![]()
Prev Page--Shell Morphology || Next Page--Family MYALINIDAE
Systematic Paleontology
The Earliest Mytilacea
Biologists are pretty well agreed that only the later ontogeny is a reasonably reliable index to the phylogenetic history of animals. Also, form may be profoundly affected through adaptation so that animals having similar shapes may not be closely related. Nevertheless, ontogenetic changes in form in the various classes of mollusks have proved to be of great supplemental value in establishing near relationships. Convergences in shell form due to adaptation, or other causes, can be commonly recognized and interpreted by reference to shell ontogeny as revealed by growth lines. Unrelated shells that may look identical at maturity are likely to appear quite different in the early growth stages. Even the discovery that some mollusks underwent most of the evolutionary change in the larval stages (proterogenesis) does not alter the established usefulness of the principles of palingenesis.
Even though Jackson (1890) long ago showed the importance of ontogenetic studies in working out phylogenies of pelecypods, only a few paleontologists have subsequently given attention to this contributory line of evidence. In existing classifications of the pelecypods are found many instances in which shells differing markedly in shape are classed together. Critical ontogenetic studies of these forms may reveal that they are polyphyletic. Extensive consideration of the problem indicates the following generalizations: (1) Some of the major tribes of pelecypods diverged early and have undergone very little differentiation in form since the Medial or Early Paleozoic; such persistence and conservatism of form is recognized to some extent in existing classifications, viz., Pectinacea, some of the Pteriacea, Trigoniacea, and others. (2) Where there has been marked differentiation in form within a single tribe, the changes have been wrought in response to adoption of a radically new type of life, such as cementation of one valve in Chama and Myochama, or adoption of the boring habit, as in Lithophaga and Teredo. (3) In the majority of superfamilies, or even families, the characters of the hinge (ligament and dentition), shell microstructure, ornamentation, and musculature show greater potentialities for variation than do certain broad features of external configuration of the shell.
To be wholly consistent with the above conclusions I should class the great host of Paleozoic shells having the mytiloid (or "modioloid") shape with the Mytilacea. To do so, however, would be very decidedly premature in view of the fact that some of the Paleozoic genera of the Ambonychiidae, Pleurophoridae, and others, while having the general appearance of Mytilacea, exhibit certain features of dentition that have caused the exclusion of these shells thus far from that superfamily in the majority of classifications. Also, some of these shells are obviously closely related to others having a pterioid aspect. There is a very marked resemblance between certain Ambonychiidae and Myalinidae, as for instance Anomalodonta Miller and Orthomyalina, n. gen., or Plethomytilus Hall and Selenimyalina, n. gen. In absence of information on the ontogenies of any ambonychiids, it is not at present possible to determine whether the resemblance is homeomorphic or is really indicative of close relationship.
In form and hinge characters the genus Modiolopsis Hall as represented in the Ordovician must be very like the ancestor of all of the Mytilacea. Modiolopsis is so closely similar to living Volsella ("Modiolus") that nobody has yet satisfactorily shown any difference. Dall (1895, pp. 527-528) was hardly justified in following Fischer in recognition of a separate family, the Modiolopsidae, in view of the absence of any known characters by which the Mytilidae and Modiolopsidae can be separated consistently. It is true that the Modiolopsidae are supposed to be invariably isomyarian, but there are living species of Volsella, for instance, which are also isomyarian. If, however, the shell microstructure in Modiolopsis differs from that of Mytilus and Volsella, I think that it might advantageously be placed in a separate family.
Any adequate understanding of the Early and Medial Paleozoic mytiloids must await further original investigations, made with particular attention to ontogenies and shell microstructure. At the present time it seems to me likely that the majority of Paleozoic genera referred to the Ambonychiidae, Mytilidae, Modiolopsidae, Myalinidae, and Modiomorphidae, as well as many unresolved genera, should be referred to the Mytilacea.
Superfamily MYTILACEA Ferussac
Family MYTILIDAE Fleming
Shell relatively thin, equivalve, obliquely elongated, with beaks at or near the relatively small anterior end; posterior margin broadly rounded, ventral margin with a slight byssal gape, at least in the young; ligament marginal or submarginal, parivincular, opisthodetic; hinge edentulous, or with cardinal teeth 1/(2a,2b), or taxodont both before and behind the ligament in direct correlation with any radial sculpture that is present; inner shell layer composed of aragonite, either homogeneous or finely prismatic, outer layer consisting of calcite, either homogeneous or finely prismatic, the prisms and optie axes more or less tangent with the shell surface and directed radially with respect to the beaks (Bøggild): musculature characteristically anisomyarian, but with a few monomyarian (some Mytilus) and isomyarian (some Musculus) species.
Remarks—A good many Paleozoic shells that should be classified in other families have in the past been placed in the Mytilidae. Some of the Ambonychiidae and all of the Myalinidae have at one time or another been classed as mytilids by several authors. Indeed, homeomorphy is so prevalent in these three families that it is not surprising that the genera Mytilus, or "Modiolus" (Volsella), have frequently been cited from rocks as old as the Silurian and Devonian. Several of the myalinas are so nearly identical with Mytilus and Volsella in form that they cannot be differentiated without some knowledge of the hinge characters (ligament) and shell microstructure. It is significant that the critical differences in shell structure between Myalinidae and Mytilidae commonly can be distinguished directly with a hand lens or low-power microscope, without recourse to thin-sections or polished surfaces. The peculiar radial-tangential prisms of the outer calcite shell layer in many genera of the Mytilidae produce a very finely striate appearance, sufficiently coarse in some instances to be mistaken for radial ornamentation. In a few instances this surface texture is reflected in good external molds. All of the Paleozoic species herein described under the Mytilidae clearly show this characteristic feature.
In the Myalinidae the outer layer of calcite is composed of short, polygonal prisms or fibers which, in the right valve, invariably are coarse enough to be seen at 10 diameters magnification. Unlike the prisms in the Mytilidae those of the myalinas are perpendicular to the shell surface instead of tangential to it. Because the prisms in both families are composed of calcite, they generally are well preserved in specimens which are not too badly recrystallized.
Genus MYTILUS Linnaeus, 1758, emend.
Mytilus Linnaeus, 1758, Syst. Nat., Ed. 10, p. 704.
