![]()
Prev Page--Contents || Next Page--Shell Morphology
Introduction
Representatives of the pelecypod superfamily Mytilacea are among the most diverse, most abundant, and best preserved of Paleozoic pelecypods. Among these the myalinids are of particular interest because of the remarkable shell beds formed by their remains. At many horizons, particularly in the Pennsylvanian and Early Permian rocks of the Mid-Continent region, there are thin beds of limestone or shale, up to a foot or so in thickness, filled with the massive valves of these pelecypods. Scores of beds of this kind are traceable for tens of miles, and constitute excellent datum horizons for field mapping. Certain "Myalina beds," such as occur in the Meadow limestone (Lansing), Vinland shale (Douglas), Stull shale (Shawnee), Caneyville formation (Wabaunsee), and Moran formation, to mention only a few, are incredibly persistent, and represent old shell "banks" comparable with the extensive Mytilus and Ostrea beds of the present day seas. Unlike modern Mytilidae the Late Paleozoic Mytilidae were not markedly gregarious and, in general, appear to be much more sporadic than the myalinas. Little is known about early Paleozoic ancestors of the Mytilacea, sensu stricto, nor are the Devonian and Mississippian Myalinidae and Mytilidae adequately known. It is certain that the splendid material from the Pennsylvanian and Permian rocks herein described will shed much light on the morphology of the more poorly preserved older faunas. The Early Permian (Wolfcampian) Mytilacea are well represented, at least in American faunas, but the later Permian appears to be poor in representatives of the superfamily.
In view of the abundance of good material available, it is surprising that so little attention has been paid by systematists to the Late Paleozoic Mytilacea. Although Hall (1884-1885), DeKoninck (1885), Frech (1891), and Hind (1895, 1897) dealt with these shells in their pioneer studies of Paleozoic pelecypods, their observations did not lead to a satisfactory understanding of this important group of fossils. Indeed, because of a marked tendency for the mytiloids to develop along a few well defined trends, homeomorphy within the group is very common, and we find the earlier students of Paleozoic pelecypods classing some of the myalinids as pteriids ("aviculids") because of a striking similarity in form between various species of the two different stocks.
Greatest interest in Late Paleozoic Mytilacea comes from their hitherto unsuspected stratigraphic value as excellent guide fossils. Several separate phylogenetic groups are recognized, some of which display marked modifications in form as they are traced through the Late Paleozoic sequence. The relative abundance of myalinid shells, together with their limited stratigraphic range, makes these fossils second to no other group of fossil pelecypods known to me as good indices of geologic age. In this respect they are superior to many of the genera of associated brachiopods, in which evolutionary trends are generally not so evident.
As the need for greater refinement in stratigraphic paleontology increases, many paleontologists are turning to statistical methods as a partly objective approach to the evaluation of fossil species. Paleontologists will do well to heed Simpson's (Simpson, 1937, pp. 303-314; also Simpson and Rowe, 1939) plea for more satisfactory documentation of fossil evidence. It is axiomatic that no two animals are exactly alike, even though they may be spawned from the same parents. This inherent variability should be taken into account in describing a species. Many paleontologists select one, or at most a few, of their best specimens for the description and the duplicates are set aside. With Simpson, I believe that the characters of a species cannot be adequately known in absence of frequency graphs or some other form of statistical data. Without some knowledge of the extent of variability of a given species, its limits cannot be defined. Of course it would be absurd to argue that inadequate material should never be described, or even in some instances named, because wholly satisfactory collections are not common. The naming of a distinctive fossil may well be justified even though only one specimen is available. However, the usefulness of such a species in evolutionary paleontology is very limited, and the limitations of such a species should clearly be recognized. Naturally a thorough statistical analysis is impossible with rare or bulky materiaL There is no justification, however, for failure to employ quantitative data in the descriptions of new species of small gregarious animals, such as Foraminifera or ostracodes. In the present study I have tried to learn the limits of variations within the collections at my disposaL
Since paleontologists seldom can evaluate the ecologic factor in modifications of structure and form of fossil animals it seems important that the statistics of a "species" be based on a collection from a single horizon and locality, wherever practicable. The holotype and "topoparatypes" then, with a high degree of probability, are conspecific and were subject to the same impacts of environment. Other supplementary paratypes obviously are not so valuable in a detailed analysis. [Note: Although I deplore the ridiculously large number of proposed categories for type specimens, the term topoparatype has particular usefulness. A paratype which is also a topotype is obvlously more reliable than a paratype which comes from a locality and horizon different from the holotype.]
Summary of Results
Paleontologists are gradually coming to admit that different taxonomic groups among fossils do not have equal time value for purposes of dating and correlating rocks. The logical inference to be drawn from this conclusion is that correlations based on objective analysis of percentages of species common to two formations are not so reliable as are the correlations founded on genetic series of certain rapidly evolving groups. However, even among the more celebrated groups of guide fossils, such as the trilobites, graptolites, ammonites, and fusulinids, there are comparatively long ranging genera and species that have only a limited usefulness in correlations.
The pelecypods are no exception to the rule. Some families appear to have been relatively conservative and relatively unchanging through great spans of geologic time; whereas, others underwent conspicuous changes as traced through the rocks. In this latter category belong the myalinas and related mytiloids, herein considered, from the Pennsylvanian and Permian rocks.
It is probable that representatives of the Mytilacea are more abundant in the American Late Paleozoic than any other single group of pelecypods. In the Mid-Continent Pennsylvanian and Lower Permian they are locally so abundant as to make shell beds. A dozen or more of these shell beds are individually traceable for more than one hundred miles.
Down through the years it has been customary to refer almost all Carboniferous and Permian species of the Mytilacea to the genus Myalina. The results of the present investigation reveal that there are around a dozen separate phyletic lines represented in these shells. Eight genera and subgenera in two families are recognized among the shells commonly called Myalina by American paleontologists.
The two families are readily distinguished by peculiarities of the ligament area and by the shell microstructure. One family, the Mytilidae, seems to be the more primitive and the longest lived, ranging perhaps from Ordovician to Recent. The other, the Myalinidae, seemingly originated in the Devonian but did not survive the close of the Jurassic period.
