laminae show no sign of deformation such as would occur if the inoceramids had been overturned. The third hypothesis is also rejected because evidence for sufficiently strong waves or currents (e. g., skeletal lag concentrates, scour-and-fill structure) is lacking in Smoky Hill strata. The fourth hypothesis is therefore accepted as most plausible. Despite possible problems attending this hypothesis, one must remember that when deposited the chalk-forming mud initially consisted mostly of water, so that oysters facing downward into the mud actually lived in a predominantly aqueous environment. The hypothesis gains strength from observation of the same phenomenon in specimens of the highly inequivalved Inoceramus (Volviceramus) grandis, the large, bowl-shaped, downward-facing lower valve of which is usually preserved in situ and which in many large specimens collected is encrusted completely by crowded specimens of Pseudoperna congesta. My collection also contains a short, but broadly expanded specimen of Durania maxima the underside of which was colonized by numerous specimens of P. congesta (Pl. 7, 2-3). The encrusted part of this rudist can in no way be interpreted as having stood above the bottom muds.
Figure 58--Vertical section through paired valves of articulated I. (Platyceramus) platinus, showing encrustation by Pseudoperna on exterior surfaces of both valves. Middle part of Smoky Hill Member in SW Sec. 1, T. 14 S., R. 26 W., Gove County, Kansas. KU108433, x 1.
Specimens of Inoceramus (Volviceramus) grandis also were host to large numbers of Pseudoperna congesta. The upper valve, or "lid," is invariably encrusted externally by oysters, and these epizoans commonly cover the surface completely. In some I. (V.) grandis, adult oysters cover the entire upper valve, with juveniles and spat stacked, in succession, on the larger specimens. In other I. (V.) grandis, large oysters are confined to the central area, and only juveniles or spat occupy marginal areas of the valve. Lower valves were crushed during compaction, so that collected materials are fragmented. Some thin lowervalve fragments from smaller specimens may have large areas that bear few epizoans, but the largest lower-valve fragments in my collection, representing thick-shelled, fully mature individuals, are entirely encrusted by P. congesta that are mostly adults. Because of the large size and thick, heavy shell the lower valves of adult I. (V.) grandis apparently lay deeply buried in the soft carbonate mud and many of the associated oysters apparently lived facing into this mud. I have not recorded large specimens that have a substantial part of the lower valve free of these epizoans, nor have I recorded a large lower valve that had been overturned.
Epizoans are rare on Inoceramus balticus. Specimens of small oysters are associated with a few I. balticus, but confirmation that these were actually attached to the inoceramid is lacking.
Pectinids
Specimens of the Pectinidae named Pecten bonneri by Miller (1968) are of uncertain stratigraphic position, and he made no mention of faunal associates or epizoans. Small pectinids that I collected from the top of Marker Unit 21 at Locality 21 lie free in the chalk and lack epizoans. It is likely that all of these pectinids were pelagic forms, which were uncommon in Kansas during deposition of the Smoky Hill Member.
Bakevelliids
A species of Phelopteria, referred to as Pteria cf. P. petrosa (Conrad) by Miller (1968, p. 35) and here called Phelopteria sp. A, occurs as free-lying specimens in chalky strata of the Clioscaphites choteauensis Zone, to which it is apparently restricted. A second form, here called Phelopteria sp. B, was recorded in the upper part of the Inoceramus (Cladoceramus) undulatoplicatus Zone at Locality 13. According to Kauffman and Powell (1977, p. 51) Phelopteria is a "byssate, commonly pendant, epifaunal bivalve," which probably was attached to inoceramids or erect algae, or may even have been epiplanktonic. Because the genus occurs in beds that are notably rich in cephalopods, and because specimens are very rare, the possibility of an epiplanktonic habit is attractive, but cannot be proved on the basis of available evidence.