Genotype (by subsequent designation), Mytilus edulis Linnaeus, Gray, Zool. Soc. London, Proc. for 1847, p. 198, 1847. Geologic range of genotype, Pliocene to Recent; cosmopolitan.
Shell elongate, thin, with terminal, pointed beaks, anterior lobe and anteroventral sinus very obscure or lacking; surface generally smooth, or with radial ribs, which may be broad and indefinite, or fine and uniform; hinge edentulous, or with cardinal teeth 1/(2a,2b) or with variable number of taxodont teeth retained from the embryonic provinculum.
Range—Mesozoic to Recent.
Genus VOLSELLA Scopoli, 1777, emend.
Volsella Scopoli, 1777, Introd. Hist. Nat., p. 397.
Modiolus Lamarck, 1799, Mem. Soc. Hist. Nat. Paris, p. 87.
Genotype (by subsequent designation), Mytilus modiolus Linnaeus, Gray, Proc. ZooL Soc. London, for 1847, p. 198, 1847. Geologic range, Pliocene to Recent; cosmopolitan.
Shell highly convex, elongate, thin, with subterminal, rounded beaks, which commonly rise slightly above the hinge line; more or less prominent anterior lobe bounded by broad anteroventral sulcus extending ventrally to broad byssal sinus at middle of anteroventral margin; surface generally somewhat rugose from projecting growth lamellae, or with fine radial ribs; hinge edentulous in adults.
Range—Mesozoic to Recent.
Remarks—Grant and Gale (1931, p. 248) have shown that legally the name Volsella has unquestioned priority over the more familiar term Modiolus. Both genera are based on the same genotype and therefore are absolute synonyms. It is unfortunate that such an authority as Thiele (1935, p. 798) refuses to recognize the prior claim of Volsella on the grounds that Scopoli misunderstood the characters of the shell which later became the genotype. An author's ideas about his genus have no legal standing whatsoever where they are at variance with the characters of the type species.
In these days paleontologists have a distaste for long-ranging genera and species, and many follow the practice of evaluating observed characters of shells in terms of their stratigraphic position, and in many instances very similar fossils have been given different names to better emphasize unlike stratigraphic occurrences. Where the stratigraphic measure is employed, however, in differentiating fossil units, the classification should by all means be based on proven shell characters and not simply on suspected distinction.