Eleven genera and three subgenera from the Late Paleozoic are considered in detail. Of these, four genera and two subgenera are here defined for the first time. Early in the progress of this work it became evident that some of the Early and Medial Paleozoic genera of pelecypods may belong to the Mytilacea. Genera of the Ambonychiidae Miller (emend. Ulrich), the Modiolopsidae Fischer, and the Modiomorphidae Miller closely resemble some of the Late Paleozoic forms herein classed as Mytilacea. A thorough revision of the early genera, beyond the scope of this study, is needed before the ancestral Mytilacea can be defined satisfactorily.
Genera in the Late Paleozoic forms are differentiated partly by external form and internal structures, including details in the placement of the muscles. The musculature was essentially like that of modern Mytilus and Volsella.
Seven species and varieties, four of them new, are recognized among the Mytilidae. It happens that the species within this family are closely similar in form all the way from the Late Paleozoic to the Recent, so that discrimination of individual species requires detailed analysis. The application of some of the elementary principles of statistics has shown that there are useful guide species within this family. Two genera within the family are recognized as new.
Thirty-three species and varieties, 15 of them new, are recognized in the Myalinidae. They are distributed. among six genera and subgenera, of which four are new.
Taxonomic innovations are as follows:
| New genera, species, and varieties described in this paper. | ||
|---|---|---|
| Mytilidae | ||
| P. romytilus, n. gen. | ||
| P. annosus annosus, n. sp., n. var. | ||
| P. annosus senex, n. sp., n. var. | ||
| P. priscus, n. sp. | ||
| P. vetulus, n. sp. | ||
| Volsellina, n. gen. | ||
| Myalinidae | ||
| Myalina DeKoninck, emend. | ||
| M. wyomingensis var. thomasi, n. var. | ||
| M. petina, n. sp. | ||
| M. miopetina, n. sp. | ||
| M. pliopetina, n. sp. | ||
| M. glossoidea, n. sp. | ||
| M. arbala, n. sp. | ||
| M. lepta, n. sp. | ||
| M. copha, n. sp. | ||
| Myalinella, n. subgen. | ||
| Orthomyalina, n. subgen. | ||
| O. slocomi var. parvula, n. var. | ||
| Selenimyalina, n. gen. | ||
| S. quadrata, n. sp. | ||
| S. dotti, n. sp. | ||
| Septimyalina, n. gen. | ||
| S. burmai, n. sp. | ||
| S. orbiculata, n. sp. | ||
| S. scitula, n. sp. | ||
| S. quadrangularis, n. sp. | ||
The Myalinidae are much more highly specialized than the Mytilidae, and their ontogenies suggest that they were derived from the latter family. The myalinas constitute some of the best guide fossils among the pelecypods, and there are no other Paleozoic pelecypods that better illustrate consistent trends in evolution than do the members of this family. The progressive modifications in shape through the rock sequence are especially striking and are faithfully recapitulated in the ontogeny of the more specialized species in each phyletic line. The more conspicuous trends in form include changes in shell obliquity and the development of a rear wing. These trends are not peculiar to the Myalinidae but are noted also in the Mytilidae, Ambonychiidae, Pteriidae, and half a dozen more or less unrelated pelecypod families. The parallelism in development recalls homeomorphic trends in the ammonite suture, in the shell form of certain brachiopod stocks, graptolites, corals, and other groups of invertebrates. In the majority of species of Myalinidae, the shells become less oblique at the higher horizons; and the angle at the posterior end of the hinge becomes progressively smaller, until in the extreme cases a posterior wing, reminiscent of some of the Pteriidae, is formed.
At the outset of this work it was found that 32 specific and varietal names had been introduced for American Pennsylvanian and Permian mytiloids. They are indicated in the following list.
| Pennsylvanian and Permian mytiloids reported from North America |
|---|
| Naiadites carbonarius Dawson |
| N. ohioensis Morningstar |
| Lithophaga subelliptica Sayre |
| Modiola subelliptica Meek |
| M. wyomingensis Lea |
| M. minor Lea |
| Myalina swallovi McChesney |
| M. aviculoides Meek and Hayden |
| M. copei Whitfield |
| M. recurvirostris Meek and Worthen |
| M. acutirostris Newell and Burma |
| M. cuneiformis Gurley |
| M. meeki Dunbar |
| M. perattenuata Meek and Worthen |
| M. orthonota Mather |
| M. recurvirostris var. sinuosa Morningstar |
| M. ampla Meek and Hayden |
| M. slocomi Sayre |
| M. meliniformis Meek and Worthen |
| M. exasperata Beede |
| M. sinuata Branson |
| M. subquadrata Shumard |
| M. deltoidea Gabb |
| M. kansasensis Shumard |
| M. apachesi Marcou |
| M. pernaformis Cox |
| M. shannoni Beede |
| M. bialata Beede |
| M. recta Shumard |
| M. girtyi Beede |
| Mytilus permianus Swallow |
| M. concavus Swallow |
Of these I was able to secure type specimens for the first 20 species. Two of the species, Modiola minor and Myalina exasperata, are represented by such imperfect types that they can be identified in only a general way. Furthermore, the exact horizon and locality from which the type specimens came is unknown for either species. M. recurvirostris is placed in the synonymy of M. wyomingensis. Two additional species, for which the types are not available, M. subquadrata Shumard and M. sinuata Branson, are more or less securely established on topotype specimens. M. deltoidea Gabb is placed in the synonymy of M. subquadrata. The last nine names in the list should be suppressed because the type specimens are either lost or destroyed, the original horizon and locality are not precisely known, and the species were so inadequately described as to be unrecognizable.
The field relations suggest that the Late Paleozoic Mytilacea had habits and habitats similar to those of modern Mytilacea.
It can be demonstrated that the ligament system of such pelecypods as Perna and Arca are fundamentally different. It seems improper to use Dall's term multivincular to describe both types of ligament. Therefore, a new term, duplivincular, is herein proposed for the peculiar ligament of Arca, Myalinidae, Ambonychiidae, Pseudaviculopectinidae, and many of the Paleozoic genera of so-called Pteriidae.
