![]()
Prev Page--Petrology || Next Page--Summary
Depositional Environment and Paleoecology
Regional Setting and Stratigraphic Framework of Greenhorn Deposition
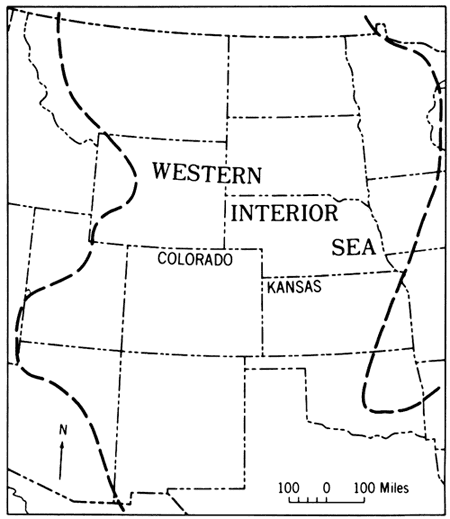
Late Cretaceous marine rocks of Kansas represent erosionally truncated deposits laid down in the eastern part of a broad seaway that by Cenomanian time extended continuously from eastern Mexico to Arctic Canada (Cobban and Reeside, 1952, p. 1022). The subsiding trough that admitted these waters has been called the Rocky Mountain geosyncline by many authors (e.g. Gilluly, 1963, p. 146; Armstrong, 1968, p. 432). Throughout the U.S. Western Interior region, subsidence was greatest along the western edge of this structural depression. Locally, as in southwestern Wyoming and central Utah the Upper Cretaceous deposits alone reach thicknesses between 18,000 and 20,000 feet (Reeside, 1944). In addition, Late Cretaceous sedimentary deposits become generally coarser grained toward the western margin of the trough (Reeside, 1957, p. 508) where, in a complexly intertongued relationship, fine-grained offshore marine shales give way to nearshore and marginal marine sandstones and these in turn give way westwardly to nonmarine deposits of coastal swamps and piedmont areas (see Pike, 1947; Spieker, 1949; Weimer, 1970, p. 273). Distribution of lithofacies delineates clearly a major western source area for the trough-filling detritus. This detritus was derived from tectonic highlands, the Sevier orogenic belt (Fig. 21) of Armstrong (1968, p. 435), that were raised along the site of the former Cordilleran geosyncline (Gilluly, 1963, p. 146; Armstrong, 1968, p. 432; King, 1969, p. 70). As a part of the stable craton, the area to the east of the seaway was low and flat (Reeside, 1957, p. 509) and for most of Late Cretaceous time was not an important source area for sediment contributed to the seaway (Weimer, 1970, p. 273).
Figure 21--Block diagram depicting a portion of the Western Interior Sea during deposition of the Greenhorn Limestone, and nature of bordering land areas.

In many parts of the Western Interior region, Late Cretaceous rocks manifest marked cyclicity; nowhere is this cyclicity marked more clearly than along the western edge of the former seaway where dark-colored shales and light-colored sandstones are intimately intertongued. Pike (1947, p. 15) has reviewed the two major hypotheses that have been put forth to account for such intertonguing, namely 1) repeated vertical crustal oscillation within the seaway during deposition, and 2) alternation of transgression caused by downward movements of the crust and regression resulting from subsequent detrital infilling of the basin margin. The latter hypothesis was favored by Pike (1947) and is accepted by the author as the most reasonable explanation of recorded sedimentary features. Details of cyclic sedimentation in the Four Corners area and in central and eastern Utah have been elaborated by Pike (1947) and Spieker (1949), respectively, and Weimer (1960) has documented on a regional basis four major cycles of sedimentation that can be recognized through much of the Cretaceous System as developed in the Rocky Mountains and Great Plains areas. The cycles are most readily deciphered along the western side of the Late Cretaceous trough where transgressions are reflected in westward-directed tongues of dark-colored shale and regressions are marked by eastward-directed tongues of sandstone. The youngest of these regressive sandstone bodies corresponds to the Fox Hills Sandstone of the classic Western Interior section, and before erosion of the Great Plains during Tertiary time, may have extended across parts of Kansas. The other regressive sandstone bodies did not extend as far east as Kansas, so in that state the pattern of cyclicity is different than in western Colorado and Utah. In the lower part of the Upper Cretaceous section in Kansas, the writer (liattin, 1964) recognized and described one complete cycle of sedimentation corresponding to the first cycle of Weimer (1960). The remainder of the Kansas section represents only part of a second and very large-scale cycle corresponding to the second, third, and probably part of the fourth cycles of Weimer (1960).
The first of the Kansas cyclothems embraces marine strata from the upper part of the Dakota Formation to the top of the Codell Member, Carlile Shale. The terminal units represent the beginning and end stages of transgression and regression, respectively. Maximum transgression (Fig. 22) is represented by relatively pure pelagic carbonates of the Jetmore and Pfeifer Members of the Greenhorn; for this reason the sequence was named Greenhorn cyclothem by Hattin (1962, p. 124). Initial deposits of the Greenhorn cyclothem are older in Colorado, where uppermost Dakota strata are of Late Albian age (Waagé, 1959), than in Kansas where marginal marine strata at the top of the Dakota are of Cenomanian age (Hattin, 1967, p. 588). The Dakota-Graneros contact is therefore diachronous, ascending chronologically in an eastward direction. Evidence has been presented above and in a previous work (Hattin, 1968) that the base of the Lincoln Member of the Greenhorn is also diachronous, climbing the section from southwest to northeast in western and central Kansas. The base of the sequence of close-spaced, resistant, chalky limestone beds of the Bridge Creek and Jetmore Members also is diachronous in an eastwardly ascending sense. The purest carbonate beds in the Greenhorn section are in the Jetmore and Pfeifer Members; the overlying Fairport Member, Carlile Shale, is lithologically similar to the Hartland and Lincoln Members and represents the initial phase of regression in the Kansas area of chalk deposition.
Figure 22--Paleogeographic map showing extent of Western Interior Sea at time (Early Turonian) of maximum transgression. Adapted from Reeside (1957, fig. 8).
The Greenhorn Limestone of Kansas represents accumulation of carbonate- dominated sediments far from the major, western source area of terrigenous detritus (Fig. 21). The writer (Hattin, 1971, p. 426) has determined that terrigenous detritus content of the Hartland and Jetmore Members increases westward, apparently reflecting influence of western sources. This pattern suggests that eastern source areas had little influence on Greenhorn sedimentation either by reason of small contribution from low-lying land areas of the central craton, or because of great distance from the eastern shoreline, or both.
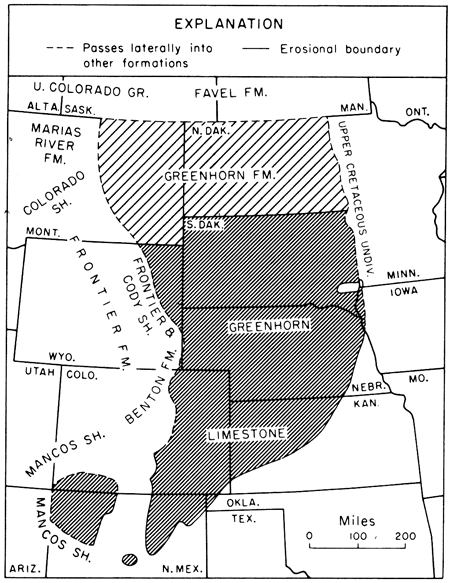
Greenhorn carbonates can be traced southwestward from Kansas across Colorado and northern New Mexico to the San Juan basin where they pass into shales and sandstones of the Mancos Shale (Dane and Bachman, 1957; Dane, 1960, p. 65, 66). Westward the Greenhorn thins gradually towards South Park, Colorado where a 40-foot-thick calcareous section containing as few as one bed of limestone was assigned to the Benton Formation by Stark and others (1949, p. 52). Beyond that area the Greenhorn interval is represented in westernmost Colorado and eastern Utah by white-weathering calcareous shale of the Mancos (Katich, 1956, p. 118). In northeastern Colorado and southeastern Wyoming the Greenhorn becomes less calcareous and is lost in a section called Benton Shale by McCrae (1956, chart); west of this area, the same interval is included in the Frontier Formation in southern Wyoming (Cobban and Reeside, 1952, chart 10b) and in northwestern Colorado (O'Boyle, 1955, p. 39). Northwestward from Kansas the Greenhorn persists as a carbonate unit as far as the Black Hills, but westward therefrom the carbonate units are replaced by shales and sandstones of the Frontier Formation and Cody Shale (Cobban and Reeside, 1952a, p. 1954-1955). Northwestward from the Black Hills the Greenhorn thins appreciably, becomes less calcareous and passes into calcareous shales and sandstones of the Colorado Shale in central Montana (Cobban, 1951, p. 2183-2185). In most of the eastern Montana subsurface, the Greenhorn equivalent is known as Greenhorn Formation (Billings Geol. Soc., 1969, p. 9). The Greenhorn extends northward from Kansas across much of Nebraska and South Dakota, and is recognized also throughout most of North Dakota by Hansen (1955, p. 29) who described the units as consisting in the subsurface of "dark gray calcareous soft shale with thin beds of very shaly limestone." In the latter state the strata apparently represent a broad transition zone wherein the Greenhorn grades progressively to less chalky and limy, and therefore less typical, lithology. North of the Canadian border, the Greenhorn is not recognized as a formal unit, but its position is recognized in the Favel Formation (second speckled shale unit of petroleum geologists) in Manitoba and Saskatchewan (Wickenden, 1945, p. 33). Regional distribution of the Greenhorn is depicted in Figure 23.
Figure 23--Map showing regional distribution of the Greenhorn Limestone and nomenclature of laterally contiguous units. Close-ruling indicates area where formal name is Greenhorn Limestone. Wide-ruling indicates area where formal name is Greenhorn Formation.
Sources of Sediment
The principal component of most Greenhorn rocks is calcium carbonate, nearly all calcite, which occurs in many forms. Examination of samples in the field and of thin sections and scanning electron micrographs (Hattin, 1971; Hattin and Darko, 1971) has served to identify virtually all carbonate components. Shaly chalk, the most abundant variety of rock in the Kansas Greenhorn, consists largely of coccoliths. These minute fossils are the skeletal remains of coccolithophores, members of the Chrysophyceae (golden-brown algae), modern examples of which are mostly floaters in the open ocean. When coccolithophores die, the remains sink to the bottom where the tiny skeletal plates accumulate as part of the sea floor ooze. Alternatively, coccolithophores are eaten and their skeletons reach the sea floor as fecal pellets or in animal carcasses (Bathurst, 1971, p. 267). Coccolithophores must have been exceedingly abundant in Cretaceous seas because the widely distributed chalk deposits of that system are generally rich in coccoliths (Hatch, Rastall, and Black, 1938, p. 167).