Painstaking research on the material available to me reveals certain obscure but consistent characters by which the Paleozoic forms herein described under the Mytilidae can be distinguished from the majority of similar Mesozoic and Cenozoic forms whose generic affinities with Volsella (or Modiolus) have not seriously been questioned. One would suppose, a priori, that a comprehensive genus like Volsella, under which there are so many hundreds of described species, both fossil and Recent, is poorly defined, possibly polyphyletic, incorporating as it does shells that vary considerably in external details. It must be noted, however, that variation in form within the genus is limited, so that modern species can nearly be matched in the fossil faunas from all of the geologic systems back at least to the Jurassic. It seems that Volsella is another example of those remarkably virile, conservative tribes of animals which, like Lingula and Limulus, has undergone little differentiation for vast ages in earth history.
Differentiation in Paleozoic Mytilidae
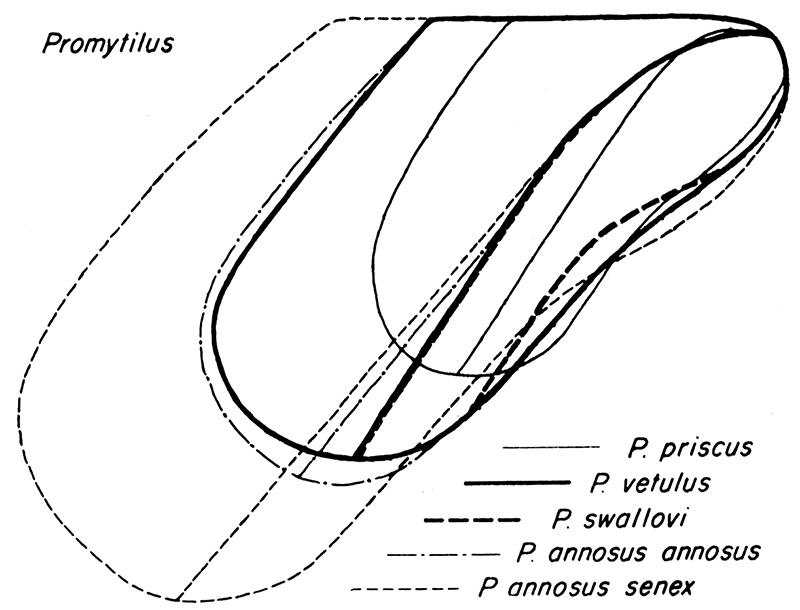
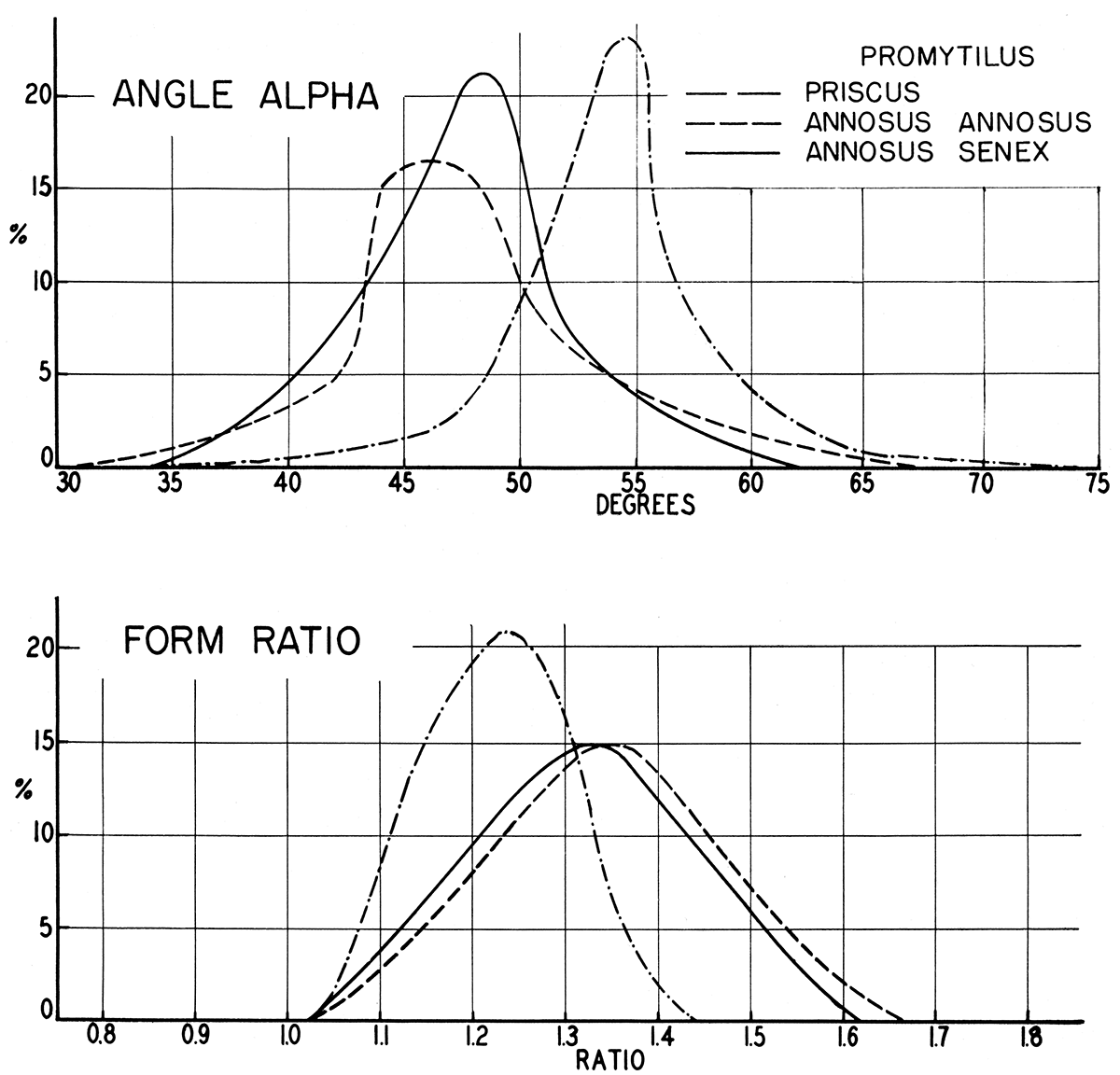
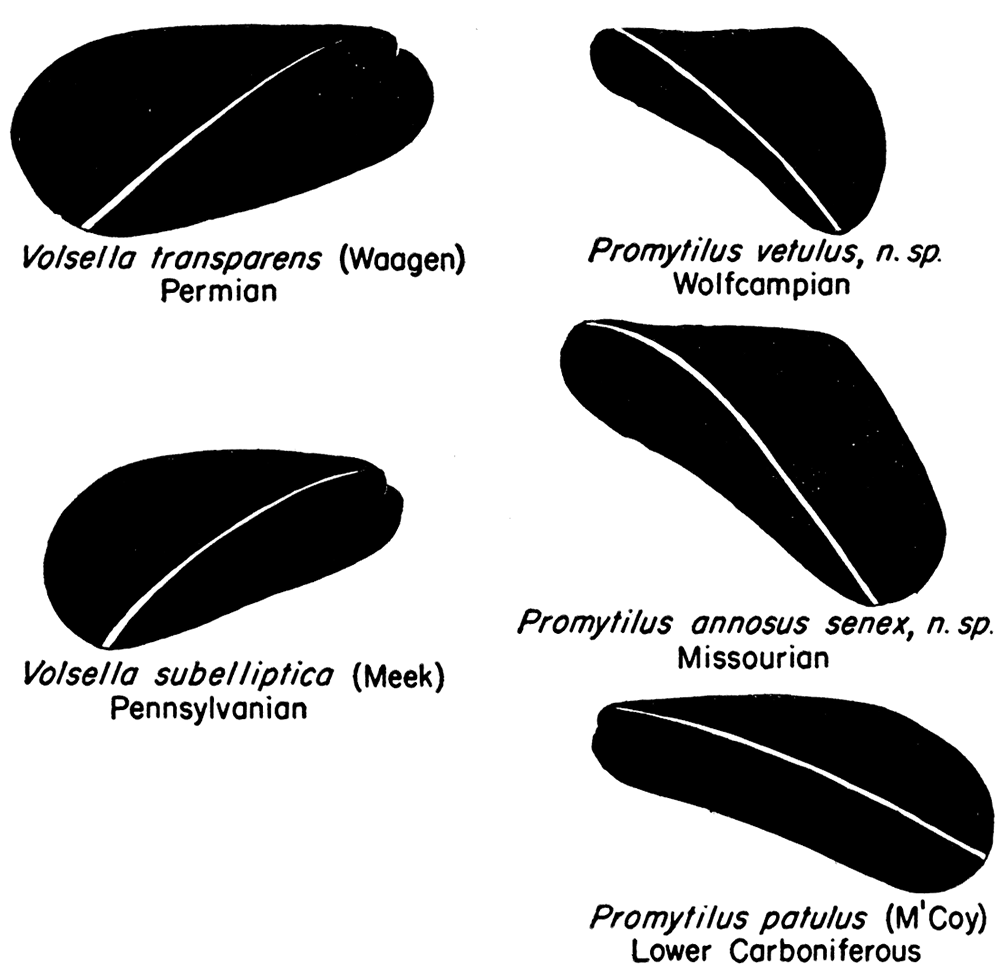
Disregarding the pre-Carboniferous species for the moment, two distinct groups of Volsella-like mytilids are recognizable in Late Paleozoic rocks. One of these, characterized by Lower Carboniferous forms like "Modiolus" patulus M'Coy, Lithodomus lingualis Phillips, Lithodomus lithodomoides R. Etheridge, Jr., from the British Isles, and Upper Carboniferous and Permian species, such as the American Myalina swallovi McChesney, Promytilus annosus, n. gen., n. sp., P. priscus, n. sp., and P. vetulus, n. sp., is characterized by shells of rather marked convexity, having a prominent, though broadly rounded umbonal ridge, and distinctly indented ventral margin. The beaks are nearly, but not quite, terminal, being in this respect somewhat intermediate between the type species of Volsella and of Mytilus. The anterior lobe, although poorly defined in some of the Paleozoic species, is more strongly developed than in typical Mytilus. This group of species seems to constitute a true gens and deserves a separate generic name. The new generic name Promytilus is proposed for this group. In the Carboniferous some of the younger species of Promytilus tend to have a relatively greater angle α and, correspondingly, a greater relative height than the older species, seemingly paralleling the marked change in form in the evolution of the Myalinidae. For instance, the Irish species "Modiolus" patulus, of Viséan age, which is here assigned to Promytilus, has a much longer shell (length/height, 2.15), and smaller angle α (30°) than any known similar Pennsylvanian or Permian species.