Collections
Practically all of the great American collections of Late Paleozoic pelecypods have been made available for this investigation. The vast collections of the Kansas and Nebraska Geological Surveys, Yale Peabody Museum, and the Texas Bureau of Economic Geology, numbering some thousands of specimens, represent a large proportion of the fossiliferous horizons in the Mid-Continent region. Choice specimens were generously loaned by the Illinois Geological Survey, the U.S. National Museum, Walker Museum at the University of Chicago, and the University of Iowa. Small collections were donated or loaned to me by many individuals. In addition, some hundreds of the Pennsylvanian specimens were collected by me. In the aggregate, the material covered in this study is unquestionably the most extensive collection of Late Paleozoic Mytilacea ever assembled for study. My own field experience, however, leads me to believe that these collections do not faithfully indicate the actual distribution of Mytilacea in the Pennsylvanian and Permian rocks. The collections are very sparse and otherwise unsatisfactory from the early Pennsylvanian and from the later Permian rocks. The Mytilacea are so ubiquitous that I am convinced that every marine fauna in the American Pennsylvanian and Permian contains, at least locally, some of these shells. Therefore, the chief gaps in the record are attributed to incomplete collections, and, if one had the opportunity to examine all of the fossiliferous horizons not now represented in our collections, I am sure that the record of evolution in LatePaleozoic Mytilacea could be made singularly complete.
Distribution
Lithologic associations.-Only too seldom have paleontologists recorded the type of rock matrix from which their fossils were derived. The matrix is, in a limited way, indicative of the bottom ecology at the burial site. It is more or less obvious to anyone who has examined shell heaps along the sea shore that the shells may become buried under conditions quite unlike the life habitat of the animals in question. However, fossil bivalve shells may reveal transportation after death through the separation of the valves. The valves of dead Mytilacea become separated almost immediately because there are no interlocking devices capable of holding the valves in position after decomposition of the ligament. In this work I have tried to record the field occurrences of each species. Where the valves are found in apposition it is reasonably certain that the site of burial was the same as the life environment. Under this condition an examination of the rock matrix and associated fossils may furnish clues regarding the life environment of the Mytilacea. Those valves which are fragmentary and separated may have been transported by waves and currents for a considerable distance before burial. Careful consideration of the available evidence indicates that the Late Paleozoic Mytilacea favored the shallow-water, nearshore habitats like those preferred by the majority of modern Mytilacea.
Stratigraphic distribution—A single chart showing the stratigraphic range of different species of Mytilacea in each of the various provinces is not feasible. Therefore, in order to indicate the general sequence of stratigraphic divisions in the main geologic provinces, the following correlation tables and synthetic range chart have been compiled from various sources, representing the best-established correlations now available.
Table 1—Principal Divisions of the American Marine Permian Rocks
| West Texas | North Texas | Northern Mid-Continent |
Colorado Plateau |
Middle Rockies |
Russia | |
|---|---|---|---|---|---|---|
| Ochoan | Absent | Absent | Absent | Absent | Tartarian Kazanian Kungurian |
|
| Guadalupian | Capitan | Quartermaster Whitehorse |
Quartermaster Day Creek Whitehorse |
|||
| Word | Absent? | Absent? | Phosphoria | |||
| Leonardian | El Reno Clear Fork Lueders Clyde Belle Plains |
El Reno Enid Sumner |
Kaibab Toroweap Coconino Supai |
Absent? | Artinskian | |
| Wolfcampian | Admiral Putnam Moran Pueblo |
Chase Council Grove Admire |
Bird Springs | Upper Wells? | Sakmarian | |
Table 2—Principal Divisions of the American Pennsylvanian Rocks
| North Texas | Oklahoma | Northern Mid-Continent | Illinois | Allegheny Plateau |
||
|---|---|---|---|---|---|---|
| Ciscoan | Thrifty Graham |
Wabaunsee Shawnee Nelagoney |
Virgilian | Wabaunsee Shawnee Douglas |
Absent | Monongahela |
| Canyonian | Caddo Creek Brad Graford Whitt |
Ochelata | Missourian | Pedee Lansing Kansas City Bronson Bourbon |
Merom | Conemaugh |
| Skiatook | Embarass LaSalle Macoupin Shoal Creek Trivoli |
|||||
| Strawnian | Lone Camp | Holdenville Wewoka Wetumka |
Desmoinesian | Marmaton | Gimlet Sparland Brereton St. David Summum |
|
| Allegheny Pottsville |
||||||
| Millsap Lake | Calvin Senora Stuart Thurman Boggy Savanna McAlester Hartshorne |
Cherokee | Liverpool Greenbush Wiley Seahorne Delong Seville |
|||
| Lampasan | Smithwick Big Saline |
Atoka Barnett Hill |
Absent | Absent | Pope Creek Tarter |
|
| Morrowan | Marble Falls | Morrow | Babylon | |||
| Absent | Pushmataha | Wayside | ||||
Table 3—Known Stratigraphic Range of Late Paleozoic Mytilacea. (Extreme known range without regard to gaps in record or geographic distribution.)