Also abundant in Greenhorn rocks are tests of calcareous foraminifera; these occur in vast numbers in many calcarenites and are conspicuous in most samples of shaly chalk and chalky limestone. The vast majority of these foraminifera were planktonic forms (Eicher and Worstell, 1970a), especially including members of the Heterohelicidae and Rotaliporidae. Like coccoliths, the foraminifers sank upon death to the sea floor where the skeletal remains became part of the accumulating pile of carbonate sediment.
In a few calcarenites of the Lincoln Member and in certain pelletoidal limestones of the Pfeifer Member, large numbers of minute spherical structures are concentrated in very thin laminae. These structures are believed to be related to the so-called calcispheres of the Paleozoic. I believe that Greenhorn structures of this kind may have been derived from some form of planktonic algae. Rupp (1966, p. 186) stated the case for algal origin of calcispheres but concluded that they are of benthonic origin. Such structures have been reported previously from the Lower Cretaceous of Texas (Thomas, 1931) and from the Cretaceous of Mexico (Bonet, 1956; Bishop, 1970) but have not been reported previously from the Western Interior region. Both of the authors regarded the Mexican calcispheres as being of planktonic origin.
Shaly chalk, many chalky limestones, and a few calcarenites contain spherical, or where compressed ellipsoidal, calcareous pellets composed of micrite. These structures are common also in chalks of the Fairport Member, Carlile Shale. The writer (Hattin, 1962, p. 106) concluded that these are fecal pellets of a coccolithophore-eating organism. The same interpretation is accorded similar pellets in the Greenhorn. Scanning electron micrographs of such pellets show them to be composed of coccoliths and ultramicroscopic blocky crystals of calcite that look like broken coccolith elements.
The remaining carbonate components are of benthonic or diagenetic origin. Inoceramid valves, valve fragments, and isolated prisms are chief among these constituents of Greenhorn carbonate rocks. The commonly fragmented condition of inoceramid valves, and almost ubiquitous occurrence of isolated elements of the prismatic layer are probably owing largely to pre-burial oxidation of an organic matrix of the kind that occurs within both nacreous and prismatic layers of living bivalves (Wilbur, 1964). Such oxidation would result in ready disintegration of valves, a process capable of producing immense quantities of skeletal debris. Apparently Tarr (1925, p. 259) was first to offer this explanation for Inoceramus debris in Cretaceous rocks. Presence in Greenhorn rocks of teeth of the shell-crushing shark Ptychodus suggest that these predators exacted a toll among the larger benthonic invertebrates. However, teeth of these sharks are sparse everywhere but at the base of the Lincoln Member, and even the predation of sharks or other carnivores could not account for the enormous numbers of prisms that were separated from parent valves. Additional skeletal remains, usually in the form of whole or little broken valves of oysters, anomiids, rare entoliids, cirripeds, and other benthonic organisms make up most of the remainder of primary carbonate material in the Greenhorn Limestone of Kansas.
Additional sources of calcium carbonate must be sought to account for the void-filling sparry calcite of biosparites and biosparrudites and for calcite that played an important role in conversion of originally carbonate muds into the microsparite or pseudosparite that comprises most chalky limestone beds. Circulating interstitial seawater furnished some carbonate for cementation of biosparites and for initial cementation of soft, highly porous bioturbated muds. A second major source for such carbonate was early diagenetic dissolution of aragonitic skeletal material. In biosparites, pressure solution phenomena suggest a third source of carbonate cement.
Silt and finer-sized terrigenous detritus comprises the bulk of insoluble residues in most Greenhorn samples. For the Hartland, Jetmore, and Bridge Creek Members details of residue analysis have been furnished in a paper by the writer (Hattin, 1971, p. 427). In these members percentages of insoluble residue range from 1.6 to 23.3 percent in chalky limestone and 5.8 to 78.4 percent in shaly chalk. The one sample of shaly chalk having less than 9.5 percent residue contains many lenses of skeletal limestone. The small number of shaly chalk samples having greater than 50 percent insoluble residue are adjacent to bentonite seams and are bentonitic. In general, less than 10 percent of any residue consists of diagenetic or epigenetic minerals. In these members the volume of terrigenous detritus increases westward suggesting greater influence in that direction of a western source area. Detailed study of residues was made of the Lincoln and Pfeifer Members only at Locality 3; the percentages of residue for chalky limestones and shaly chalk of these members fell within the ranges stated above. Lincoln residue percentages are comparable with those of the Hartland Member and reflect relatively high rates of terrigenous detrital influx while transgression was in progress. Pfeifer residue percentages compare more closely with those from the Jetmore Member and reflect the smaller amount of terrigenous detritus that reached central Kansas during the stage of maximum transgression when the shores were most remote from the area (Fig. 22). During the Early or Middle Cenomanian, central Kansas was the scene of fluviatile and deltaic sedimentation (Plummer and Romary, 1942, p. 342; Hattin, 1967, p. 587) for which the principal transport direction was toward the southwest (Franks and others, 1959, p. 237). By the time of later Greenhorn deposition, eastward transgression of the sea had pushed the shoreline far to the east or northeast of the present line of outcrop, so that the influence of cratonic source areas to the northeast were probably of less importance volumetrically than were the orogenic source lands to the west.
Arenaceous terrigenous detritus, consisting of silt and very fine to fine-grained sand, dominantly quartz, but including small quantities of feldspar, is common at the base of the Lincoln Member. Coarser detritus of this kind is associated with high-energy rudaceous skeletal limestones. Across much of central Kansas, basal Lincoln strata rest sharply on Graneros Shale along a disconformity that manifests clear evidence of erosional truncation (Hattin, 1965a, p. 11; 1968). Occurrence of relatively coarse terrigenous detritus only in basal beds of the Greenhorn suggests reworking from the Graneros Shale.
Except in the Lincoln Member, bentonite seams in the Greenhorn of Kansas can be traced for long distances along the outcrop (Hattin, 1971). These bentonites are composed predominantly of montmorillonite as determined by X-ray diffraction techniques. These seams represent devitrified volcanic ash from sources in the Sevier orogenic belt (Fig. 21). In some sections bentonite occurs also as pebbles, apparently reworked from the Graneros Shale, mostly in rudaceous biosparites lying at the base of the Lincoln Member.
Skeletal grains of vertebrate origin, especially including shark teeth, teleost scales and teleost bone fragments are virtually ubiquitous in Greenhorn carbonate rocks but range widely in abundance. Such grains tend to be concentrated in the biosparites and biosparrudites and are especially abundant in the basal part of the Lincoln Member. However, scales, bones, or bone fragments were recorded in shaly chalk units at every section examined in detail and these rocks contain scattered shark teeth and sparse, semi-articulated teleost skeletons as well. Vertebrate remains are least common in chalky limestone. Except for the likelihood that vertebrate remains have been concentrated in the basal part of the Greenhorn by reworking of the Graneros in areas of unconformable contact, the fish that yielded these skeletal remains were inhabitants of the Western Interior Sea during deposition of Greenhorn sediments. Also of probable vertebrate origin are rounded, cylindrical, dark brown phosphatic pebbles less than one centimeter in diameter. Most of those observed were in basal skeletal limestones of the Lincoln; rare pebbles of this kind have been recorded higher in the Greenhorn. These pebbles are probably coprolites.
The final primary constituent of Greenhorn rocks consists of organic matter. Many thin sections of shaly chalk contain very fine wispy grains of deep-amber to black material that is oriented parallel to laminations. Samples of weathered and nonweathered shaly chalk were analyzed for organic carbon. The nonweathered rock, which is of olive gray to olive black color, contains greater percentages of organic carbon than the pale yellow and orange-colored weathered rocks. Furthermore, all nonweathered shaly chalk units analyzed have significantly greater quantities of organic carbon than do adjacent chalky limestone beds. Percentages of organic carbon in shaly chalk and chalky limestone units in the Hartland, Jetmore and Bridge Creek Members have been reported previously by the writer (Hattin, 1971, p. 427). The preservation of greater amounts of organic carbon in shaly chalk was interpreted as the result of reducing conditions in the interstitial environment of those sediments. The same interpretation is advanced here for shaly chalk beds of the Lincoln and Pfeifer Members. In analyzed samples of nonweathered shaly chalk from all members the percentage of organic carbon ranges from 0.37 to 6.3. In a total of 20 samples, only three contained less than 2.3 percent organic carbon, and all three of those samples are from a unit so diluted by bentonite that the rock is less than 50 percent carbonate. In contrast, 10 analyzed samples of little-weathered chalky limestone contain 0.1 to 0.5 percent organic carbon, averaging only 0.28 percent.
The organic matter in Greenhorn chalky rocks may be in part finely divided particles of carbon derived from vegetable matter in terrigenous source areas. However, the relatively high percentages of organic carbon (up to 6.3 percent in one Lincoln sample), and lack of obvious carbonaceous specks and woody fragments in most Greenhorn rocks, suggests that the carbon has been derived largely from organic matter of marine origin, i.e. from decay of coccolithophores, foraminifera, fish, Inoceramus, etc. on the sea floor.
Sequence of Depositional Events
During early Late Cretaceous time the Western Interior sea spread gradually eastward across the central Great Plains region. This transgression is manifested clearly in the succession of lithofacies embraced in the stratigraphic interval extending from the upper part of the Dakota Formation to the top of the Greenhorn Limestone. Within the Dakota fluviatile sandstones and mudstones give way upward to estuarine and other marginal marine sediments lying in the upper few feet of the formation (Plummer and Romary, 1942, p. 345; Hattin, 1964, p. 207; 1967, p. 582; Siemers, 1971, p. 30). The formation is overlain conformably by the Graneros Shale which consists in its lower part of noncalcareous sandy and silty shales and thin, discontinuous sandstones. These beds grade upward into less sandy, locally calcareous shales containing thin, discontinuous beds of calcareous, shelly sandstone and oyster-rich skeletal limestone (Hattin, 1965a). The stratigraphic sequence of lithologic, macrofaunal, and microfaunal changes suggests convincingly that as Graneros contains proportionately more planktonic, as was progressively farther from the edge of the still-advancing eastern shoreline, and that ultimately the sea in that area attained normal or nearly normal salinity (Hattin, 1965, p. 50) ' Towards its top the Graneros contains proportionally more planktonic, as contrasted to benthonic, foraminifera and the shales are locally calcareous to chalky; these lithologic and faunal changes reflect the onset of environmental conditions that produced the Greenhorn carbonates. The shift from dominance of terrigenous detrital deposition (Graneros) to carbonate deposition (Greenhorn) is a manifestation of increased distance from shore, such that accumulation of dominantly pelagic carbonate sediment was volumetrically greater than contributions from the terrigeneous detrital dispersal system.