The second group of species, typified by "Modiolus" subellipticus (Meek) from ·the American Pennsylvanian, and "Modiolus" transparens (Waagen), from the uppermost Permian of the Salt Range, is certainly distinct in having flatter valves, beaks distinctly posterior to the anterior margin, and only a very obscure indentation of the ventral margin. The angle α is consistently small in these forms (40°); and, judging from the trends in other related groups, these species probably are more primitive, more like the ancestral stock of Volsella, than the Promytilus alluded to above. Furthermore, the crest of the umbonal ridge is narrow, in some instances quite angular, a circumstance not noted in Volsella, Mytilus, or Promytilus. Although specimens of the "Modiolus" subellipticus group are rare, a comparatively large proportion of the specimens known to me possess both valves intact, which suggests that these forms preferred an environment lacking current or wave action competent to dissociate the valves of dead individuals. This situation is in marked contrast to specimens of the Promytilus patulus gens in which bivalved specimens are extremely rare. For "M." subellipticus and related species the generic name Volsellina is proposed.
Judging from the general appearance of shells of the Promytilus patulus group, it is not at all unlikely that this stock contains the ancestry of the typical Mytilus of Mesozoic and later time. It is highly probable that some of the citations to Mytilus in the Paleozoic are to species that should be referred to the Myalinidae. The Mesozoic and Cenozoic volsellas, with their beaks placed well back of the front margin of the shell, are more like the Volsellina subelliptica group.
In the Devonian, the genus Mytilops Hall seems to be a near relative of Volsellina, being characterized by a very elongate form and a relatively small angle α. Mytilops is inadequately known, so that critical comparison with Late Paleozoic species is not possible.
Unfortunately, my collections are relatively extensive for only the Missourian, Virgilian, and Wolfcampian divisions of the Late Paleozoic. Nowhere abundant, specimens of Volsellina and Promytilus are especially rare in the Lower Pennsylvanian and later Permian rocks of North America, and I have had no opportunity to study material from these parts of the geologic column.
Genus PROMYTILUS Newell, n. gen.
Genotype, Promytilus annosus Newell, n. sp. Geologic range of genotype, Missourian series, Pennsylvanian.
Shell elongate, thin, with terminal beaks, and a slender anterior termination of the shell, corresponding to the beaks and anterior lobe; lobe well defined but not extended anteriorly beyond the beaks, set off from main part of shell by a broad but well defined sulcus, which extends from the beaks backward to the ventral margin where it coincides with a broad, clearly marked sinus in the shell margin; umbonal ridge well defined, rounded; shell microstructure and hinge as in Mytilus.
Range—Mississippian to Permian, Mesozoic?
Remarks—For a time I debated using Mytilus or Volsella for the Late Paleozoic shells commonly referred to Myalina swallovi. However, the well developed lobe readily separates these species from forms like Mytilus edulis, and the terminal character of the beaks distinguishes the Paleozoic forms from typical volsellas of the modiola type. Details of form, such as the prominent umbonal ridge and ventral sinus, also differentiate the species in question from Recent Volsella and Mytilus. In many respects Promytilus is structurally and morphologically intermediate between Volsella and Mytilus. Inasmuch as the Paleozoic genus is somewhat more specialized than Volsella, it probably does not lie in the ancestry of that genus. More probably the reduction of the anterior lobe led to development of Mytilus, because Mytilus passes through an ontogenetic stage similar in form to Promytilus. Some of the Triassic and Jurassic shells commonly referred to Modiolus may belong to Promytilus.
Figure 8—Comparison of form in species of Promytilus.
PROMYTILUS SWALLOVI (McChesney)
Plate 1, figures 8a, b, 13
Myalina swallovi McChesney, 1860, Descriptions of new species of fossils from the Palaeozoic rocks of the Western States, p. 57; ——, 1865, Plates illustrating in part the new species of fossils from the Palaeozoic rocks of the Western States, and two new species noticed March 1860, published 1865, pl. 2, figs. 6a, b.
Valves "modioloid," with a well developed anterior lobe, and a pronounced anteroventral byssal sinus, which is symmetrical in respect to the anterior and ventral extremities; characterized especially by a relatively upright form, and large angle α, around 55°.
Dimensions of the plastotype are as follows: length, 24 mm, height, 18 mm; hinge length, 20 mm ±; convexity, 4 mm ±; greatest dimension, 26 mm; angle α, 60° ±.