| Stratigraphic Range | Guadalupian | Leonardian | Wolfcampian | Virgilian | Missourian | Desmoinesian | Lampasan | Morrowan | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Myalina (Myalinella) acutirostris Newell and Burma | * | * | ||||||||||||||||||||||||||||||
| Myalina (Myalina) wyomingensis thomasi, n. var. | * | * | ||||||||||||||||||||||||||||||
| Myalina (Myalina) sinuata Branson | * | * | ||||||||||||||||||||||||||||||
| Myalina (Myalina) aviculoides Meek and Hayden | * | * | ||||||||||||||||||||||||||||||
| Promytilus vetulus, n. sp. | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Myalina) copei Whitfield | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Myalina) pliopetina, n. sp. | * | * | * | * | ||||||||||||||||||||||||||||
| Septimyalina burmai, n. sp. | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Orthomyalina) subquadrata Shumard | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Myalina) petina, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Myalina (Myalina) miopetina, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Myalina (Orthomyalina) slocomi parvula, n. var. | * | * | ||||||||||||||||||||||||||||||
| Promytilus priscus, n. sp. | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Myalina) glossoidea, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Promytilus annosus annosus, n. var. | * | * | ||||||||||||||||||||||||||||||
| Selenimyalina quadrata, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Promytilus annosus senex, n. var. | * | * | ||||||||||||||||||||||||||||||
| Lithophaga subelliptica Sayre | * | * | ||||||||||||||||||||||||||||||
| Promytilus swallovi McChesney | * | * | ||||||||||||||||||||||||||||||
| Myalina (Myalina) arbala, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Septimyalina orbiculata, n. sp. | * | * | ||||||||||||||||||||||||||||||
| Selinimyalina meliniformis Meek and Worthen | ? | * | * | * | * | ? | ||||||||||||||||||||||||||
| Myalina (Orthomyalina) slocomi Sayre | * | * | * | * | * | * | * | * | ||||||||||||||||||||||||
| Myalina (Orthomyalina) ampla Meek and Hayden | * | * | * | * | * | ? | ||||||||||||||||||||||||||
| Myalina (Myalinella) meeki Dunbar | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | ? | ||||||||||||||||
| Septimyalina scitula, n. sp. | * | * | * | * | * | * | * | * | * | * | * | * | * | |||||||||||||||||||
| Selenimyalina dotti, n. sp. | * | |||||||||||||||||||||||||||||||
| Myalina (Myalina) wyomingensis Lea | * | * | * | * | * | * | * | * | * | * | * | * | * | * | ||||||||||||||||||
| Myalina (Myalina) lepta, n. sp. | * | |||||||||||||||||||||||||||||||
| Septimyalina quadrangularis, n. sp. | * | |||||||||||||||||||||||||||||||
| Myalina (Myalinella) cuneiformis, Gabb | * | |||||||||||||||||||||||||||||||
| Septimyalina sinuosa (Morningstar) | * | * | * | * | ||||||||||||||||||||||||||||
| Myalina (Myalina) copha, n. sp. | * | |||||||||||||||||||||||||||||||
| Volsellina subelliptica (Meek) | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | ||||||||||||||||
| Septimyalina perattenuata (Meek) | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | ||||||||||||||||
| Septimyalina orthonota (Mather) | * | * | * | * | ||||||||||||||||||||||||||||
| Naiadites ohioensis Morningstar | * | * | * | * | ||||||||||||||||||||||||||||
| Naiadites carbonarius Dawson | * | * | * | * | * | * | * | * | ||||||||||||||||||||||||
Acknowledgements
This study was accomplished with the aid of a financial grant from the Wisconsin Alumni Research Foundation, used principally to secure the assistance of Mr. Benjamin Burma and Mr. Frank Merchant. The importance of the aid from these two men can hardly be estimated because constant pressure from various duties would certainly otherwise have prevented me from completing the project for many more months. Mr. Burma prepared all of the photographs, under my direction, and compiled most of the statistical data for the descriptions of species. Mr. Thomas Roberts has critically read the manuscript and has offered valuable suggestions.
Especial thanks are due Dr. Raymond C. Moore, University of Kansas; Dr. G. E. Condra, Nebraska Geological Survey; Dr. Carl O. Dunbar, Yale University; Dr. Fred B. Plummer, University of Texas; Dr. J. Marvin Weller, Illinois Geological Survey; Dr. Carey Croneis, University of Chicago; Dr. A. K. Miller, University of Iowa; and Dr. G. Arthur Cooper, U.S. National Museum. These men have placed the great collections under their care at my disposal for purposes of this study. Nearly one hundred other individuals have been of great service in contributing select specimens for study, in granting access to type specimens, and otherwise materially aiding in the furtherance of the study. I have tried to credit these men, many of them foreign colleagues, at appropriate places in the text. If I have neglected to acknowledge specific aid, the oversight is unintentional.
Habits and Habitats
Mytilidae—Considerable is known about the bionomics of living Mytilidae. Species of Mytilus, particularly, have been intensively studied and a large amount of information has been summarized in an excellent monograph by White (1937), from which much of the following generalizations are taken.
Mytilus edulis Linnaeus, the genotype of Mytilus, is a cosmopolitan contemporary species of the littoral and shallow neritic zones. Specimens have been found in deeper waters but are smaller and less abundant than normal. The preferred habitat is from the high water mark to depths of only a few fathoms. Although there is a slight ventral gape of the valves for extrusion of the byssus between the closed margins, an extension of the periostracum around the edge, together with manipulation of the mantle margins within the shell, make it possible for the animal to seal itself hermetically within the valves. These "mussels" can live out of water for a long time if the shell is tightly closed, because a quantity of sea water is retained in the pallial cavity. In fact, the animal can live longer if the exterior of the shell remains dry, because periodic wetting of the shell causes the animal to open the valves and the contained sea water then will escape. Mytilus under observation has lived normally for over a month in sterile water, and individuals have been found alive after twenty-four days in the hot sun. The adductor muscles still show movements after forty days of anaerobic conditions. It is seen, then, that Mytilus edulis is well suited for life under the difficult conditions of the littoral zone.
This species is tolerant of great variations in salinity and occurs in salt, brackish, and in nearly fresh water. The animal can respire normally in water varying from the average salinity up to an admixture of 30 per cent fresh water or with an increase of salinity resulting from the addition of from 5 to 10 per cent of water that has been concentrated to a salinity of 1 per cent. However, addition of fresh water to 60 per cent, or an increase in salinity to 35 per cent of added water, which has been concentrated to 7 per cent salinity, kills the animal quickly.
Variations in salinity, particularly freshening of the water, produce changes in the animal that are shown in shell characters. Decrease below normal salinity of the water produces a corresponding stunting of growth. Studies in the Kaiser Wilhelm canal and in the Baltic, where salinity decreases from west to east, show that there is a progressive decrease in the size of the mussels. The shells seem also to become thinner and more fragile in brackish water.