In a few places, especially including Locality 12 (Kearny County) and Locality 6 (Washington County), the upward change from noncalcareous silty clay shales of the Graneros to chalky Greenhorn strata is transitional through several feet of conformable strata. However, at all localities from Ford County (Loc. 8) to at least as far north as Lincoln County, the Greenhorn rests on the Graneros with sharp discontinuity (Hattin, 1968) where noncalcareous or only weakly calcareous shales are overlain by well-washed skeletal limestones (biosparites and biosparrudites) of the basal Lincoln Member. Even in places of lithologic transition between the two formations the lower part of the Lincoln contains many beds and lenses of skeletal limestone. These limestones reflect high-energy conditions at the sediment-water interface, with considerable sea-floor scour across much of central Kansas. Reworking of the Graneros is evident in quartz- dominant sand and silt, bentonite pebbles, and certain macroinvertebrate remains contained in basal Lincoln skeletal limestones. Dominantly chalky strata above these limestones are firm evidence against regression having occurred at this time. Concentration of skeletal limestone is characteristic of the basal Lincoln across much of the Great Plains region and the explanation of these limestones requires a regional rather than a local mechanism. In an essay on epeiric clear water sedimentation, Irwin (1965, p. 450-454) discussed a model that explains development of skeletal limestones in a high-energy, offshore sea-floor zone of wave impingement and dissipation in a belt tens of miles wide. The effects of wave dissipitation in offshore zones of epeiric seas have been elaborated also by Shaw (1964, Chap. 6, 7). Both Irwin and Shaw were concerned with essentially autochthonous sediment models which, as applied to the Dakota-Graneros-Greenhorn example, must be modified so as to accommodate terrigenous detritus as the dominant kind of sediment landward to the high-energy zone. The basal Lincoln skeletal limestones may be viewed as the initial result of eastward or northeastward sweep across Kansas of a relatively wide, high-energy, offshore zone of wave impingement on the bottom during the transgressional phase of the Greenhorn depositional cycle. A broad, nearer-to-shore, mostly lower zone characterized by deposits of terrigenous mud (Graneros) was at first scoured, deeply in places, and then buried beneath deposits of skeletal debris. The time elapsed during passage of this high-energy zone across western and central Kansas is manifest in the diachroneity of the Graneros-Greenhorn contact (Hattin, 1968).
Impure carbonate mud, later weakly consolidated into shaly chalk, was the dominant sediment above the base of the Lincoln. This represents deposition of largely pelagic sediment in a lower energy, far offshore portion of the eastern shelf region of the Western Interior Sea. Sporadic surficial sediment reworking, and consequent winnowing of the fines, concentrated in thin lenses and lensing beds the skeletal debris that comprises biosparites in the middle and upper parts of the Lincoln Member. As transgression continued, the production of skeletal sands and silts waned, first in western Kansas, then at the southern end of the central Kansas region and finally in the northeastern part of the outcrop. Lincoln deposition was punctuated by ash falls, now bentonite seams, which attest to crustal unrest beyond the western edge of the seaway.
Shaly chalk, chalk, and chalky limestone of the Hartland Member reflect continued deposition of largely pelagic sediments in a zone still farther offshore than for Lincoln sedimentation, and in water too deep for the bottom to have been affected significantly by surface waves. This corresponds to zone X in the epeiric sea model of Irwin (1965, p. 450). At least in one place, however, a large lens of biosparite was produced where a scour channel was filled by a current-washed deposit of foraminifer tests (Fig. 9, C). From time to time the influx of terrigenous detritus waned, and layers of purer carbonate accumulated (Hattin, 1971). Where least well developed these are now beds of micritic chalk; where best developed these layers formed beds of microsparitic chalky limestone. Nearly all of these beds were extensively bioturbated.
During deposition of the Jetmore Member, and the stratally equivalent part of the Bridge Creek, impure carbonate mud deposition continued to predominate. These beds have been consolidated into shaly chalk. As during deposition of the Hartland and lower part of the Bridge Creek, whenever terrigenous detrital influx was reduced significantly, a bed of relatively pure carbonate mud accumulated. During deposition of the lower submember, and equivalent part of the Bridge Creek, 9 such beds were formed. All of these beds were homogenized by a burrowing endobenthos. In the interval between deposition of JT-1 and JT-2, currents swept the sea floor across most of the present outcrop and produced a widespread thin to very thin lensing bed (Fig. 11,D) or group of very thin lenses of foraminifer-dominated biosparite. Widespread development of this unit, together with unusually uniform stratigraphy in this part of the section, including regionally developed time-parallel limestone beds (Hattin, 1971) and thin bentonite seams, bespeak a nearly featureless sea floor of exceptionally low gradient.
In the upper foot or two of the lower submember, shaly chalk beds are crowded with valves, fragments, and small lenses of inoceramid remains that served as substrate for clusters of Pycnodonte. Better circulation, occasioned by general renewal of current impingement on the sea floor, generated these shell-rich beds. Abundance of inoceramid remains persists into the overlying submember, which includes the three hard beds, JT-10 to JT-12. The valves are typically disarticulated, but breakage was limited. These limestones contain burrow structures, but these are progressively fewer upward in the section. Like the subjacent limestone beds, the hard beds resulted from periods of reduced terrigenous detrital influx. Early diagenetic loss of skeletal aragonite from the myriads of inoceramids is offered as explanation for the superior hardness of these limestone beds. The very large numbers of inoceramid valves may have served to reduce burrowing activity in these beds.
In the third submember of the Jetmore, shaly chalk-forming mud continued as the dominant sediment type. The abundance of Mytiloides was reduced sharply and oysters once again were scarce. Bottom conditions favored a limited epifauna, but conditions below the sediment-water interface virtually excluded burrowers. From this part of the section upward chalky limestone is commonly concretionary, a fact that may relate to the greater purity of carbonates in shaly chalk beds of the Jetmore and Pfeifer Members. Diagenetic segregation of calcium carbonate resulted in development of oblate spheroidal concretions and non-burrowed thin beds of chalky limestone that are common above JT-12.
The Shellrock limestone bed represents widespread, prolonged reduction of terrigenous detrital influx, coupled with conditions favorable for proliferation of Mytiloides (Fig. 13,D) and extensive burrowing in the lower part of the bed. The upper part of the bed was formed by diagenetic segregation of calcium carbonate as evident in the mammilated and commonly concretionary (Fig. 13,C) nature. Except for portions containing crowds of Mytiloides and a thalassinoid type of burrower, the sparse benthos suggests that bottom waters were poorly circulated during deposition of the Shellrock bed.
Pfeifer deposition, like that of underlying members, was dominated by accumulation of impure pelagic muds that later became shaly chalk. These beds contain less impurity than those in the Lincoln and Hartland Members. The lower two thirds of the Pfeifer, from the top of JT-13 to PF-2 contains much inoceramid debris and many small lenses of biosparite, as well as an epifauna of somewhat greater density and diversity than most beds of the Jetmore. Improved circulation of bottom waters fostered this fauna. Gentle currents broke up fragile inoceramid valves and concentrated the skeletal debris as small lenses. This activity was less pronounced during deposition of an equivalent part of the Bridge Creek in western Kansas. Decreased detrital influx may have been responsible for widespread chalky limestone beds like PF-1, as well as for the less obviously continuous, irregular, shell-rich beds below PF-2 (sugar sand). In all these beds, as well as chalky limestones in the upper part of the Jetmore, density of allochemical grains is much greater, on the average, than for chalky limestones of the Hartland and lower Jetmore (Table 3). This suggests exceptionally slow rates of terrigenous detrital sedimentation and apparently also a slower rate of fine carbonate mud deposition. The high proportion of fecal pellets in upper Jetmore and Pfeifer chalky limestones (Table 4) corresponds with the appearance of rocks having higher densities of allochems. One may speculate that for these beds greater-than-usual volumes of coccoliths or coccolith debris reached the sea floor in fecal pellets.
The irregular, discontinuous beds of shell-rich chalky limestone in the lower one-half to two-thirds of the Pfeifer is indication that primary concentration of carbonate was expanded by diagenetic segregation of CaCO3. Hemispheroidal concretions attached to these beds, and oblate spheroidal concretions amongst the beds lends force to this conclusion.
During deposition of Upper Pfeifer sediments, from just below the sugar sand to the Fencepost limestone bed, the sea floor was once again quiescent. Currents that produced lower Pfeifer skeletal lenses had waned so that production of these lenses ceased. Foraminifera, calcispheres, and fecal pellets occasionally fell to the sea floor in sufficient numbers to form grainstone laminae, but for the most part shaly chalk-producing muds were deposited. Within the sediments of this quiet sea floor, large, oblate spheroidal concretions developed beneath, or sometimes enclosing, the decaying remains of various organisms.
The terminal event of Greenhorn deposition was deposition of the Fencepost limestone bed. This bed, like many others in the Pfeifer, has high allochem density and abundant fecal pellets. Very slow influx of terrigenous detritus, coupled with very high fecal pellet production, produced yet another widespread bed of rock having a relatively low volume of matrix. The depositional regime that produced the upper few feet of the Greenhorn persisted with no change during the early part of Carlile sedimentation.
Physical Aspects of the Environment
General Statemtent
Much has been written about the depositional environment and paleoecology of Cretaceous chalk, especially the English and northwest European chalk. Until very recently the American chalk in general has received little study in this regard and the Greenhorn Limestone has been almost wholly ignored. The past few years have witnessed much-increased interest in this unit, with numerous authors making contributions to the better understanding of the conditions under which the formation was laid down. Still, no previous author has attempted embracive environmental synthesis of the Kansas Greenhorn. In the following sections pertinent environmental parameters are examined from the standpoint of organic and inorganic evidence. Paleoecology of important macroinvertebrate groups is given separate attention.
Depth of Deposition
No environmental parameter of chalk deposition has been discussed at so great length and by so many authors as depth of deposition. For over a century debate has waxed and waned as whether the chalk is an ancient analog of deep sea oozes or the product of accumulation in a much shallower depositional regime. Space permits review of only a few of the major papers and the conclusions therein.