Table 4—Species of Promytilus, Showing Age and Minimum, Average, and Maximum Dimensions1, in Millimeters
| Species | Age | Length | Height | Greatest Dimension |
Form Ratio |
Angle α |
|---|---|---|---|---|---|---|
| P. vetulus | Wolfcampian | 27 | 20 | 31 | 1.35 | 63.5° |
| 21 (6) | 16 (6) | 23 (6) | 1.34 (6) | 55.0° (6) | ||
| 16 | 12 | 17 | 1.33 | 49.5° | ||
| P. priscus | Virgilian | 32 | 27 | 37 | 1.44 | 72.0° |
| 15 (40) | 13 (40) | 18 (40) | 1.21 (39) | 55.5° (41) | ||
| 6 | 5 | 7 | 1.04 | 35.0° | ||
| P. annosus annosus | Upper Missourian |
29 | 26 | 32 | 1.67 | 62.5° |
| 21 (42) | 17 (42) | 24 (42) | 1.31 (42) | 50.5° (49) | ||
| 14 | 9 | 15 | 1.06 | 34.5° | ||
| P. annosus senex | Middle Missourian |
36 | 29 | 42 | 1.63 | 61.5° |
| 28 (124) | 21 (121) | 31 (124) | 1.33 (122) | 48.0° (128) | ||
| 19 | 14 | 20 | 1.07 | 36.0° | ||
| P. swallovi | Lower Missourian |
24 | 18 | 26 | 1.33 | 64.5° |
| 21 (4) | 16 (4) | 22 (3) | 1.32 (4) | 55.0° (4) | ||
| 13 | 10 | 15 | 1.30 | 47.5° | ||
| 1 Figures in parentheses indicate number of specimens used. Dimensions given in millimeters. | ||||||
Comparison—Table 4 shows that Promytilus swallovi differs from P. annosus and its varieties in having a greater value for the angle α, and in the smaller average size of the shell. The present species characteristically is somewhat more robust in shell size than is the ease in P. priscus. Although the angle α is approximately the same in the two species, specimens of P. swallovi are relatively longer (1.32) on the average than those of P. priscus (1.21). Inspection of average dimensions of specimens of the various species tabulated in table 4 indicates no significant difference between P. swallovi and P. vetulus. The two are readily distinguished by differences in form, however, especially the greater identation of the byssal sinus in P. swallovi.
Material—This is not a common species, and seemingly all but the original references are to other species of Promytilus. Only four of the specimens before me are referable to P. swallovi, and these include a sulphur cast of the original. The holotype was long ago destroyed, but artificial casts of it are preserved in the U.S. National Museum and Walker Museum, at Chicago. Only the exterior is visible in the shells at hand, but the form and peculiar shell structure of the Mytilidae is well shown. Commonly, the corrosion of a Promytilus shell produces the illusion of faint radial costellae by the unequal solution of the fine radially directed calcite prisms. The artificial cast of the holotype of P. swallovi shows faint radial markings, suggestive of radial prisms in the original.
Occurrence—As known at present, the species seems to be characteristic of the lower part of the Missourian series, but future collecting probably will reveal a somewhat longer stratigraphic range.
There is some uncertainty regarding the horizon from which the holotype was collected, the only information available being that it came from "12 miles northwest" of Richmond, Mo. The stratigraphic section at this locality ranges from uppermost Des Moines beds through the lower part of the Missourian series. Other hypotypes came from lower Missourian beds near Pleasant Hill, Mo. and the middle Missourian Winterset limestone, Dennis formation, Kansas City, Mo.
The shells occur in argillaceous and carbonaceous limestone, associated with brachiopods and mollusks, indicative of a marine environment. One specimen (pl. 1, figs. 8a, b) from Pleasant Hill, Mo. has the two valves in apposition. indicating relatively quite deposition and burial in situ.
PROMYTILUS ANNOSUS ANNOSUS Newell, n. sp., n. var.
Plate 1, figures 9, 10
Shell obliquely elongate, with a well developed anterior lobe and sulcus, of which the latter joins the anteroventral margin near the middle to produce a marked sinus; distinguished by an intermediate average value for the angle α, 50.5°, and an intermediate form ratio (length/height), 1.31.
Dimensions of the holotypes are: length, 17 mm; height, 16 mm; greatest dimension, 21 mm; angle α, 62.5°.
Comparison—The typical variety of the species Promytilus annosus is most similar to P. annosus senex, from which it differs in having slightly less robust shells, in being relatively shorter, and in having a larger angle between the umbonal ridge and the dorsal margin. Although the differences are well marked where a large suite of specimens of the two varieties are compared, it is probable that they could not be distinguished from each other if adequate collections were lacking. Comparison of the measurements of P. annosus annosus with other Late Paleozoic forms is indicated in table 4.
Material—The variety is based on 42 specimens, all of which were measured and studied. The shell material is relatively well preserved; and, although the inner ostracum seems to be recrystallized, the outer ostracum of both valves shows the peculiar radial prismatic structure of the Mytilidae in the majority of specimens. Hinge characters and musculature were not observed. The holotype (no. 7559) and paratypes are at the University of Kansas.
Occurrence—As represented in the collection before me, the variety Promytilus annosus annosus occurs in molluscan limestone facies of the upper Kansas City and Lansing groups (upper Missourian), especially Farley limestone, various localities in northeastern Kansas; Bonner Springs shale, shell coquina in the vicinity of Kansas river; Springhill limestone (Plattsburg), Bonner Springs, Ottawa, and Garnett. Kan.; undifferentiated Stanton limestone at the "Mound," Bartlesville, Okla., same, see. 32, T. 28 S., R. 15 E., Kansas; sandstone of the Rock Lake member, Stanton limestone, sec. 27, T. 11 S., R. 22 E., Kansas; South Bend limestone member, Stanton limestone, 2 miles south of Lansing, Kan., at Nine-Mile school (holotype).
The shells occur in shaly and sandy beds and in shell breccias, suggesting that the living animal preferred a littoral or sublittoral habitat, or at least lived in an environment where the shells could periodically be attached and broken up by waves and currents. All known specimens are separated valves, indicating disturbance of the shells after death.
PROMYTILUS ANNOSUS SENEX Newell, n. sp., n. var.
Plate 1, figures 11, 12; plate 2, figures 4a, b, 6
Myalina? swallovi Sayre, 1931, Kansas Geol. Survey, Bull. 17, p. 117, pl. 11, figs. 1, 1a. (Date of imprint, 1930).
Shells similar to the typical variety, but on the average more robust with greatest dimensions 31 mm, as compared with 24 mm for Promytilus annosus annosus, with a somewhat longer shell, and a smaller value for the angle α (48° as compared with 50.5°).
Dimensions of the holotype are: length, 29 mm; height, 23 mm; greatest dimension, 34; angle α, 53°.
Comparison—Comparative measurements are given in Table 4. The variety Promytilus annosus senex has a relatively slender form, due to the small average angle α. In respect to the size of the angle this variety has a smaller angle, 48° on the average, than any of the other Pennsylvanian and Permian species thus far discovered. Also, specimens of P. annosus senex are larger than other comparable material examined during the present study. Since absolute size of individuals in Volsella and Mytilus is commonly dependent on ecological conditions, this character in the present form may be without phylogenetic significance.