Cirripeds
Smoky Hill cirripeds recorded to date include the borings of acrothoracicans and stalked forms with armored peduncles that belong to the two lepadomorph families Scalpellidae and Stramentidae. The stramentids are attached to valves of Inoceramus (Volviceramus) grandis, I. (Platyceramus) platinus, cephalopods or, rarely, oysters and rudists. Stramentum haworthi and Zeugmatolepas sp. are the most common species. The former occurs mostly as juveniles on inoceramid valves, as juveniles on living chambers and septate portions of Clioscaphites choteauensis, and as an adult (holotype specimen only) on Pseudoperna congesta. Adult specimens of S. haworthi have been recorded mostly on molds of a large, smooth species of Baculites from several places, including the middle and upper parts of the Smoky Hill Member of Kansas, the upper chalk member of the Smoky Hill at Pueblo, Colorado (Scott and Cobban, 1964), and from near Irene, South Dakota (W.A. Hasenmueller, oral communication, 1973). Miller (1968, p. 61) states that S. haworthi was "pseudoplanktonic," not benthic, but the presence of this and other lepadomorphs on inoceramid valves, a rudist, and an oyster demonstrates benthic existence and suggests that specimens on cephalopod molds may have settled on empty conchs that lay on the sea floor. I regard S. haworthi as a benthic organism. Zeugmatolepas sp. and other scalpellids, represented by articulated and disarticulated skeletons, are associated almost exclusively with inoceramids and oysters and are also interpreted as benthic forms. Numerous articulated specimens of Zeugmatolepas sp. are preserved on the right valves of oysters attached to the upper valve of an Inoceramus (Volviceramus) grandis specimen that was collected from just below Marker Unit 5 at Locality 12. In addition, J. D. Stewart (written communication, 1982) has recorded the occurrence, in large numbers, of Zeugmatolepas on downfacing valves of some I. (P.) platinus. These are clearly benthic forms. The acrothoracican borings, assignable to the genus Rogerella, are common throughout the lower half of the Smoky Hill Member, and occur locally in great profusion (Fig. 59). The borings are preserved on inner and outer surfaces of inoceramid and rudist valves as well as on valves of Pseudoperna congesta. In some inoceramid valves rough alignment of borings in rows parallel to growth lines of the host suggest that the larvae settled in grooves for reasons of protection. However, many borings are on growth ridges (Fig. 59), thus dispelling the notion that sheltered settlement sites were a requirement for these boring cirripeds. Common occurrence of aerothoracican borings in oysters suggests that oyster colonization of inoceramid hosts occurred before acrothoracicans established themselves, but in a few inoceramids the acrothoracican borings were vacated, then occupied by oyster spat.
Figure 59--Valve fragment of Inoceramus ?(Platyceramus) platinus, showing profusion of acrothoracican cirriped borings (Rogerella). From lower part of Smoky Hill Member in SW Sec. 16, T. 14 S., R. 26 W., Gove County, Kansas. KU108434, X 1.

All of the cirripeds were suspension feeders, which strained food from the water column by means of appendages that projected from the capitulum.
Uintacrinids
The nature of Uintacrinus occurrences has been described in elaborate detail by Springer (1901). Specimens occur in very thin lenses of well-cemented limestone, with the most perfect specimens preserved only on the undersides. In each lens, overlying specimens are crushed, disarticulated, and cemented together in jumbled fashion. Springer (1901) described one slab 12 mm thick and 15.2 m wide that contained 1200 mostly adult specimens on the underside (Pl. 9, 2). All known Kansas specimens occur in aggregations of many individuals, and must, according to Springer (1901, p. 12), have lived in free-swimming swarms like the living stalkless crinoids. Springer's description of occurrences suggests that the species is restricted to a small stratigraphic interval that is within the zone of Clioscaphites choteauensis. The occurrence of such large numbers of individuals in solitary lenses suggests that the crinoids lived in separate schools and were killed during one or more mass-mortality events. Beecher (1900, p. 268) notes the preferred orientation of crinoid arms and an associated Baculites mold, which suggests that a gentle current prevailed at the time of settlement on the sea floor. The contrast in preservation of specimens on undersides of limestone lenses versus those above suggests further that the lowermost, buried specimens were better protected from decay, probably in an anaerobic situation, whereas those higher in the lenses decayed and fell apart before burial (cf. Kauffman, 1978, p. 35).
Cephalopods
The sparse examples of teuthids and belemnoids in Smoky Hill strata may be evidence of their general scarcity in the Western Interior Sea. It is possible that these forms were preyed upon heavily by large pelagic organisms, such as mosasaurs, turtles, sharks, or teleosts. Indeed, remains of teuthids have been recorded in several vertebrate coprolites U. D. Stewart, written communication, 1982). Because living coleoids are all nektonic forms we have little reason to believe that those preserved in the chalk beds were not also swimmers in the open sea.
The sparse examples of large, spirally coiled ammonites belonging to the Collignoniceratidae may also have been rare in the Western Interior Sea. If they were a food source for large predators such fate is unsubstantiated by Smoky Hill specimens that preserve bite marks. At least occasional vertebrate predation on ammonoids is suggested by scaphite remains in a coprolite from the Clioscaphites choteauensis Zone (Stewart and Carpenter, in prep.).