Adults are always attached by the byssus, and the living animals are consequently not found on sandy shores unless there is some adequate supporting structure present, such as the shells of other mollusks. Empty shells of Mytilus very commonly are concentrated on sandy beaches by storm waves, so that the environment of burial is not necessarily the same as the life environment. The mussel is markedly gregarious, so that, where present at all, it usually is abundant; and the surface of the ground at low tide commonly is completely covered with them, no free space being left for other sessile benthos. It has been computed that the density of individuals in a community may be as great as 16,000 to the square foot, but of course this number is exceptional.
Although normally attached by the byssus, the animal is capable of locomotion, sometimes propelling itself by the foot in a way similar to that of the snail. Young individuals crawl with the anterior end foremost, but adults reverse the direction of movement. Mussels have been observed to climb up a perpendicular glass by means of the suctorial tip of the foot, lifting themselves on the foot and then making a byssal thread fast. A second thread is attached above the first, after which the latter may be broken off by a sudden jerk of the foot. When the animal desires more or less permanent anchorage several threads are attached to the substratum, radiating out in various directions. A considerable amount of limited movement is accomplished by means of the byssal muscles, which can, by their contraction and relaxation, vary the tension on various byssal threads. In this way jerky movements can be produced within the limits of the byssus. This is the way in which mussels remain covered for a longer period by receding water, being able to lower themselves somewhat into the water by the byssus.
Experiments indicate that Mytilus edulis thrives best in the presence of medium sunlight and a temperature range between 10° and 20° C., but they can retain ordinary activity at temperatures between 0° and 26° C.
The animal is microphagous, filtering detrital material from the sea water. It has been estimated that ten gallons of water pass through the gills in twenty-four hours. Investigations indicate that live phytoplankton does not represent nearly as important food for pelecypods as detritus.
The annelid worm, Polydora ciliata, commonly infests oysters and mussels, living in burrows drilled into the shell substance of the pelecypod. The worm enters through a hole the size of a pin and makes a U-shaped burrow in the shell. It does not attack the soft tissues of the animal, but sometimes it does damage to the pelecypod by boring completely through the shell, causing the growth of pearly excrescences on the interior of the shell next to the mantle. Paleozoic Mytilacea were infested by similar boring annelids (pl. 8, figs. 4, 5).
The meroplanktonic larvae of pelecypods permit relatively wide distribution of many species. The larvae of the majority of forms, however, are so sensitive to the physical and biological environment and enjoy such a short free-swimming period that there are innumerable barriers to migration during the larval stage.
Recently it has been shown by Nelson (1928) that Mytilus edulis is exceptional, perhaps unique, among pelecypods in adopting the pelagic habit during the early dissoconch stage. Shells up to 941 microns in length have been found floating. According to Nelson (1928, pp. 183-185)
the presence of well developed dissoconchs floating freely in the water at once raises the question of the means by which this is effected in the absence of the swimming organ or velum of the prodissoconch. When brought to the laboratory for examination ... a large bubble was found to occupy the posterior part of the pallial cavity, its buoyancy causing the young bivalve to hang suspended in the water umbones downward, with the posteroventral margin of the valves turned upward.
Some young mussels maintain themselves at the surface through means other than the gas secretion employed by larvae at a depth. It was observed that once in contact with a solid object, such as the wall of the chamber, a rock, or a fragment of sea weed, the mussels exhibited a marked negative geotropism and climbed straight upward until the surface was reached. Here the distal one-third to one-tenth of the foot was extended to the surface film, and with a quick contraction of the foot, aided apparently by contraction also of the pedal retractor muscles, the ventral margins of the mantle were brought into contact with the surface film. While lying with the entire ventral margin of the body in contact with the surface film the byssus gland in a few seconds secreted onto the surface film a small holdfast similar to that which is laid down on rock or piling for the attachment of each byssus thread. A thread 1 to 2 mm long serves to support the young mussel from this float and with foot withdrawn it may hang suspended indefinitely. . . . The 'float' is not a buoyant structure, since when pushed beneath the surface it rapidly sinks. It maintains its position in the surface film, supporting mussels up to 4 mm in length, solely through surface tension.
Other individuals were observed in which the young mussel supports itself solely by the end of the foot in the surface film, recalling the familiar habit of pond snails of hanging from the entire foot spread out in the surface film. Another, but less common, mode of suspension from the surface consists in extending the tops of the tentacles of the incurrent siphon into the surface film and hanging from there.
Of the known genera of marine pelecypods, Mytilus edulis and Teredo navalis alone are circumpolar in their distribution over the shores of the northern hemisphere. General adaptability to changing conditions and the power to resist adverse surroundings, together with relatively low spawning temperatures, have aided these two forms in attaining their present wide distribution. Transportation through attachment to vessels or to other floating wood has likewise aided in their dispersal, being for Teredo the only means by which any great distance could be covered. In the case of Mytilus edulis, however, the ability to bridge the period of metamorphosis while remaining pelagic must have been an important factor in securing the wide dispersal which this mollusk now enjoys; as well as a great aid in bringing to a suitable place of attachment a fair proportion of the larvae produced each season. The largest of the pelagic dissoconchs found in Frenchman Bay was fully a month old, during which time it must have been transported over long distances by the tide.
Figure 1—Mytilus edulis, early dissoconch (X90), showing one method of suspension from surface film of water by byssal thread. [Image scaled and magnification adjusted for web presentation.]