Material—The variety Promytilus annosus senex is based on more extensive collections than commonly are available for Paleozoic Mytilidae, 128 specimens having been used in framing the above description. The shell material, although somewhat recrystallized, is fairly well preserved, and the characters of form and of the hinge are well shown. In general, the outer surface of the shell is perfectly preserved and shows no appreciable wear. The radial prismatic structure of the outer octracum is evident in both valves in the majority of specimens.
The holotype (no. 7563) and many of the paratypes are at the University of Kansas. Other paratypes are deposited in the Walker Museum, University of Chicago; Peabody Museum, Yale University; and the paleontological collection, University of Wisconsin.
Occurrence—This variety characterizes the molluscan facies of the cross-laminated oolitic limestones of the Westerville and Drum formations (Kansas City group) of Kansas and Missouri and is represented by numerous specimens in every large collection of fossils from these rocks throughout Kansas and Missouri. Thus far, the variety is not well represented in nonoolitic facies in the Kansas City group. The holotype was collected from the Westerville oolite, at Kansas City, Mo. Other specimens of this variety are from the Graford and Brownwood formations (Missourian), Texas.
The valves are invariably dissociated, and their occurrence in strongly cross-laminated beds indicates burial in the zone of strong wave or current action. Few of the specimens are worn or were broken before burial, but many are coated with a calcareous deposit reminiscent of the algal. deposits that are commonly noted on shells of modern Volsella and Mytilus.
Figure 9—Frequency graphs showing comparison of form ratio (length/height) and angle α (obliquity) in three Pennsylvanian forms of Promytilus, n. gen. Promytilus priscus is geologically the youngest, P. annosus senex, the oldest. The number of specimens used in drawing the graphs was, respectively, 40, 42, and 124.
PROMYTILUS PRISCUS Newell, n. sp.
Plate 1, figures 1, 2, 5-7
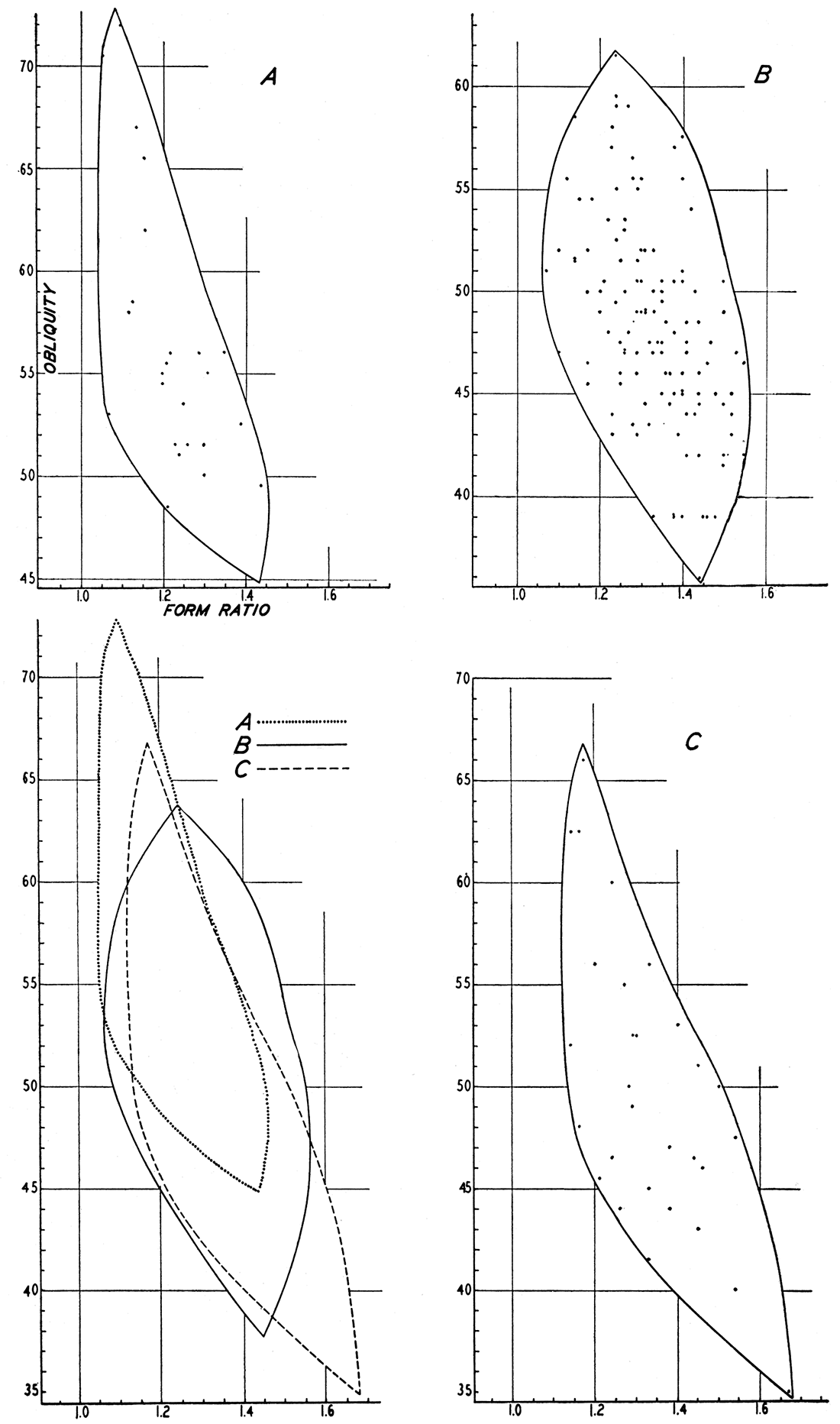
Form obliquely elongate, with well developed anterior lobe and sulcus, and otherwise similar to other Late Paleozoic forms; distinguished by the relatively small size of individuals, and particularly by the small form ratio (1.21) as compared with similar species (1.31-1.34).
Dimensions of the holotype are: length, 20 rom; height, 15 mm; greatest dimension, 25 mm; angle α, 46.0°.
Comparison—Comparative measurements of Promytilus priscus and other similar forms are given in table 4. The relatively great height (form ratio 1.21) as compared with similar species distinguishes the present form.