In the absence of clear evidence for widespread shell destruction by predators, and because other cephalopods (Baculites, Clioscaphites) are common in some stratigraphic intervals and thus prove the ready preservability of ammonoids in the chalk, I conclude that the scarcity of non-heteromorph ammonoids is owing to environmental conditions. The few collignoniceratids that have been reported to date are probably strays from farther south, having drifted in small numbers into the Kansas area before death, or even after death. Belemnoids, which are characteristic of the so-called "Boreal realm," are rare not only in Kansas but also are very localized and mostly rare even in deposits of the Western Interior Sea in western Canada (Jeletzky, 1971, p. 1653). These belernnoids apparently are strays from the most northerly regions of North America.
The life habits of the scaphites and baculites are mostly speculative because comparable forms are not alive today. Regarding scaphitoids, a wide range of views is manifest in such works as those by Diener (1912, p. 78), who believed that Scaphites was perhaps a crawling form; by Scott (1940, p. 307), who suggested "a nectobenthonic crawling or swimming habit of limited extent"; and by Trueman (1941), whose studies of buoyancy centers suggested for scaphitoids a floating habit, with aperture facing upward, at least at rest, and possible life as plankton feeders at the surface. Similar diversity of opinion has been expressed with regard to the life habits of Baculites. The controversy has been summarized by Klinger (1981), who suggests that "a horizontal swimming position for orthoconic ammonites must be viewed with extreme caution." Clioscaphites and Baculites were probably poor swimmers, like most other ammonoids (Lehman, 1975; Mutvei, 1975; Kennedy and Cobban, 1976, p. 81). Heptonstall (1970) has implied that most ammonoids were able to control buoyancy by addition or removal of water from the living chamber, although this may have resulted only in slow vertical movements in the water column. He postulated that forms with a small siphuncle, which would include the scaphites, may have been restricted to narrow depth range and many, perhaps all, such forms were probably benthic (Heptonstall, 1970, p. 324). Indeed, Lehmann (1975) suggested that all ammonites were benthic forms. In marked contrast is the opinion of Mutvei (1975) that ammonoids were pelagic organisms that occupied the upper 1000 m of oceans and were probably capable of large diurnal migrations in the water column. Westermann (1975) believes that this figure is too large by a factor of five, which implies maximum depth range of only about 200 m. The contradictory opinions regarding ammonoid paleoecology thus continue to modern times and make impossible any realistic assessment of position in the water column or depth relations of Smoky Hill heteromorphs.
Kennedy and Cobban (1976, p. 46, 47) state that Clioscaphites choteauensis has a wide geographic distribution in the Western Interior and occurs in lithology of wide variety, from chalk to sandstone. This distribution does not indicate strong facies control or intimate association with the bottom. Abundance of C. choteauensis in Smoky Hill strata at several localities shows that this species, at least, was well adapted to environmental conditions that prevailed during deposition of that zone.
The large, smooth species of Baculites recorded in Plate 8 is abundant only in certain stratigraphic intervals of the Smoky Hill chalk, including Inoceramus (Cladoceramus) undulatoplicatus, Clioscaphites vermiformis, and C. choteauensis Zones. This distribution implies that conditions were favorable for baculite proliferation only sporadically. From the C. vermiformis Zone upward beds containing few benthic macroinvertebrates are also deficient in Baculites. Apparently conditions favorable to benthic macroinvertebrates were favorable also to Baculites, which implies that Baculites distribution was controlled more by benthic environment than by that of superjacent parts of the water column. I regard Baculites sp. (smooth) as a nektobenthic form.
Little is known about the feeding habits of ammonoids, but recent studies by Lehmann and Weitschat (1973) show that in genera studied the crop contains foraminifers, ostracodes, crinoid remains, and small ammonites.