The abundance, or apparent abundance, of planktonic foraminifera in some chalk deposits led many early authors to conclude in favor of oceanic depths for deposition of the British and European chalk deposits (e.g. Huxley, 1858; Fuchs, 1883; Hume, 1894). At this time only Cayeux (1897, p. 527) argued forcefully against the deep-water interpretation, stating that the maximum depth of Chalk deposition in the Paris Basin was less than 150 fathoms (275 m). An intermediate viewpoint on depth of the British Chalk is found in the classic monograph of Jukes-Browne (1903, 1904). On the basis principally of foraminifera, supported by evidence from other fossil groups, be concluded that the Lower Chalk represents deposition at depths of 400 to 500 fathoms (732 to 915 m) (1903, p. 358), the Middle Chalk at depths at most times exceeding 500 fathoms (915 m) (1903, p. 557) and probably reaching a maximum of approximately 650 fathoms (1185 m), and the Upper Chalk at depths up to 700 fathoms (1281 m) (1904, p. 377). The evidence from foraminifera, including depth determinations by several authors, has been reviewed extensively by Earland (1939) who fairly well demolished the notion that chalk is to be compared with deep sea deposits. He concluded (ibid., p. 20) that foraminifera and other fossils of the British Chalk suggest deposition at depths to 50 fathoms (92 m) or less (for impure chalk near the base of the Chalk) to a maximum of 300 fathoms (549 m). Earland reviewed the work of Jukes-Browne (1903, 1904), pointing out many errors in the interpretation of foraminifera.
Sorby (1861., p. 197) was aware that foraminifera may be comparatively rare in chalk, and that coccoliths are the chief constituent of the chalk matrix and on the basis of coccolith abundance he implied that the Chalk is a Cretaceous analog of modern deep Atlantic muds. Quite recently, however, Hay and others (1967, p. 431) mentioned the considerable abundance of coccoliths in shallower shelf deposits. Furthermore, Scholle and Kling (1972 have reported concentrations of coccoliths forming up to 20 percent of the sediment in lagoonal environments off British Honduras at depths no greater than 140 feet (43 m).
In a summary of chalk genesis, Tarr (1925, p. 253) cited the conclusions of workers who believed in great depth of deposition but noted that much of the chalk may have been deposited in depths less than 20 fathoms (37 m). In the past few decades, opinion has shifted in favor of shallower water origin for the European chalk. Boswell (1933, p. 201) stated that "conclusions as to the evidence of fossils would have been stated more boldly in favor of shallow depths but for considerations of the early globigerina ooze analogy." Recent study of percentages of planktonic foraminifera (Barr, 1961) suggests that the British Chalk need not have been deposited at depths much greater than 90 m and, for some parts of the section, at depths much greater than 30 m. By the use of discriminant functions to maximize differences in foraminiferal assemblages, and by comparing Chalk Marl foraminifera with comparable living forms Burnaby (1961) concluded that definite fluctuations occurred, with definite deep and shallow water phases. For the latter he estimated depths as little as 5 fathoms (9 m). For a higher part of the British section and on the basis of comparison with modern sponge bathymetry, Reid (1962) concluded that the Late Turonian Chalk Rock was deposited in water not less than 50 fathoms (92 m) deep and not necessarily more than 110 fathoms (201 m). For the Chalk as a whole he (Reid, 1968, p. 558) stated that the hexactinellid sponge evidence indicated depths of at least 100 m but not necessarily more than 200 to 300 meters. Continental geologists also have been revising to smaller figures the maximum depth of deposition of the Late Cretaceous chalks. Nestler (1965, p. 113) concluded that the benthos of the German Rügen Chalk could have lived at depths ranging from 100 to 250 meters. This is in marked contrast to the estimates of Voigt (1929) who, on basis of bryozoan abundance, concluded that the Rügen Chalk was deposited at depths ranging from 300 to 1000 m or more. Steinech (1965, p. 195) believed that this formation was deposited at depths not greater than 300 m on grounds of comparison with modern depth ranges of some brachiopod genera found in the Rügen Chalk.
European and British chalk deposits contain numerous hardgrounds, nodular beds of chalk formed by submarine lithification. Voigt (1959, p. 144-145) has summarized some of the main arguments regarding hardground formation. Many authors believe hardgrounds represent subaerial exposure of carbonate muds, others believe that the hardgrounds represent submarine lithification during a period of shallowing and sedimentary stillstand when the sea floor had become sufficiently shallow to be affected by wave and current action. The occurrence of numerous, closely spaced incipient or fully developed hardgrounds, as in the section at the east cliff, Dover, England, suggests that the. sea floor was subjected repeatedly to shallow water conditions. The origin of hardgrounds during episodes of shallowing is a view shared also by Bromley (1967, p. 507). For the well-known English hardground known as the Chalk Rock, he suggested that glauconitization occurred during shallowing to depths not less than 100 m and that phosphatization in this bed occurred during shallowing to depths not less than 50 m. In the Irish "chalk" near Glenoe, Northern Ireland, Mr. Robin Reid showed me some stromatolitic structures associated with a hardground which apparently represents near-sea-level conditions. For chalk deposits with hardgrounds the range of possibilities of depth of deposition is wide, but relatively shallow water, rather than oceanic depths, seems more likely.
Hudson (1967) concluded that the softness of the English Chalk is related to absence of aragonite in sediments initially. He interpreted this as the result of aragonite solution in a bathymetric range where particulate calcite is not dissolved. He stated that this would occur between depths of about 150 m and 280 m.
The preponderance of recent opinion favors deposition of the English and European chalks at depths less than 300 m with the sea at times less than 50 m in depth.
Little work as been done with regard to determination of Upper Cretaceous chalk deposition in the Western Interior and adjacent seas of North America. So far as known at present no European-type hardgrounds are present. Probable bathymetric ranges of 20 to 80 or 100 fathoms (37 to 146 or 183 m) were suggested by Scott (1940, p. 322) for the Eagle Ford and Austin groups of Texas on the basis of ammonite morphology. In a comparison of overall faunal characteristics of the Austin Chalk and Taylor Marl of Texas, Clark and Bird (1966, p. 323) stated that the range of depositional depth could be from 200 feet (61 m) to 6000 feet (1830 m) but this range was reduced to between 200 and 1600 feet (61 m and 488 m) on the basis of accompanying lithologic characteristics. Farther north the Niobrara Chalk is the Austin equivalent. For this unit, primarily physical evidence prompted Miller (1968, p. 18) to conclude that the Niobrara of Kansas was deposited in water only 40 m deep. The abundant occurrence of Pycnodonte specimens in the Fort Hays Limestone Member led Frey (1972, p. 48) to suggest that initial deposits of the member represent deposition in waters possibly less than 50 feet (15 m) deep with progressive deepening later resulting in deposition of the Smoky Hill Member at depths of 200 to 500 feet (61 to 153 m). These last figures are based largely on depth estimates made by Kauffman (1969, p. 238) who estimated bathymetric ranges of oysters by comparison with modern analogs.
These interpretations of relatively shallow depths lack agreement with depths estimated by Asquith (1970). Basing his interpretation on considerations of depositional topography and with data corrected for compaction, he determined depths exceeding 2000 feet (610 m) for deposition of submarine slope deposits of the lower Cody Shale in Wyoming, and, by implication, for the correlative Niobrara deposits of the adjacent basin.
For the Greenhorn Limestone, Kauffman (1969) has suggested depths of deposition ranging from 100 to 500 feet (31 to 153 m). Much greater depths were postulated by Eicher (1969a) who concluded from foraminiferal evidence that the central (eastern Colorado) part of the seaway during Greenhorn deposition was 1640 feet (500 m) or more in depth. From determinations of paleoslope he (Eicher, 1969a) postulated maximum depth of 2000 to 3000 feet (610 to 915 m). My own (Hattin, 1971, p. 421) estimation for maximum depth of Greenhorn deposition is founded on stratigraphic considerations. The terminal deposit of the Greenhorn Cyclothem (Codell Sandstone Member of Carlile Shale) is a thoroughly bioturbated shallow water deposit representing slow sedimentation prior to a period of sedimentary stillsand and sublevation. The stratigraphic interval from the top of the Greenhorn to the Codell is a maximum of 300 feet in west-central Kansas. Allowing a generous 50 percent compaction factor this interval was originally only 600 feet thick. If the minimum depth of Greenhorn deposition was near the 1640 feet (500 m) suggested by Eicher (1969a), then even after 600 feet of post-Greenhorn sediment was laid down, the entire western Kansas area would have had to be uplifted at least 1000 feet to bring the sea floor into shallow water depths by the inception of Codell deposition. We have no physical evidence for significant uplift of the area during Greenhorn deposition. Assuming that the Codell is a shallow water sand body, and that the pre-compactional thickness of Carlile Shale was about 600 feet, the maximum depth at the beginning of Carlile deposition would be about 600 feet (183 m). This figure makes no allowance for progressive compaction of the Greenhorn, or of continued subsidence, during Carlile deposition.
Whereas an abundance of fossil planktonic foraminifera may in general suggest deep water deposition, these organisms may be abundant also in shallow water habitats (Phleger, 1960, p. 242). The very high ratios of planktonic to benthonic foraminifera in the Greenhorn are not, in my opinion, an indication of the great depths suggested by Eicher, but are a reflection of bottom conditions unfavorable to the establishment of large numbers of benthonic foraminifera. In the absence or near absence of benthonic forms, the planktonic-benthonic ratio would be high for sediments deposited at any depth where planktonic foraminifer tests were settling to the bottom. As pointed out by Phleger (1960, p. 242) planktonic foraminifera can be found in abundance in inner shelf environments. Although an abundance of planktonic foraminifera may be an indication of open sea conditions, an abundance of these forms can also be an indication of high organic production (Bandy and Arnal, 1960, p. 1927). Certainly this last condition is indicated by the organically rich shaly chalk beds and the vast numbers of coccoliths in the Greenhorn. I see no compelling reason to assume that a high planktonic to benthonic foraminiferal ratio has any relationship to depth in an unusual environment such as that represented by the Kansas part of the Western Interior Sea during Greenhorn deposition.