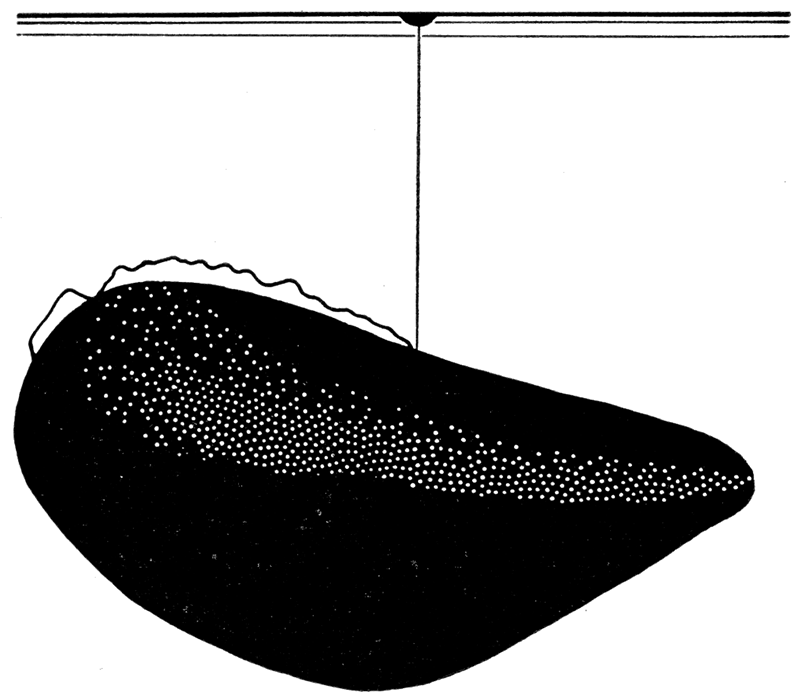
The genus Lithophaga Bolten (Lithodomus Cuvier) employs the byssal mode of fixation during the early ontogeny; but at maturity the animal, which is found only in regions of calcareous rocks, bores holes in the rocks by the aid of the acid secretion of glands situated in the anterodorsal and posterodorsal regions of the mantle (Pelseneer, 1906, p. 252). There are a number of citations in literature to Late Paleozoic species of Lithophaga. However, I know of no examples of borings in limestone produced by Late Paleozoic pelecypods. A modioloid rock-borer, Coralidomus concentricus, from the Cincinnatian rocks of Ohio, was described by Whitfield; but it is not certain that this form belongs to the Mytilacea.
Myalinidae—Like the Mytilidae, the myalinas were gregarious; and, judging from the musculature, they were byssate for at least a part of their life span. The more specialized myalinas, however, unlike Mytilidae, are very thick-shelled and during life may have had very little need for a byssus. Only relatively strong waves and currents would be able to move some of these shells, and it is doubtful that these animals were sufficiently powerful to drag their shells about after maturity was reached.
The shells of myalinas are found in various sediments ranging from bituminous or argillaceous shales to sandstones. In all of these rocks, shells with both valves in apposition are common, indicating little or no transportation after death. A few species are restricted to, or are more common in, certain kinds of rock. Naiadites seems to have preferred the brackish or fresh-water habitat, whereas Myalinella meeki is found associated with various biota that suggest quite a range in salinity.
It is doubtful that the larger myalinas were very active, because their heavy shells commonly are encrusted with lamellar deposits of calcium carbonate, suggestive of algal deposits, and the shells commonly are perforated by the U-shaped burrows of annelid worms. Furthermore, the living animal was in some instances very small compared with the size of its shell.
Judging from the commonness of bivalve specimens of Myalinidae having the two valves in apposition, it seems probable that the myalinas preferred relatively quiet water. Separated valves are found, but these seldom show evidences of markedly turbulent water. Since deposits of the littoral zone rarely are preserved in the fossil record, except perhaps in the paralic swamps, there is little to indicate whether or not many of the myalinas favored areas that frequently were exposed to the atmosphere. The Naiadites, however, so characteristic of the carbonaceous shales of the early Pennsylvanian in eastern North America and Europe, probably led an existence much like some Mytilus edulis, advancing far inland at the estuaries, attaching themselves to salt marsh vegetation. The faunal associates of Naiadites suggest that species of this genus did not normally favor the more typically marine environment.
The more strikingly gregarious myalinas, such as Orthomyalina, Myalina, and Septimyalina, in some instances occur crowded in a stratum to the near exclusion of other fossils. Perhaps the crowding among the myalinas made some localities practically uninhabitable for other shelled benthos.
The relatively pure limestones in the Pennsylvanian and Permian do not commonly contain representatives of the Myalinidae or Mytilidae, and these seldom are found in direct association with fusulinids. The supposition is that the pure limestones and the majority of other fusulinid-bearing rocks were deposited at depths too great, or distances from the shore too great, to be favored by the Mytilacea. Students of the Pennsylvanian apparently are agreed that the bulk of the deposits in the Mid-Continent region were deposited in water less than 50 fathoms in depth; therefore, it is highly probable that the myalinas and mytiloids preferred even shallower water of the near-shore environment.
There is much to indicate that some of the myalinas were among the first immigrants into the Mid-Continent region, during the frequent incursions of the sea, and among the last marine invertebrates to be destroyed by withdrawal of the sea. It has been shown (Moore, 1936, pp. 20-38) that the Pennsylvanian rocks are characterized by a great number of cyclic repetitions of marine and nonmarine rocks, indicative of numerous minor oscillations of the sea. Very commonly, the myalinas are most numerous shortly below and just above plant-bearing shales and coal beds, in deposits which presumably were laid down closer to the shore than some of the fusulinid-bearing, dominantly calcareous beds further removed, stratigraphically, from the nonmarine beds. Experience shows that members of the Mytilacea are common only in beds 0.2, 0.7, and 0.8 of the "ideal" cyclothem (Moore, 1936, pp. 24-25).
Considering all available evidence, it seems probable that many of the Paleozoic Mytilacea lived under conditions similar to those favored by modern Mytilus and Volsella, viz., the shallow, turbid waters of the shore zone, tolerating an unusually great amount of variation in the salinity of the water.
Evolutionary Trends
Hardly any group of fossil organisms illustrates parallel trends in evolution any more strikingly than some of the pelecypods. Progressive characters, i.e., characters in which the direction of evolution is in some degree inevitable and, therefore, likely to be repeated in parallel lines along a number of separate lines of descent, are well known in graptolites, corals, cephalopods, brachiopods, foraminifers, and mammals, and can be demonstrated in many other animal groups. Homeomorphy is so common in certain tribes as to suggest that there is a marked "preference" for certain kinds of variation. Certainly the fossil record is replete with illustrations of evolution that seem to be unidirectional as regards specific structures. Some progressive characters, such as increase in maximum size of individuals, and increase in specialization, whether or not obviously adaptive, have affected all sorts of animals during their history. There are a few kinds of progressive modifications that have affected so many tribes of pelecypods as to be reasonably regarded as standard evolutionary trends. Some of these trends are to be anticipated in many of the pelecypod stocks. The classification of pelecypods most generally employed by modern zoologists is Pelseneer's, which was based fundamentally on gill structure. The majority of students of fossil pelecypods, and a few of the zoologists now regard gill characters as belonging to the category of progressive characters. Thus Pelseneer's groups are merely grades of gill development, not groups of common ancestry. Douville has developed a classification based on the idea of adaptive radiation, a concept which has proved so successful in the taxonomy of the vertebrates. Parallel development due to similar adaptation is too well understood to require further attention here. It should be noted, however, that there are very many examples of parallelism, among the invertebrates, that can as readily be ascribed to a tendency for directed variation as to similarity in adaptation.