Trace Fossils
The only common evidence of infaunal macroinvertebrates in Smoky Hill strata is burrow structures produced by deposit-feeding, worm-like organisms (Fig. 40, 41). Most such structures are compressed-ellipsoidal in vertical section and the larger ones, up to 1.3 cm in maximum dimension, are probably referable to Planolites. Slender burrows, possibly referable to Chondrites, were recorded sparingly only at Locality 13. Forms such as Trichichnus, Zoophycos, and Teichichnus, which are abundant in underlying Fort Hays beds, were not recorded in the Smoky Hill. In the lower half of the Smoky Hill, beneath the top of Marker Unit 11, bioturbated intervals are common, usually standing out as light-colored bands in fresh exposures or as resistant ledges in weathered exposures. Average insoluble residue content of these beds is 24 percent as compared to 34.3 percent for nonbioturbated, nongranular chalks from the same interval. The more pure beds of carbonate mud may represent slightly firmer substrates than normal, which were at least marginally suitable for inhabitation by the burrowers. Above Marker Unit 11, evidence of bioturbation is rare and consists mostly of sparse, minute structures that occur in beds of granular chalk. An exception is the thoroughly bioturbated calcarenite lens recorded between Marker Units 21 and 22 in SE Sec. 1, T. 14 S., R. 31 W., Gove County. It is probably no coincidence that the part of the section in which benthic macroinvertebrates are sparse to rare also lacks the obvious kinds of bioturbated intervals that are so common in the more fossiliferous part of the member. The two lines of evidence suggest that during much of upper Smoky Hill chalk deposition the bottom was oxygenated marginally, and in fact may have approached an anaerobic condition.
Infaunal Body Fossils
Except for rare molds of Lucina sp. A (Miller, 1968; this paper), reported from the zones of Inoceramus (Cladoceramus) undulatoplicatus and Clioscaphites choteauensis, Smoky Hill strata lack preserved remains of infaunal body fossils. This fact might reflect diagenetic removal of originally aragonitic shells, such as characterize most infaunal bivalves, or may indicate that infaunal organisms, other than trace-making worms(?), were excluded from these deposits for environmental reasons. The first possibility is rejected because the Smoky Hill contains numerous baculitid and scaphitid fossils, the aragonitic skeletons of which were removed during early diagenesis, but are nevertheless very well preserved as molds. The second possibility is supported strongly by the absence also of body fossils from calcitic, infaunal forms such as echinoids. Furthermore, burrow traces attributable to infaunal bivalves or echinoids have not been recorded in Smoky Hill strata. Possibly the carbonate ooze was generally too fluid to support infaunal, skeleton-secreting organisms. I disbelieve that anoxic interstitial conditions were a principal factor, because many infaunal organisms maintain siphonal contact with the benthic waters, which were sufficiently well oxygenated to support an abundance of epifaunal life such as inoceramids and oysters during much of Smoky Hill deposition. The lucinids reported by Miller (1968, p. 37) occur in strata that are rich in other benthic and nektobenthic forms, and that probably represent an episode of exceptionally favorable benthic environments.
Community Structure and Habitat
Smoky Hill macroinvertebrates comprise low-diversity communities, the basic elements of which were large inoceramids and vast numbers of small ostreids that encrusted them. These organisms made up the minimal preservable assemblage and are virtually the only forms present in many parts of the composite section. In an environmental setting involving soft, oozy substrates and poorly circulated bottom waters, the inoceramids established themselves by virtue of growth forms that prevented sinking beneath the mud-water interface, and served as "islands" upon which oyster spat settled and commonly thrived. At times, bottom circulation improved sufficiently to foster proliferation of inoceramids and their epizoic oysters, and addition to the limited benthos of rudists, acrothoracic and lepadomorph cirripeds, and occasionally sponges?, lucinids, and serpulids. Conversely, when circulation decreased below the norm, the Smoky Hill macroinvertebrate benthos diminished nearly to zero. That improved circulation of bottom waters was a major factor controlling diversity increase is manifest in the lithologically similar Fairport Chalk Member, Carlile Shale (Hattin, 1962), in which strata containing well-washed calcarenites include not only large inoceramids and abundant ostreids, but abundance also of cirripeds, serpulids, and bryozoans.
In Smoky Hill assemblages a high proportion of oysters and inoceramids have articulated valves, and the rudists and especially the larger inoceramids are preserved apparently in life positions, the inocerarnid-ostreid or inocerarnid-ostreid-cirriped association was recorded repeatedly throughout the composite section, pre-burial shell fragmentation was minimal, and along individual horizons wide size range is evident among individuals of a given species. These features suggest that Smoky Hill assemblages represent fossil communities in the sense of Fagerstrom (1964). The only common modification of original hard-part distribution is the separation, probably by weak bottom currents, of some small, thin right oyster valves from the larger, attached left valves. This accounts for scattered occurrence of isolated right valves of ostreids throughout the member, but does not necessitate classifying the assemblages as residual communities (Fagerstrom, 1964, p. 1202).