In his comprehensive analysis of Late Cretaceous macroinvertebrate assemblages of the Western Interior region Kauffman (1967, p. 122) stated that small Ostrea, like O. beloiti, occur today in inner shelf waters at depths of 10 to 200 feet (3 to 61 m). In this assemblage small, inequivalve robust Inoceramus, I. rutherfordi, is a common associate. In Kansas this assemblage characterizes the upper part of the Graneros Shale and occurs locally in the basal part of the Lincoln Member of the Greenhorn. Kauffman (1969, p. 238) concluded that the forms occupied a depth range of 100 to 200 feet (31 to 61 m). At Locality 13 the lower part of the Hartland Member contains small assemblages consisting almost exclusively of the paper pecten, Syncyclonema?. Kauffman (1969, p. 240) stated that today such forms are found at mid- to outer-shelf depths of 200 to 500 feet (61 to 153 m). For the sort of assemblage preserved in the Sciponoceras gracile Assemblage Zone, Kauffman (1969, p. 239) postulated a depth range of 300 to 500 feet (92 to 153 m). Assemblages containing flat Inoceramus covered with small oysters and commonly having other epizoans such as stalked barnacles and serpulid worms, and also containing diverse ammonites were ascribed to depths raneing from 200 to 500 feet (61 to 153 m) by Kauffman (1969, p. 238). This assemblage is most typical of the Fairport Member, Carlile Shale, but a modified version of it occurs in the lower part of the Pfeifer Member. A small, squamous form of Pycnodonte, occurring on large, flat Inoceramus, occurs in the middle part of the Jetmore Member and is another version of the same assemblage. Kauffman's interpretations suggests depths of Greenhorn deposition ranging from 100 to 500 feet (31 to 153 m), with minimum range of 100 to 300 feet (31 to 92 m). The shallowest depths are for the upper Graneros and lowermost Lincoln. His estimates are entirely compatible with the physical evidence of decreasing levels of environmental energy upward from the basal Lincoln calcarenites, and with my own conclusions regarding maximum depth as determined from considerations of sedimentation and subsidence. The total absence in all thin sections of evidence of algal borings in valves of Inoceramus, oysters, etc., is accepted as reasonable evidence for minimum depth greater than 60 to 80 feet (18 to 24 m) (see Swinchatt, 1969, p. 1934; Halsey and Perkins, 1970, p. 565).
Bottom Conditions and Substrates
Through much of Greenhorn deposition the sea floor consisted of impure carbonate mud that lithified eventually to form fairly homogeneous shaly chalk. This sort of mud bottom was an unfavorable habitat for most forms of benthonic life; on such muds the macro invertebrate epifauna was restricted largely to inoceramid bivalves and the infauna was practically nonexistent. Benthonic foraminifera were mostly rare on such bottoms (Eicher and Worstell, 1970a, figs. 9, 10).
The preservation of thin laminae in many beds and paucity of current-formed structures such as lenses of skeletal debris are evidence that bottom currents were nil. Fagerstrom (1964, p. 1205) suggested that the mutual occurrence of pelagic and benthonic organisms may be evidence that a preserved assemblage has not been altered by action of bottom currents; such assemblages prevailed generally on Greenhorn chalk-producing substrates. I suggest that poor circulation of bottom waters, and resulting low levels of dissolved oxygen were the principal factors limiting the benthonic faunas in most shaly chalk units. Relatively high content of organic carbon in analyzed shaly chalk samples (average 3.3 percent for 20 nonweathered samples) and near absence of burrow traces in such rocks, suggesting oxygen deficiency in interstitial waters, were a consequence of poor circulation in this environment. A modern parallel has been reported by Calvert (1964) who determined that in the Guaymas Basin, Gulf of California, laminations are preserved where low levels of dissolved oxygen in bottom waters preclude development of a mobile infauna. If Greenhorn sediments had been well oxygenated, one would expect, in such a muddy substrate, a preponderance of deposit-feeding organisms (Purdy, 1964, p. 254) but evidence of any kind of deposit feeders is rare in shaly chalk strata. During deposition of these beds the surficial sediments probably were relatively stable and turbidity of overlying water was low owing (a) to lack of sediment resuspension by deposit-feeding organisms (see Rhoads and Young, 1970), and (b) to lack of mechanical agitation of the sea floor. In such an environment the broad-valved, suspension-feeding inoceramids thrived, apparently by virtue of tolerance to dissolved oxygen levels too low for most other macroinvertebrates. Low taxonomic diversity is characteristic of benthonic environments having low levels of dissolved oxygen (Rhoads and Morse, 1971, p. 419). Except in certain stratigraphic zones, described below, epizoans are rare on valves of inoceramids preserved in shaly chalk beds.
Carbonate muds that lithified to form chalky limestone beds of the Hartland and Jetmore Members, and equivalent parts of the Bridge Creek Member, differ from the shaly chalk-producing muds in two important respects. The former were much more pure than shaly chalk-producing muds and are extensively burrow mottled. The burrow-mottled chalky limestones represent periods of reduced influx of terrigenous detritus (Hattin, 1971, p. 427). Although these beds lack evidence of increased current activity, oxygen content in sediments apparently was greater so that an extensive, mobile infauna of sediment-ingesting organisms could be sustained. It is suggested that these purer sediments were more permeable than those containing greater quantities of fine-grained terrigenous detritus, and that the reduced rate of sedimentation also involved lesser incorporation of organic matter into the sediment. In sediments containing large quantities of organic matter, available oxygen is completely consumed; apparently this happened in the case of shaly chalk-forming substrates. Chalky limestones seem to record the opposite effect because organic carbon levels are low in analyzed samples (average 0.28 percent in 10 nonweathered samples), and the rocks are correspondingly lighter in color than the shaly chalks. Similar environments have been reported from the Santa Barbara Basin off California by Berger and Soutar, (1970, p. 275), who reported that below sill depth available oxygen is readily consumed and resulting sediments are dark colored and do not host burrowing organisms. Above sill depth, where the basin is well oxygenated, the sediments are lighter-colored and have been homogenized by the burrowing benthos. Although this recent example is not a true analog of the Greenhorn environment the influence of oxygen availability on composition and structure of the rock is parallel. I did not detect in the Greenhorn rocks the kinds of differences in skeletal composition and abundance reported from the contrasting environments studied by Berger and Soutar (1970, p. 279). My interpretation of the alternating burrow-mottled and non-burrowed beds of the Hartland and Jetmore Members is the same as that set forth by Hallam (1967, p. 210) for alternating red, mottled, and greenish laminated marls of the Jurassic Ammonitico Rosso of Italy and southern Switzerland.
In comparison with shaly chalk beds of the Greenhorn many of the chalky limestone beds contain a greater frequency of macro invertebrates in addition to Inoceramus. This may be in part a result of reduced sedimentation rate and in part owing to the greater likelihood of finding specimens in chalky limestone. Nevertheless, a few beds, like HL-1 and HL-2, contain not only greater numbers of macroinvertebrates but also greater diversity of both pelagic and benthonic forms. That the added benthonic forms in these beds, especially including Phelopteria and Cerithiella, represent a real change in benthonic conditions is manifested by increase also in this stratigraphic position of calcareous benthonic foraminifera (Eicher and Worstell, 1970a, p. 276). Special local conditions were not responsible for the increased macroinvertebrate diversity recorded in beds HL-1 and HL-2 because these beds are part of the Sciponoceras gracile Assemblage Zone which shows similar increase of diversity across much of the Western Interior region. Eicher and Worstell (1970a, p. 278) attributed the increased diversity of benthonic foraminifera in this zone to widespread improvement of water circulation in the Western Interior sea.
Above the Shellrock limestone bed (JT-13) burrow structures are sparse in all beds of chalky limestone. Although these beds were probably initiated in the same way as Hartland and Jetmore chalky limestones, the chalky limestone-producing substrates of the Pfeifer did not support a diverse endobenthos nor did they support large endobenthonic forms. Sparse mineral-filled burrows were recorded in irregular discontinuous, shell-rich concretionary beds of the Pfeifer, and parts of these beds have an isotropic probably bioturbated fabric, but some of these beds contain good evidence of internal stratification. More continuous beds like PF-1 and the Fencepost limestone bed lack internal stratification and sparse burrows, consisting mostly of minute, calcite-filled structures, occur especially in the Fencepost bed. The isotropic fabric of these two beds as viewed in thin sections suggests that these units also have been bioturbated, but intensive weathering at all exposures studied apparently has eradicated evidence of extensive burrowing.
The general condition of poor bottom circulation during Greenhorn deposition has exceptions. The upper half of the Jetmore Member, from about the level of marker bed JT-8 to shortly above JT-12, and the lower two-thirds of the Pfeifer, from shortly above JT-13 nearly to the Sugar sand (PF-2), are characterized by a profusion of valves and valve fragments of inoceramids as well as by small, commonly poorly washed lenses of foraminiferal and/or inoceramid debris. The epizoal oysters Pycnodonte and Pseudoperna, locally in large clusters, scattered remains of pedunculate cirripeds, and anomiids are a conspicuous element of the macroinvertebrate fauna in shaly chalk and chalky limestone beds. These fossils are interpreted as the result of generally improved circulation that produced conditions favorable for proliferation of the macroinvertebrate epifauna, including the slowest rates of sedimentation during deposition of the Greenhorn Limestone. Impingement on the sea floor of gentle currents brought supplies of dissolved oxygen to the otherwise nearly stagnant bottom and served to sweep skeletal debris into small heaps through much of the designated intervals. These conditions prevailed during periods of slow as well as more rapid sedimentation because the increased numbers and diversity of macroinvertebrates occurs in both shaly chalk and chalky limestone beds of the two intervals. On an arbitrary energy scale of 1 to 4 these beds represent energy-level 2. Most of the Greenhorn represents energy level 1.
A still higher level of energy expenditure by sea floor currents is represented by the thin, discontinuous beds and zones of lenses of well-washed, commonly cross-laminated skeletal limestone that characterizes most Lincoln sections. The widespread bed of foraminifer- and calcisphere-rich limestone lying between Jetmore marker beds JT-1 and JT-2, and a few lenses in the lowermost Pfeifer also are in this category. Limestones of this sort represent more thorough washing of sediment during short-lived episodes of sea floor scour that set finer sediments in suspension and produced lag concentrates of skeletal debris as much as 0.4 foot thick. On the arbitrary energy scale mentioned above, these rocks represent energy level 3.
Units of crossbedded skeletal limestone as much as 1.5 foot thick and local lenses, possible pararipples, as much as 0.5 foot thick, lie at the base of the Lincoln Member at many Kansas localities, especially in the area extending from Ford County to Jewell County. At most places these thicker units of limestone rest with sharp contact on underlying Graneros strata. These limestones consist largely of Inoceramus debris, especially isolated prisms, as well as shark teeth, bentonite pebbles, coprolites, bits of fossil wood, local intraclasts, and reworked Graneros fossils. Diachroneity of the Graneros-Greenhorn contact (Hattin, 1968; this paper) suggests that these prominent lag-concentrate limestones were produced as an offshore, high-energy belt (Shaw, 1964; Irwin, 1965, p. 450) swept progressively eastward or northeastward during the Greenhorn transgression. The attending environment supported a benthonic macroinvertebrate community consisting principally of Inoceramus but supporting also at least 3 species of oysters, a hamitid, and sparse cirripeds. The shifting substrate precluded all but local establishment of abundant macroinvertebrates and only locally do the numbers of Inoceramus or oysters compare with those of the upper Jetmore or lower and middle parts of the Pfeifer. On the arbitrary energy scale these beds represent energy level 4, the highest represented in the Greenhorn Limestone of Kansas.