In addition to gill modifications and changes in the construction of the ligament described later, there are trends in form and shell structure that have affected many unrelated stocks of pelecypods. Those trends that are thus far recognized in the various tribes of Mytilacea are discussed below.
Size—Progressive increase in size of adults is a common trend in all kinds of animals, although it is by no means certain that this trend applies to all genera or families of animals. In the Late Paleozoic Mytilacea, increase in size is admirably exemplified by species of Myalina s. s. The oldest representatives of the genus, that I have seen, occur near the middle of the Mississippian rocks (St. Louis limestone, Viséan limestone). The largest of these specimens measure less than 70 mm in length. Specimens of Myalina from the Pennsylvanian are commonly somewhat larger, and Permian individuals having a length of 120 mm are common. Throughout this range, normally small adults of separate species are also found. Of course, progressive increase in size in any race of fossil animals is not easy to demonstrate without fairly complete collections. Ecologic variation in size should also be taken fully into account. The progressive increase in size of individuals in the collections under consideration is so marked that it can reasonably be attributed to evolution.
Obliquity—Compared with the hinge axis, the body of the shell in all primitive Pectinacea, Pteriacea, and Mytilacea, as well as certain other pelecypods, is markedly oblique. That is, a line of symmetry drawn through the umbo forms an acute angle with the hinge. This condition I have called prosocline. During the evolution of several of the families, as shown by the fossil sequence, as well as ontogenies, there is a tendency for the shell to assume a more upright form so that the axis of the shell body is more or less normal to the hinge (acline). The more specialized species show a continuation of this trend to a condition in which the ventral part of the shell is carried forward and the mature part of the shell assumes a backward obliquity (opisthocline). This trend is well shown in the Myalinidae and in some of the aviculopectens (Newell, 1938, p. 17). The obliquity is best expressed in angular degrees (angle α) as measured between the umbonal ridge at a mature stage and the dorsal margin. In the more specialized species the umbonal ridge is markedly arcuate, with a forward concavity, and the ontogenetic increase in the angle is very striking.
Posterodorsal extremity—The tendency to develop a rear projection of the shell at the hinge margin is very marked in Pteriacea and Pectinacea and is known to be a mark of specialization. Primitive species, as shown by both the ontogenies and the fossil record, invariably lack the posterior auricle. Two genera of the Myalinidae (Myalina, Septimyalina) independently developed rear auricles to a marked degree, giving them a pterioid aspect. In primitive species, the rear margin of the shell intersects the dorsal margin at an obtuse and somewhat indefinite angle (angle β). Progressive reduction in this angle is accompanied by increasing prominence of a posterodorsal salient. When the angle becomes acute a well defined auricle is formed. Although posterior auricles are not known in all of the genera of Paleozoic Mytilacea, the majority of them show a distinct tendency for progressive reduction in the value of angle β.
Anterior lobe—Primitive Mytilacea have distinctly a "modioloid" expression. That is, in addition to possessing the general proportions of "Modiolus" (Volsella), they are characterized by an anterodorsal salient, called the anterior lobe, in front of and below the beaks. Reduction of this lobe, with a tendency thereby for the beaks to become progressively more terminal, is distinctly a mark of specialization. Mytilus passes through a Volsella stage in its ontogeny and was, presumably, derived from the latter through reduction in the anterior lobe. Among the Myalinidae, the genus Myalina shows this trend quite strikingly. Primitive myalinas, such as M. wyomingensis, are quite lobate, almost modioloid, whereas specialized forms, such as M. copei and the species of Orthomyalina, have a very inconspicuous anterior lobe.
Loss of symmetry—The least specialized among living pelecypods show a high order of bilateral symmetry (equivalve). Adaptation to a pleurothetic existence has, apparently, been responsible for loss of the primitive symmetry in a number of tribes of pelecypods, such as the Ostracea, Pteriacea, Pectinacea, etc. The majority of Mytilacea are essentially bilaterally symmetrical and show no preference for one side over the other when they assume an orientation other than the vertical one. Under crowded conditions, living Mytilus seems to tolerate any orientation, even commonly suspending themselves by the byssus, in an inverted position, beneath overhanging ledges of rock. Primitive Myalinidae look very much like Mytilus, being distinguished chiefly by differences in shell composition and microstructure, which are readily observed in all but poorly preserved material. However, even the most primitive known Myalinas are slightly asymmetrical, having a slightly less convex right valve than left valve. In the more specialized and geologically younger forms the difference in convexity between the two valves is quite marked; and, in some instances, the valves become markedly discordant, that is, the right valve fits slightly within the margin of the left valve. Accompanying this trend, the anterior lobe of the right valve becomes obsolescent more rapidly than that of the left valve; so that there are several species of Myalinidae in which there is little or no distinguishable lobation of the right valve, while a small lobe is still apparent in the left valve. In all of these shells that show marked asymmetry of the two valves, there is a tendency for the beak of the left valve to be especially prominent, so that it overhangs the beak of the opposite valve.
There are no living analogues of the Myalinidae; so it cannot be proved, conclusively, that these animals normally lie on the right valve when at rest. However, a clue may be found in the primitive Pectinacea, which certainly lie with the right valve undermost (Newell, 1938, pp. 18-20). In the pectens, as in the myalinas, there is a very marked tendency for the prismatic structure to be obscure or lost in the left valve, while the primitive prismatic condition remains unmodified in the right valve. I have supposed that the relative conservatism of right valves of Pectinacea (idem, pp. 20-21) is due to the greater stability of the environmental influence, lying, as they often do, with the right valve partly buried in the mud. Also, it may be stressed that right valves of species of Myalinidae are less varied than left valves, so that there are many instances in which the right valves of two distinct species can scarcely be distinguished from each other. By analogy with the pectens, we may suppose that the peculiarities of the right valve in the myalinas were brought about by the assumption of a life position in which the right valve was undermost.