Figure 10—Scattergrams comparing shape in three forms of Promytilus. Obliquity is expressed in size of the angle α. A, P. annosus annosus; B, P. annosus senex; C, P. priscus.
Material—Forty-one specimens, all dissociated valves, were used in framing the above description. The majority of the specimens are well preserved, but a few, including the holotype, show signs of some surficial corrosion, suffered probably before fossilization. The characteristic structure of the outer ostracum of the Mytilidae is well shown by the radial striations of etched specimens. Hinge characters and interior not observed. The holotype (no. 7751) and several topoparatypes are at the University of Kansas. A few paratypes are in the paleontological collections at the University of Wisconsin.
Occurrence—The new species Promytilus priscus seems to be rather widely distributed in Virgilian rocks of the northern Mid-Continent area. In the collections before me specimens of the species are recognized from the Tarkio limestone (Wabaunsee), sec. 32, T. 10 S., R. 10 E., Kansas; Auburn shale (Wabaunsee), near Foraker, Okla.; Wakarusa limestone (Wabaunsee), near Thurman, Iowa and Osage county, Okla.; limestone above Brownville horizon (Wabaunsee?), near Thurman, Iowa; Lawrence shale (Douglas), Ft. Leavenworth, Kan.; Doniphan shale (Shawnee), Snyderville, Neb. and Greenwood county, Kan.; and the Dover limestone (Wabaunsee), NW cor. sec. 14, T. 13 S., R. 13 E., Kansas (includes holotype and several topoparatypes).
The specimens occur in association with other pelecypods and gastropods in arenaceous and argillaceous limestones. The fact that the valves are separated suggests burial within the zone of wave and current action. However, none of the material at hand is badly macerated, and few valves show any signs of wear prior to burial.
PROMYTILUS VETULUS Newell, n. sp.
Plate 1, figures 3, 4
This species is closely similar to Promytilus swallovi as regards size, proportions, and the value of the angle α. There is, however. a slightly more pronounced indentation of the anteroventral margin in P. swallovi, and in the present species the anterior part of the shell is somewhat more slender. Comparative measurements of P. vetulus are given in table 4.
Dimensions of the holotype are as follows: Length 27 mm; height, 20 mm; greatest dimension, 31 mm; angle α, 55°.
Material—This species is based on seven specimens from various horizons in the Big Blue (Wolfcampian) rocks of Kansas. The material is not adequate to permit generalizations regarding the normal environment under which the shells were buried. The matrix adherent to the specimens, however, is an argillaceous limestone and argillaceous dolomite, and in each case the valves are separate, suggesting some current action at the site of deposition. Some of the specimens are sufficiently well preserved to exhibit the shell structure of the Mytilidae, but internal characters are not shown.
The holotype (no. 7552) and paratypes are deposited at the University of Kansas.
Occurrence—Thus far the species has been recognized only in rocks of Wolfcampian age. The holotype was collected from the Admire group, 1.7 miles south of Americus, Kan. Para types came from the Admire group, just west of Neosho river, Kansas; Fort Riley limestone (Chase group), at the railroad quarry, NE sec. 36, T. 16 S., R. 7 E., Kansas; Florence limestone (Chase group), Grand Summit, Kan.
Figure 11—Form series in Volsellina and Promytilus. There is a tendency for younger Promytilus to become more upright in form.
Genus VOLSELLINA Newell, n. gen.
Genotype, Pleurophorus subellipticus Meek. Geologic range of genotype, Pennsylvanian.
Shell elongate, thin, with subterminal beaks, and a small, but distinct anterior lobe, which extends slightly ahead of the beaks; dorsal and ventral margins relatively straight, diverging posteriorly in the order of 20°, more or less; anterior sulcus and ventral sinus very obscure or lacking; umbonal ridge narrow, subangular, only slightly arcuate, with backward convexity, extending to the posterior end of ventral margin where the shell is slightly angular; convexity of valves rather low, posterior end rounded, laterally flattened, nearly spatulate; ligament, shell microstructure, as in other Mytilidae; endentulous.
Range—Pennsylvanian to Permian.
Remarks—Superficially, this group of shells recalls the heterodont genus Pleurophorus, but it is nearly certain that they are not closely related, differing markedly in hinge structures. From Volsella ("Modiolus") and similar Mytilidae it differs in the flattened valves, narrow costa-like umbonal ridge, and in the nearly straight ventral margin. Almost all of the shells examined by me are in the bivalved condition, denoting quiet water deposition, as contrasted with the Promytilus, and were found in argillaceous shales. One specimen of V. subelliptica from the Atoka formation is preserved as an internal mold in a fine, calcareous sandstone. Only one valve is represented.
In addition to the Pennsylvanian Volsellina subelliptica, this new genus is represented by V. transparens (Waagen), from the Upper Productus limestone (Cyclolobus zone of the Permian).
VOLSELLINA SUBELLIPTICA (Meek)
Plate 1, figures 14-16
Pleurophorus subellipticus Meek 1867, Am. Jour. Sci., vol. 44, p. 181.
Modiola? subelliptica Meek, 1872, U.S. Geol. Survey of Nebraska, p. 211, pl. 10, fig. 5.
Shell elongate, laterally flattened, lenticular, equivalve, with relatively straight dorsal and ventral margins which diverge posteriorly at about 20°; beaks low, broadly rounded, not extended generally above the hinge line; hinge margin about two thirds of the shell length, the shell being broadly spatulate and wedge-shaped in the posterior one-third; posteroventral margin angular where the straight ventral margin meets the rounded posterior margin; umbonal ridge narrow, subangular, extending nearly straight across the shell, or in a shallow arc, from the beaks to the posteroventral angle of the shell margin; the angle α is relatively small, 35° to 44°, increasing only slightly during growth, because of the small curvature of the umbonal ridge; form ratio about 2.0, and the length equal to the greatest dimension.
| Measurements of Volsellina subelliptica | ||||
|---|---|---|---|---|
| Length (mm) |
Height (mm) |
Thickness (mm) |
Angle α | |
| Holotype | 24.0 | 13.0 | 38° | |
| Hypotype 1 | 22.0 | 11.0 | 3.5 | 44° |
| Hypotype 2 | 34.0 | 17.0 | 6.0 | 44° |
Comparison—The only species with which I am acquainted that is closely comparable to this one is Volsellina transparens (Waagen), from the uppermost Permian of the Salt Range in India. From Waagen's figures it seems that the Permian species differs from V. subelliptica in having a relatively short hinge, extending only about one half the shell length. Also the Indian specimen is larger than the largest known American specimen. As in V. subelliptica, the Permian species has a narrow and well defined umbonal ridge, but there is a pronounced tendency in the latter for the ridge to become subdued and ill-defined toward the posterior extremity.