Salinity
The broad uniformity of fauna, flora, and lithology in Greenhorn rocks suggests that salinity remained essentially constant throughout the period of deposition. Most abundant of all identified organic remains are coccoliths which characterize shaly chalk units throughout the formation. In present-day seas most species of coccolithophores cannot tolerate seawater having a salinity that departs widely from that of the open ocean (Black, 1965, p. 136), and few species of coccoliths can flourish at salinities greater than 38‰ or less than 25‰. The extant species Braarudosphaera bigelowi (Gran & Braarud), known to be tolerant of salinities as low as 17‰, (Bukry, 1974, p. 358), and uncommon in waters of full marine salinity (ibid.), was reported only from the upper part of the Jetmore and the Pfeifer Member in a study of the Greenhorn section at Locality 3 (Russell County) by Cepek and Hay (1969, p. 327) and was not recorded in Greenhorn samples from any of the Colorado, Wyoming and South Dakota localities studied by Trexler (1967). Furthermore, the variety of coccoliths recorded in the Greenhorn (Trexler, 1967; Cepek & Hay, 1969) is comparable with that of equivalent rocks in the Gulf Coast section (Gartner, 1969) and suggests water of normal salinity.
Second in observed abundance in Greenhorn rocks are tests of planktonic foraminifera. Eicher and Worstell (1970a) reported a total of 22 planktonic species from the formation at Localities 3, 13, and 14, with as many as 15 of these species occurring in a single lithic unit. Recent planktonic foraminifers are found characteristically in marine waters of normal salinity (Smith, 1955, p. 147; Phleger, 1964, p. 34). Although diversity of these fossils differs from bed to bed within the Greenhorn, the foram assemblage is everywhere dominated by planktonic forms. This evidence is accepted as indicating water of normal or close-to-normal salinity. Stratigraphic variations in planktonic foraminifer diversity documented by Eicher and Worstell (1970a) may represent fluctuations in salinity but if so, these variations are judged to have been small. According to Douglas (1972, p. 26) the northern limit of oceanic conditions in the Western Interior is marked by the limit of keeled planktonic foraminifers. During Greenhorn deposition this boundary lay well to the northwest of central Kansas.
Virtually all parts of the Greenhorn contain ammonites, although these fossils are not distributed uniformly through the section. Largest concentrations appear in the lower part of the Lincoln, in the Sciponoceras gracile zone of the Hartland and Bridge Creek (twelve species), in the Jetmore Member, and in the Fencepost limestone bed. With few exceptions modern cephalopods are strictly marine organisms (MacGinitie and MacGinitie, 1949, p. 389) and inhabit marine water of normal salinity (Scott, 1940, p. 308). Kummel (1948, p. 64) noted that cephalopods apparently cannot endure decreased salinities but may be able to tolerate above-normal salinity. Cretaceous cephalopods, like nearly all modern ones, most probably inhabited waters of normal salinity; this conclusion is reinforced by the fact that in the Greenhorn cyclothem, beds believed on other evidence to represent brackish water environments are lacking in cephalopod remains. The upper part of the Dakota Formation (Hattin, 1967; Siemers, 1971) and the lower part of the Graneros Shale (Hattin, 1965a) have been lithologically and faunally linked with brackish water environments. Even where remains or molds of originally aragonitic skeletons are preserved abundantly in these units, the remains of cephalopods are lacking in Kansas.
Pycnodont oysters, such as characterize part of the Jetmore Member, are characteristic of normal marine environments according to Stenzel (written communication, 1972). This genus, and the small oysters of the Pfeifer Member are apparently ancient counterparts of the living Ostrea equestris, a similarly small, adnate form which is characteristic of higb-salinity waters (Abbott, 1954, p. 373; Parker, 1955, p. 203).
The abundance of Inoceramus in most parts of the Greenhorn is also taken as evidence of normal or nearly normal marine salinity. In the Greenhorn cyclothem this genus has stratigraphic distribution nearly parallel with that of ammonites. Vokes (1947, p. 128) believed that Inoceramus was normally an inhabitant of water of normal salinity. Inoceramids have not been recorded in brackish water assemblages of the upper part of the Dakota Formation in Kansas and are very sparse in the lower part of the Graneros Shale. They are abundant in the upper part of the Graneros Shale where small oysters, ammonites, and planktonic foraminifera collectively indicate waters of normal or near normal salinity.
Temperature
More than half a century ago knowledge of climatically controlled latitudinal differentiation of Cretaceous biotas was advanced sufficiently to be summarized in some detail by Dacqué, (1915, p. 423-426) in his classic textbook on paleogeography. At that time geologists recognized a warm water Tethyan or Mediterranean belt, characterized by rudists, corals, actaeonellids, nerineids and larger calcareous foraminifera, and a cool water boreal belt characterized by belemnites. Reeside (1957, p. 512) agreed that the Mediterranean realm had a warm or perhaps tropical climate but noted that the more northerly (boreal) realm had a temperate, possibly cold temperate, but not an arctic climate. Jeletzky (1971, p. 1647; 1971a, p. 12-13) also pointed out that the term "boreal" is misleading because paleontological evidence from the higher latitudes indicates at least a warm temperate climate "everywhere in the Cretaceous 'Boreal' Realm." Kauffman (1973, p. 367) has substituted the term "north temperate realm" for "boreal" because in this belt the fauna consists dominantly of warm to mid-temperate organisms. According to Sohl (1971, p. 1611) the Cretaceous of North America was characterized throughout its history by three main biotic provinces including (1) a Caribbean-Central American-Baja Californian province having a warm water (tropical-parentheses mine) fauna of Tethyan affinities, (2) a Coastal Plain-southern Western Interior province having a subtropical to warm temperate assemblage, and (3) a California-northern Western Interior province having a boreal fauna. In a review of biogeographic data on Cretaceous foraminifera, Bergquist (1971, p. 1567, 1568) recognized a climatic zonation comprising tropical, warm temperate, mild temperate, cool temperate, and cold zones. According to this scheme the warm temperate zone included the area of the present Gulf Coastal Plain, the mild temperate zone embraced the southern part of the Western Interior region (including Kansas), the cool temperate zone included the northern part of the U.S. Western Interior and all of western Canada, and the northern cold zone included the Arctic portion of Alaska and Canada. It is doubtful that the Western Interior region was as finely zoned climatically as Bergquist (1971) suggested. Indeed, Kauffman (1973, p. 367) has noted that the Cretaceous north temperate realm contains "no truly Arctic faunas and has few north-temperate bivalves." As Valentine (1967) has pointed out, the latitudinal range of individual species is narrow at times of increased climatic zonation. In the Western Interior region many Late Cenomanian and Early Turonian species have wide latitudinal distribution and the faunas include many European and cosmopolitan species. These observations suggest broad climatic zones of equable temperature. Despite contradictory statements regarding whether the so-called boreal region was cold temperate or warm temperate, pronounced south-to-north differences in Western Interior faunas have been documented extensively (e.g. Sohl, 1971; Kent, 1969; Bergquist, 1971; Eicher, 1969) and climate, i.e. temperature, is accepted generally as the controlling factor.
Certain taxa have been regarded as biogeographically diagnostic. In the southern part of the Western Interior region the Greenhorn Limestone includes endemic species like Exogyra aff. E. boveyensis, Pycnodonte sp. A, Anomia sp. A, and Pseudoperna bentonensis as well as cosmopolitan species like Mytiloides labiatus, Mammites nodosoides, and Collignoniceras woollgari. Some species are migrants from other parts of the same faunal province, for example Acanthoceras wyomingense and Dunveganoceras spp. which are characteristic of the northern Western Interior and western Canada. The occurrence of A. wyomingense only sparingly in the southern Western Interior and the discovery of only a few specimens of Dunveganoceras in the Greenhorn suggests that the Kansas area lay at the southernmost geographic limit of these forms. No belemnites have been recorded from the Kansas Greenhorn and only four specimens are known from the overlying Fairport Chalk Member of the Carlile Shale. This evidence suggests that Kansas lay at or below the southern limit of the northern Western Interior province (Sohl, 1971) in late Cenomanian and early Turonian times.
Although rudists have been recorded sparingly in the Niobrara Chalk of Kansas, none have been discovered in the Greenhorn. Likewise, no actaeonellids, corals, or nerineids are known from the Kansas Greenhorn, and only one coral specimen has been recorded from the Greenhorn farther to the north (Cobban, 1951, p. 2184). The Kansas area lay north of the influence of the Tethyan faunal realm. Jeletzky (1971, p. 1654; 1971a, p. 76) noted that the Western Interior south-to-north temperature gradient is expressed by gradual northward disappearance of the Tethyan elements. Studies of gastropod assemblages by Sohl (1971) and foraminiferal assemblages by Bergquist (1971) yield evidence that is in substantial agreement with this concept. During Greenhorn deposition Kansas lay near the southern limit of the north temperate realm; the faunas have a strong affinity with Gulf Coastal Plain assemblages rather than with those of the Arctic area which suggests a warm-temperate rather than a cool-temperate climate. This conclusion is borne out by the evidence of occasional incursions of warm water gastropods (Sohl, 1971, p. 1620) and planktonic foraminifera (Bergquist, 1971, p. 1602) farther northward than usual during the time of Greenhorn deposition.
Oxygen isotope studies of Late Cretaceous fossils, largely belemnites, have been used extensively in paleotemperature analysis. Data from many parts of the world confirm a pattern of poleward temperature decline throughout the Late Cretaceous of the Northern Hemisphere (Lowenstam, 1964, p. 244). A tabulation of North American analyses was presented by Lowenstam and Epstein (1954, p. 220-22-9) but most of their specimens are from Maastrichtian units of the Atlantic and Gulf Coastal Plains. These analyses yielded temperatures ranging from 16° to 23°C which suggests a marginally subtropical environment. Nonbelemnoid analyses, mostly from Coon Creek, Tennessee bivalves, yielded paleotemperatures ranging from 20° to 28°C which suggests a subtropical environment (ibid., 1954, p. 229). Later, Lowenstam and Epstein (1959, p. 67) reported a baculite analysis from the Coon Creek locality that indicated a paleotemperature of only 17°C.