Figure 2—Time value of form trends in some Paleozoic genera and subgenera of Mytilacea. The more striking form stages in the phylogeny are superposed in composite diagrams. The known stratigraphic range of each stage is indicated in the bar scale below each diagram. The three spaces for the Mississippian are: Kinderhookian, Osage-Meramecian, Chesterian; four spaces for the Pennsylvanian are: pre-Desmoinesian, Desmoinesian, Missourian, and Virgilian; four spaces for the Permian represent: Wolfcampian, Leonardian, Guadalupian, and Ochoan. In a few instances a given form stage is represented by more than one species.
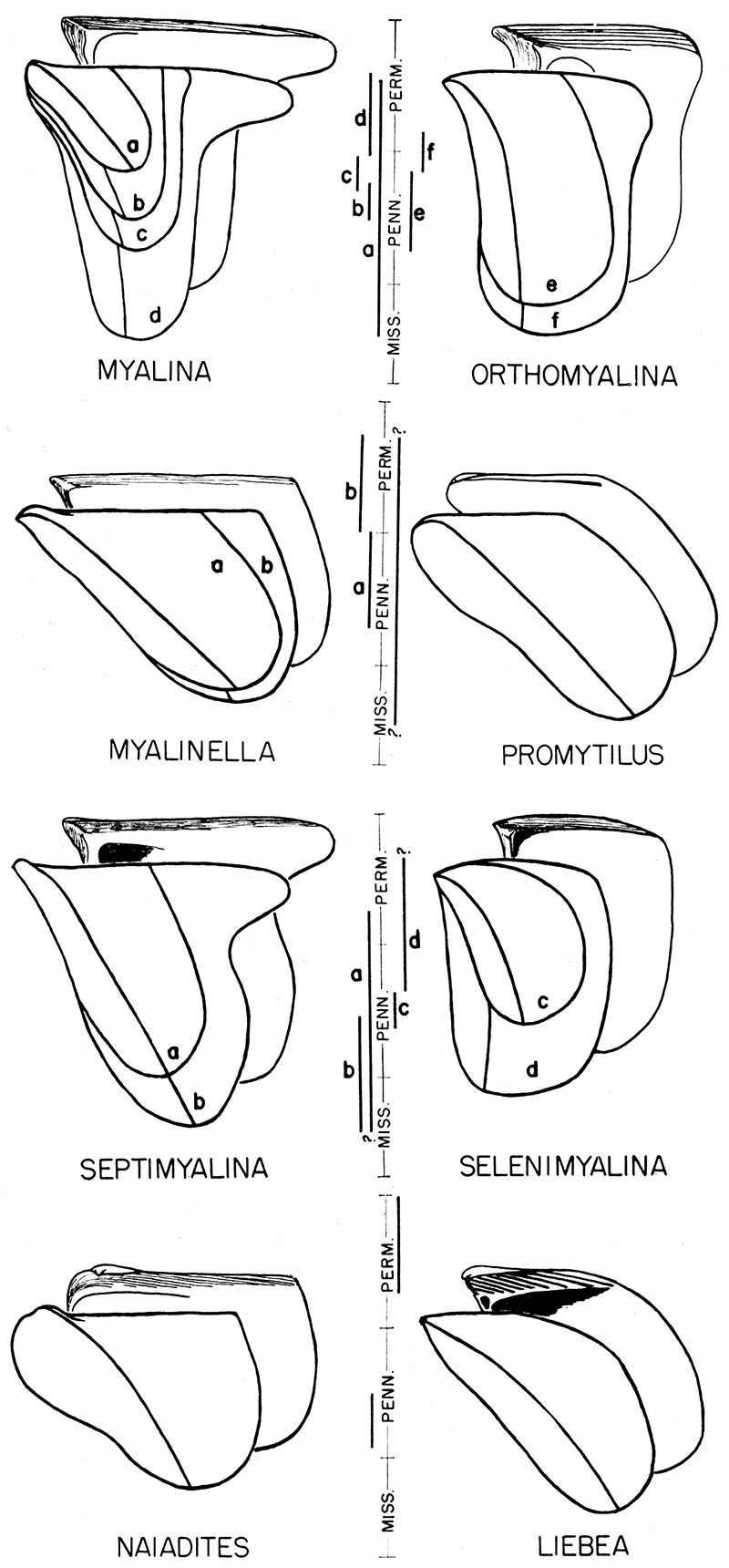
Orientation of muscles—Cursory examination of figure 6 will reveal that the posterior muscle system in several genera of the Mytilacea is differently placed in respect to the rear extremity of the hinge axis. In all of these shells there is a tendency for the anterior and posterior adductors to assume a position in line with the greatest linear dimension of the shell. For instance, the greatest dimension of the shell in a primitive Myalina, such as M. goldfussiana (fig. 6E), is measured between the beaks and the posterior margin of the shell. In any mytiloid shell having this general form the posterior muscle system is placed high in the shell not far below the terminus of the hinge. On the other hand, in markedly acline shells, such as those shown in figure 6B, C, G, the greatest dimension of the shell is measured from the ventral margin to the dorsal margin of the shell; and the adductors lie in a line that is nearly dorsoventral in orientation. It is obvious that there is a migration of the posterior muscles toward the ventral part of the shell as the shell itself gradually changes from the prosocline to the acline or opisthocline grade of obliquity. Concurrently with the migration of the posterior muscles there is indicated also a tendency for the anterior byssal retractor insertions to migrate in a posteroventral direction. These changes in the placement of the muscles apparently are adaptive in the sense that they tend to maintain mechanical efficiency during marked changes in shell form.
Prev Page--Contents || Next Page--Shell Morphology
Kansas Geological Survey, Geology
Placed on web Dec. 18, 2017; originally published 1942.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/Vol10_2/02_intro.html