Except for burrows of deposit-feeding worms(?), Smoky Hill benthic macroinvertebrates were exclusively suspension feeders. Among paleoecologists there is widespread belief that muddy biotopes are (were) generally unsuited for inhabitation by suspension feeders (e.g., Purdy, 1964; Levinton and Bambach, 1970, p. 97; Carter, 1972, p. 329; Thayer, 1974, p. 135), and that faunas associated with fine-grained deposits are (were) mostly infaunal deposit feeders. Evidence from macroinvertebrate body-fossil assemblages of the Greenhorn, Fairport, and Niobrara chalks demonstrates need for revision of this concept, and exceptions to the generality' stated above have been noted by Walker (1974). Paucity of suspension feeders in muddy environments has been attributed to such factors as clogging of filtration systems and sinking into soft substrates (Thayer, 1974, p. 135). Morphologic adaptation of Smoky Hill inoceramids and rudists to soft substrates has been noted above, and the occurrence of vast numbers of suspension feeders, to the complete exclusion of deposit feeders (other than worms(?)), shows that the water was rarely, if ever, sufficiently turbid to be a controlling factor in trophic composition. The constant rain of food supplies, especially including plant debris, fecal pellets, and plankton, perhaps distributed by gentle bottom currents, was more than adequate to sustain communities of suspension-feeding organisms. Absence of deposit-feeder body fossils in Smoky Hill strata poses problems with respect to currently accepted concepts. Fine-grained deposits normally contain more organic matter than those that are coarse grained (Trask, 1939, p. 433; Purdy, 1964, p. 243; Levinton and Bambach, 1975, p. 98; Stanton and Dodd, 1976, p. 328). The abundance of fecal pellets and particulate organic matter in Smoky Hill rocks indicates that absence of deposit-feeding bivalves was not owing to food shortage, which has been suggested as a controlling factor by Wright (1974, p. 432). Furthermore, the preservation as molds of lucinids, scaphites, baculites, and collignoniceratids in the composite section makes unlikely the possibility that diagenetic removal of aragonitic skeletons accounts for absence of deposit-feeder skeletons in the Smoky Hill. As noted by Hallam (1976, p. 251), infaunal or semi-infaunal bivalves required a moderately firm muddy substrate. Exceedingly soft substrates, possibly combined with marginally oxygenated bottom waters, seems to account for absence in Smoky Hill deposits of infaunal deposit feeders other than worms(?).
Despite the presence in Smoky Hill strata of shell-crushing shark remains, such organisms were apparently nowhere abundant. During my study numerous remains of bony fish and aquatic reptiles were noted, but teeth of shell-crushing sharks are scarce, indeed. Evidence of predation, such as bite marks on bivalve shells, coprolites filled with shell debris, and scatterings of broken shells, are sparse in the Smoky Hill. Predation on bottom dwellers thus does not appear to have been a major element of community life in the Smoky Hill environment. Greatly elongated, almost funnel-shaped ostreids occur locally in the section. These apparently attached themselves to small shell fragments that sank gradually under the increasing weight of the rapidly upward-growing oysters. Such shell fragments may be the result of predation, but oysters having small attachment surfaces and elongate growth forms are common only in a few parts of the Smoky Hill. Growth to giant size of so many inoceramids and rudists indicates slow depositional rates. This conclusion is supported by development on these organisms of as many as four generations of epizoic oysters. Obviously, the inoceramids and rudists were able to maintain an exposed position for many years. Heavily encrusted hosts may eventually have sunk beneath the mud surface, as suggested by the fact that the youngest oyster generation consists usually of spat or juvenile forms. More rapid rates of deposition may be indicated by large inoceramids on which only oyster spat are preserved, but such occurrences could be explained also as evidence of dead or dying individuals that could no longer remain "afloat" on the mud surface.
The duration of Smoky Hill deposition was approximately five million years (Obradovich and Cobban, 1975, p. 50), and total thickness of my composite section is approximately 180 m. This equates to an average depositional rate of approximately 0.036 mm/yr for Smoky Hill deposition, which is very close to the figure (0.038 mm/yr) calculated by Fischer (1980, p. 100) for the same unit at Pueblo, Colorado. These figures confirm that rates of Smoky Hill deposition were slow, and help to account for the multiple generations of small oysters that encrust large inoceramids through most of the member.
Prev Page--Deposition || Next Page--Gas Production, Summary
Kansas Geological Survey, Geology
Placed on web Feb. 20, 2015; originally published Dec. 1982.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/225/08_paleo.html