Few belemnite analyses have been reported from the Western Interior region; however, Lowenstam and Epstein (1954) reported paleotemperatures ranging from 26° to 31°C for three specimens from the Colorado Group at Fort Benton, Montana and 23°C for belemnites from the Coniacian near Winnecook, Montana (Lowenstam and Epstein, 1959, p. 68). These temperatures suggest a marginally tropical environment and are in conflict with statements by those who concluded in favor of a warm to mild temperate climate for the Western Interior region. Similarly high paleotemperature values were recorded by Tourtelot and Rye (1969) for specimens of Baculites and belemnites of Campanian and Maastrichtian age from localities distributed between northern New Mexico and southern Canada. In this area they recorded temperatures averaging 21° to 33°, by state, the wide variation being interpreted as the result of isotopic variation within the sea that deposited the Pierre Shale. The generally higher temperature values for Western Interior localities as compared with localities peripheral to the continent was interpreted as resulting from isotopically lighter water owing to meteoric water influx (ibid., p. 1920). These authors believed that the paleotemperature data do not encourage use of oxygen isotopes for climatic analysis of the North American Late Cretaceous.
Despite discrepancies at individual localities for data derived from different organism groups, it is desirable to continue the gathering of data, especially from areas not previously sampled. The Fairport Chalk Member of the Carlile Shale represents deposition under apparently normal marine conditions, far from any shoreline. Belemnites from two localities in Ellis County, Kansas were processed for oxygen isotope analysis in order to compare the paleotemperature values with those of other Western Interior localities, farther west (Tourtelot and Rye, 1969) and with the Gulf Coast values (Lowenstam and Epstein, 1954; 1959). These specimens yielded paleotemperatures of 65°C and 48°C. These temperatures are impossibly high and suggest that the specimens submitted for analysis have suffered considerable alteration of the skeletal carbonate.
Paleoecology of Major Invertebrate Groups
Foraminifera
Tests of foraminifera are detected readily in many Greenhorn beds and are commonly concentrated in laminae, lenses, and thin beds consisting principally of these fossils. Foraminifera were observed in all of the more than 300 thin sections examined in this study. The recent, excellently illustrated study of Greenhorn foraminifera by Eicher and Worstell (1970a) made detailed treatment of the group unnecessary. However, my petrographic study confirms the overwhelming preponderance of planktonic species as reported by Morrow (1934, p. 188) and by Eicher and Worstell (ibid.), as well as the overwhelming predominance of forms belonging to the genera Hedbergella and Heterohelix.
The writer (Hattin, 1965a, p. 55) recorded within the Graneros Shale a stratigraphically upward change in foraminiferal assemblages from those consisting wholly of agglutinated forms to those containing a large proportion of planktonic forms. This change was interpreted as resulting from gradual establishment of normal salinity as open-sea conditions developed during the Cenomanian transgression. The predominance of planktonic species in all Greenhorn foraminiferal assemblages reflects approach to and attainment of the transgressional maximum during deposition of the formation. This conclusion is in substantial agreement with that of Eicher and Worstell (1970a, p. 270) who noted that the change to predominantly planktonic assemblages corresponds nearly with the Graneros -Greenhorn contact and coincides with~"the opening of the western interior seaway to major open ocean currents." They recognized three major Greenhorn foraminiferal zones: (a) lower planktonic zone, (b) benthonic zone, and (c) upper planktonic zone. The absence of calcareous benthonic species in (a) was judged by them to be a result of unfavorable bottom conditions, probably low levels of oxygenation. The appearance of diverse species but mostly small numbers of calcareous benthonic forms in (b) was ascribed to improved bottom conditions resulting from improved circulation (Eicher, 1969a, p. 1078; Eicher and Worstell, 1970a, p. 278). In Kansas (b) is only 5 to 7 feet thick and corresponds to the Sciponoceras gracile Assemblage Zone which is unique with regard also to the diverse macroinvertebrate fauna that it contains. In this zone the abrupt and widespread appearance of a diverse benthos on a bottom occupied earlier almost exclusively by Inoceramus suggests rapid response to improved physical conditions by opportunistic species (see Levinton, 1970). The upper planktonic zone reflects return to "limiting environmental circumstances" (Eicher and Worstell, 1970a, p. 280) similar to those of the lower planktonic zone. They note, however, that the disappearance of benthonic forms is not abrupt; calcareous benthonic species persist sparingly into the Fairport Member, Carlile Shale. Contradiction to their suggestion that increased circulation explains abrupt development of the benthonic zone is seen in skeletal limestones that characterize much of the Lincoln Member, and occur also in the upper part of the Jetmore and the lower part of the Pfeifer. These skeletal limestones imply as good or better circulation of bottom waters as during deposition of the Sciponoceras gracile zone, but the accompanying strata are deficient in or lacking benthonic foraminifera. It seems that the abrupt appearance of the Sciponoceras gracile assemblage, including its benthonic foraminifera, requires an explanation less general than "increased circulation" especially when one recalls the enormous geographic distribution of the zone.
On the basis of percentage of planktonic foraminifer specimens Eicher (1969a, p. 1079) concluded that the lower part of the Bridge Creek Member, i.e., the Sciponoceras gracile, zone, was deposited at a minimum depth of 1640 feet (500 m). For Greenhorn beds below the S. gracile Assemblage Zone, which contain no benthonic foraminifers, Eicher (1969a, p. 1076) postulated a silled-basin environment and stated that for such a basin depth estimates based on proportion of planktonic foraminifera would be meaningless. Throughout the Graneros Shale and Greenhorn Limestone, assemblages of benthonic macro- and microinvertebrates lack the broad diversity that is characteristic of modern open-ocean floors. During Graneros and Greenhorn deposition bottom conditions obviously were not conducive to proliferation of a diverse benthos; this makes direct comparison with modern open-sea environments very tenuous. Whereas I agree with Eicher that foraminiferal ratios are of little utility when applied to the Greenhorn beds having no benthonic foraminifera, I do not believe that the mere presence of benthonic foraminifera in the overlying beds is in itself basis for valid application of foraminiferal ratios to the problem of paleobathymetry. Conditions may have improved sufficiently to foster growth of sparse benthonic foraminifera in the S. gracile Assemblage Zone and overlying beds, but macroinvertebrate and sedimentologic data suggest that the sea floor remained, for the most part, poorly oxygenated.
Ammonites
On the basis of gross morphology, which may have significance with respect to, mode of life, Greenhorn ammonites are classifiable into 7 major types. These are 1) partially uncoiled types like Worthoceras and Scaphites, 2) irregularly uncoiled forms like Stomohamites and Allocrioceras, 3) straight uncoiled forms like Sciponoceras and Baculites, 4) involute, nearly smooth, laterally compressed forms like Borissjakoceras, Desmoceras, and mature Tragodesmoceras, 5) robust, coarsely ribbed, horned or spined ammonites like Acanthoceras, Mammites, Colligoniceras, and Kanabiceras, 6) robust, densely ribbed, tuberculate forms like Eucalycoceras, Calycoceras, Pseudocalycoceras, and Watinoceras, and 7) involute, ribbed, laterally compressed forms like Metoicoceras and youthful Tragodesmoceras. This grouping differs somewhat from that of Scott (1940) and represents only 4 of the 7 groups he described. Opinion regarding life habits of the ammonites are diverse and no consensus has been reached; Arkell (1957, p. L120) stated that debate on the subject is at least 90 percent conjecture. Reyment (1958) summarized in considerable detail the contradictory conclusions reached by various cephalopod workers over a 68-year period beginning in 1890. An example of this diverse opinion is found in Birkelund (1965, p. 145) who recounted that different authors have interpreted the scaphites as benthonic, nektonic, or planktonic forms. Excepting certain uncoiled forms Scott (1940, p. 320) concluded from evidence of lithologic association that most Texas Cretaceous ammonites were nektobenthonic and that depth was a major factor controlling distribution. In agreement with Scott, both Arkell (1957, p. L122) and Ziegler (1967) believed that most ammonites were nektobenthonic. Ziegler (1967) believed that in the European Upper Jurassic ammonite distribution was strongly controlled by depth but he believed (p. 453) that the species and genera are generally independent of the substrate. Arkell (1957, p. L119) also noted that Scott's conclusions regarding relationship of shell form to lithology are not applicable generally. Reyment (1958) concluded that a majority of ammonite conchs floated after death and could be transported widely by marine currents, but Arkell (1957, p. L122) believed that current distribution of larvae was probably more important than post-mortem drifting as a means of effecting wide distribution of ammonite species.
Smooth-shelled, involute discoidal Greenhorn ammonites like Desmoceras and Borissjakoceras are not unlike modern Nautilus and like the last were probably excellent swimmers. Heavily ribbed, strongly tuberculate or horned ammonites like Acanthoceras, Collignoniceras, and Pseudocalycoceras lacked streamlining and apparently were not able to swim rapidly but moved about rather sluggishly, probably close to the sea floor. Modern heavy-shelled Strombus gigas which is both ribbed and horned bears a striking resemblance to certain acanthoceratid ammonites when viewed apically; S. gigas is well adapted to a foraging habit on the sea floor. The chambered acanthoceratids were certainly more buoyant than Strombus, and rather than crawling on the bottom probably utilized their buoyancy to hover above the sediment-water interface.
The baculitids probably swam in schools, somewhat like modern squids. The concentrations of huge numbers of baculitids locally in Hartland marker bed HL-2 and in the Shellrock limestone bespeak a gregarious existence during at least a part of the life cycle. The loosely uncoiled ammonites are among the most curious of all ammonites; why uncoiling occurred is not certain but the question has been reviewed recently by Wiedmann (1969) who concluded that the uncoiling has nothing whatever to do with evolutionary degeneracy. Modern uncoiled mollusks like Vermicularia are strictly benthonic, but lack the chambers possessed by ammonites. Chambers in Stomohamites and Allocrioceras suggest sufficient buoyancy to facilitate short swimming motions but I am inclined to believe that these were essentially benthonic forms. In the Greenhorn they occur almost exclusively in beds that contain benthonic bivalves in addition to Inoceramus. The genus Worthoceras is a most puzzling form as are the true scaphites. The change from regularly coiled to uncoiled and finally to recurved conch in the adult stage suggests that juvenile Worthoceras was a good swimmer and that adults were probably sluggish swimmers. Involute, ribbed, compressed forms like Metoicoceras and juvenile Tragodesmoceras probably were fair swimmers, intermediate in ability between smooth-walled involute forms and the heavily ribbed, horned forms.