Material—This species has been cited from many horizons and localities; but in the absence of adequate illustrations, I am relatively certain of the identification in only two collections that have been available to me. Meek's type material came from the Willard shale (Wabaunsee), at Nebraska City, Neb. A poorly preserved internal mold was collected from the lower Atoka beds near Muskogee, Okla. Two hypotypes included in this study are from a shale in the Jacksboro limestone (lower Virgil), 4 1/2 miles east of Jacksboro, Tex. The holotype is at the U.S. National Museum and two hypotypes are at the University of Wisconsin.
Occurrence—Stratigraphic occurrences are indicated above. Undoubtedly this species has a relatively long range and will ultimately be recorded from many horizons in the Pennsylvanian. Probably bases for specific or varietal differentiation within these shells will be discovered when adequate material is available. Both the Texas and Nebraska occurrences are in fine argillaceous shale, and specimens with the valves in position indicate burial in relatively quiet waters as contrasted with the usual occurrence of Promytilus, in which the valves generally are separated and commonly broken, being imbedded in cross-bedded coquinoid limestones.
Genus LITHOPHAGA Bolten, 1798
Lithophaga J. F. Bolten, 1798, Mus. Boltenianum, p. 156.
Lithodomus Cuvier, 1817, Le Régne, vol. 2, p. 471.
Genotype, by monotypy, L. mytiloides Bolten = Mytilus lithophagus Gmelin. Geologic range of genotype, Recent.
Shell relatively small, smooth, elongate, subcylindrical, with terminal or subterminal beaks and a laterally compressed wedge-shaped posterior.
Geologic range—Carboniferous to Recent.
Remarks—The shell of Lithophaga is distinguished from Volsella and similar Mytilidae chiefly by its cylindrical form, being similar to other mytilids in other respects. Lithophaga is, however, highly specialized as to habit and habitat, modern species being confined to the shallow water littoral and sublittoral zones of limestone coasts in warm waters, where the living animal bores cylindrical holes in the limestone within which the animal spends its life. According to Pelseneer (1906, p. 252), Lithophaga
bores its hole by the aid of the acid secretion of glands situated in the anterodorsal and posterodorsal regions of the mantle.
Specimens of Lithophaga are quite rare in Paleozoic rocks, but several species have been described from the Lower and Upper Carboniferous and Permian in many parts of the world. There is nothing to indicate whether or not the Paleozoic lithophagas had acquired the rock boring habit. It is even possible that the close resemblance between these early forms and the Recent species is homeomorphic. However, no good reason has thus far been advanced for placing the Paleozoic species in a separate genus or subgenus.
LITHOPHAGA SUBELLIPTICA Sayre
Plate 4, figures 3-5
Lithophaga subelliptica Sayre, 1931, Kansas Geol. Survey, Bull. 17, p. 123, pl. 12, figs. 9, 9a. (Date of imprint, 1930).
This species is extremely rare, being represented in our collections by only three specimens that are at all instructive, and of these only the holotype, an immature individual, is complete. The holotype is clearly congeneric with many of the other Carboniferous species from Europe and Asia that have been referred to Lithophaga. It should be obvious, however, that the specified characters cannot be known from the scanty material at hand. Two topotypes before me are many times larger than the holotype, but they both are incomplete so that the details of form are not well shown.
All of the specimens are subcylindrical, relatively smooth, with subparallel ventral and dorsal margins; slight anterior lobe extends just perceptibly beyond the low, rounded and non-protuberant beaks; approximately two thirds of the shell length behind the front margin, the posterior edge slopes downward obliquely at a low angle to form a somewhat narrow and extended posterior extremity. The dimensions of the holotype are as follows: Length, 11.5 mm; height, 4.5 mm.
Occurrence—Westerville limestone (Kansas City group), Kansas City, Mo. (topotypes, nos. 7566 and 7567, Univ. Kansas) and Muncie, Kan. (holotype, no. 59, Univ. Kansas). The shells were found in cross-bedded oolite, indicative of deposition under conditions of strong current action.
Genus LITHODOMINA Waagen, 1881
Lithodomina Waagen, 1881, India Geol. Survey, Mem., Palaeontologia Indica, ser. 13, pp. 264-266.
Genotype, Lithodomina typa Waagen, 1881; range of genotype, uppermost Permian, India.
According to Waagen, the shells of Lithodomina
are more or less elongate, equivalve, very excentric, with subanterior beaks, little developed anterior part of the shell and very elongated posterior part, and with nearly parallel dorsal and ventral margins. The beaks are singularly flattened as in Lithodomus (Lithophaga). The hinge is edentulous. Along the hinge line extends a long, not very deep, furrow for the insertion of an internal ligament. There seem to be two muscular impressions, between which an entire pallial line seems to extend, but no distinct observation on this point is possible. The structure of the shell is very thin, devoid of every kind of sculpture and, if well preserved, shining smooth. Only with a strong magnifying lens, very fine distant striae of growth are observable.
From all these characters it appears that the shells belonging to Lithodomina are most nearly related to Lithodomus, the chief difference consisting in a more strongly developed anterior part of the shell, which is very likely coincident with a stronger development of the anterior adductor.
Geologic range of the genus, zone of Cyclolobus, Salt Range, India.
Remarks—There is sufficient ground for separating this group of shells from Lithophaga on form differences alone. Unfortunately, the characters of Lithodomina are not adequately known and Waagen admitted the possibility that these shells may not even belong to the Mytilacea. At the present time the genus is unknown outside the original occurrence in the uppermost beds of the Upper Productus limestone, in the Salt Range.
Prev Page--Shell Morphology || Next Page--Family MYALINIDAE
Kansas Geological Survey, Geology
Placed on web Dec. 18, 2017; originally published 1942.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/Vol10_2/04_system.html