Several Greenhorn ammonite species have broad geographic range, including such forms as Acanthoceras wyomingense, Calycoceras? canitaurinum, most species of the Sciponoceras gracile zone, Mammites nodosoides, Watinoceras reesidei, and Collignoniceras woollgari. Furthermore, these species have been recorded in rocks of differing lithology. These facts might be construed as positive evidence for a wholly nektonic existence well above the sediment-water interface and unaffected by bottom conditions. However, several of the accompanying strictly benthonic species, especially including Ostrea beloiti, Inoceramus prefragilis, Mytiloides labiatus, and I. cuvieri also have wide distribution. Therefore, the wide distribution of ammonite species is as easily explained by current spreading of larvae as by migrating swarms of adults.
The occurrence together in a single bed such as Hartland marker bed HL-2 of 6 out of the 7 ammonite forms mentioned above, each of which suggests at least minor difference in mode of life or adaptation to environment suggests some underlying control which influenced also the distribution of the gastropod Cerithiella, Phelopteria and probably also the introduction at or near this stratigraphic position of abundant benthonic foraminifera. The evidence is strong that this host of new forms resulted from rather abrupt improvement of bottom conditions and favors a nektobenthonic interpretation for the ammonites, as suggested by Scott (1940) and Ziegler (1967).
Other Greenhorn beds have less diversity of ammonites than marker bed HL-2. Basal Lincoln skeletal limestones locally contain several species, but the texture and structure of these rocks, as well as erratic distribution and fragmentary condition of many specimens shows that many of the basal Lincoln specimens were transported to the place of burial.
Isolated occurrences of abundant ammonites of small size, like those of Eucalycoceras sp. B in the middle Lincoln at Locality 26, may be explained as resulting from small-scale catastrophes, but it is also possible that such forms are also abundant elsewhere and have been overlooked because of small size and imperfect fissility of the shaly chalks that contain them. The nearly ubiquitous occurrence of mostly juvenile forms of Watinoceras reesidei in Jetmore marker bed JT-1 suggests that this species was distributed as consistently as certain species of the Sciponoceras gracile zone and that W. reesidei suffered high infant mortality. Collignoniceras woollgari is a ubiquitous form in the Fencepost limestone bed and like W. reesidei in JT-1, is mostly preserved as juveniles. If these two species had lived far above the bottom where waters were better circulated it seems likely that more adult specimens of each would be found. It is suggested that bottom conditions were only intermittently suitable for such forms, and that a majority of specimens never reached adulthood. The paucity of ammonite molds in most shaly chalk beds, as compared with the limestone beds, fits well with the conclusion stated above that the lowest energy levels and least favorable bottom environments are represented by shaly chalk beds.
Bivalves
Greenhorn specimens of Exogyra columbella are all from skeletal or intraclastic limestone beds lying at or near the base of the formation. Unlike E. aff. E. boveyensis, E. columbella was attached originally by only a small area of the left valve, and lay with ventral margin upward and with plane of commissure lying at a usually steep angle to the horizontal (Hattin, 1965a, p. 58).
Exogyra columbella is common only at Locality 23, in the lower 1.5 feet of the Lincoln, and is preserved there in conglomeratic skeletal limestone containing pebbles of limestone, bone fragments and teeth, inoceramid and oyster shell debris, and coprolites. Textural evidence suggests a turbulent and probably shallow-water depositional environment such as Jourdy (1924, p. 35) postulated for small species of Exogyra. At Locality 23 excellent preservation of thin Exogyra valves indicates that most of these shells were not subjected to the environmental rigor that broke into pieces many of the ammonite conchs preserved in the same beds. Nonetheless, apparent absence of right-hand Exogyravalves, and nearly uniform convex-up orientation of the collected specimens are evidence that these oysters were transported to the site of burial. During early life the individuals were attached to a solid substrate (shell fragments, etc.) as shown by shell deformation in the beak and umbonal regions of some specimens. In many specimens, as the shells grew, the posterior part of the left valve came to lie directly on the substrate. As a result, some of the left valves are irregularly flattened, the flattened area merging with the rest of the valve along a deflection that essentially marks the sediment-water interface. Assuming originally horizontal orientation of flattened areas and/or areas of juvenile attachment, specimens of E. columbella at Locality 23 lay with planes of commissure inclined at angles 21° to 84° (![]() =54°) from the horizontal in 6 specimens for which this measurement could be made.
=54°) from the horizontal in 6 specimens for which this measurement could be made.
Lincoln specimens of E. columbella lack totally the sponge and other borings that are so commonly observed in thicker shelled Late Cretaceous exogyras such as E. ponderosa, E. costata, E. cancellata, and E. mesabiensis. Absence of borings and possession of thin, small shells suggests for Greenhorn specimens of E. columbella rapid growth, a short life, and early burial. The last factor is consistent with the texture of rocks in which I have collected the species.
In the Lincoln Member of Kansas Exogyra aff. E. boveyensis is more common and widespread than E. columbella. The former species is characteristic of basal Lincoln skeletal grainstones across much of central Kansas, and ranges locally to above the middle of the Member. At Locality 55 (Hodgeman County) E. aff. E. boveyensis occurs well above the base of the Lincoln and at Localities 8 (Ford County) and 12 (Kearny County) it occurs in the upper few feet of the member. Occurrence of this oyster almost exclusively in parts of the section containing an abundance of skeletal grainstone indicates preference for habitats in which relatively strong bottom currents prevailed. Exogyra aff. E. boveyensis is broadly attached by the left valve to valves of Inoceramus prefragilis, and only rarely occurs as clusters in which more than one generation of individuals is represented. The shells are mostly less than 2.5 cm across, are thin, and lack epizoan borings. Collectively these features suggest a short life history for individuals and for clusters, occasioned perhaps by the frequent shifting of substrates by currents. Right valves are usually separated from the left valves, but in the cluster illustrated (Plate 3,D) several articulated specimens can be seen. Only rare specimens of Ostrea beloiti and even fewer specimens of E. columbella have been collected from beds containing E. aff. E. boveyensis.
The small oyster Ostrea beloiti forms a thin biostrome-like bed at the base of the Lincoln Member at Locality 8 (Ford County). Although these oysters are commonly well preserved, the valves are disarticulated, many are broken (Fig. 7,A) and the, bed contains much quartz sand and rests sharply on the Graneros Shale. The species does not range above this bed, which may represent a lag deposit of shells derived locally from the upper Graneros by pre-Lincoln sea floor scour (Hattin 1965a, p. 44). The species occurs also as well-preserved valves scattered through the Graneros-Greenhorn transition beds at Locality 12 (Kearny County); the chalky matrix surrounding these oysters suggests low energy conditions and little transport of specimens. Here also the species does not range upward into more typical chalk units of the Lincoln. In central and northern Kansas the sharp contrast between Graneros and Lincoln lithology, the conglomeratic nature of basal Lincoln strata at many places, and the occurrence of sparse, disarticulated valves of O. beloiti only in basal Lincoln skeletal limestones suggests that here, too, the species has been reworked from the Graneros.
Ostrea beloiti is abundant in the upper half of the Graneros Shale (Hattin, 1965a, p. 40) in central Kansas where it occurs mostly in calcareous quartzose sandstones and skeletal limestone beds, commonly as thin biostromal layers. The upper part of the Graneros is also rich in O. beloiti, including biostromal units, at Locality 12. On lithologic, stratigraphic and paleontologic grounds the writer (Hattin, 1965a, p. 67) concluded that the Graneros oyster-bearing beds were deposited relatively far from shore in waters of normal marine salinity and probably at a depth not exceeding 100 feet. Kauffman (1967, p. 122) stated that recent small Ostrea mostly inhabit the inner shelf regions in moderately quiet water at depths ranging from 10 to 200 feet, and suggested an inner to middle shelf environment for the O. beloiti biostromes of the Western Interior Sea. His conclusions are compatible with my own. The energy levels reflected in basal Lincoln skeletal limestones were unfavorably high for O. beloiti and the coccolith-rich muds that make up the bulk of the Lincoln apparently represent an environment in which energy levels were too low, or the substrate too soft, for colonization by O. beloiti.
Above the basal part of the Lincoln Member Ostrea is not represented, and Exogyra aff. E. boveyensis occurs high in the member only at Localities 6, 8, 12 and 55. Elsewhere all but the basal Lincoln and essentially all of the Hartland Member are devoid of oysters. General absence within this stratigraphic interval of all clearly benthonic forms save Inoceramus, and excepting the Sciponoceras gracile zone, suggests bottom conditions inimical to habitation by epizoal oysters and other epifaunal species. The probable nature of the environment has been discussed elsewhere.
Beginning at the base of the Jetmore Member small, fragile valves of oysters are preserved sparingly in chalky limestones and, rarely, in shaly chalk beds. In the interval extending from the top of marker bed JT-7 to shortly above JT-12, but most abundantly and consistently in the shaly chalk unit lying between marker beds JT-9 and JT-10, a small adnate oyster, Pycnodonte sp. A, is the only conspicuous benthonic form other than Mytiloides. The small fragile valves of these oysters are attached commonly as well preserved clusters (Pl. 8,E) of articulated valves, in growth position, on large inoceramid valves. Abundance of pycnodonts in this interval coincides with abundance of whole and fragmentary inoceramid valves. These oysters are less abundant above the base of JT-10 and apparently do not range to the top of the member. In the lower and middle parts of the Pfeifer Member, as high stratigraphically as the sugar sand unit (PF-2), a niche similar to that of Pycnodonte sp. A is occupied by Pseudoperna bentonensis. The latter species is also commonly well preserved as articulated valves in growth position on large inoceramid valves in a part of the section that is rich in inoceramid remains. The conditions that produced great numbers of inoceramids fostered also the development of the two groups of epizoal oysters. Ample hard substrates, more highly oxygenated bottom waters, and sedimentation rates sufficiently slow to prevent premature burial of oyster spat are believed to be the chief factors permitting development in the Jetmore and Pfeifer Members of abundant epizoal oysters. Common occurrence of perfectly preserved, articulated specimens of Pycnodonte and Pseudoperna is evidence of quiet, non-agitated or little-agitated bottom conditions. In the uppermost part of the Pfeifer and lowermost part of the overlying Fairport Chalk Member, Carlile Shale, where large oblate spheroidal concretions of chalky limestone are abundant, inoceramids are relatively sparse and epizoal oysters are rare, probably owing to reduced circulation and low levels of bottom-water oxygenation.
Kauffman (1967, p. 122) stated that on the modern Atlantic shelf Pycnodonte is the dominant or sole ostreid found in deeper water environments of 50 to 100 to 300 feet, and (1967, p. 124) indicated a quiet water, middle to outer shelf habitat for the genus. For gryphioid Pycnodonte assemblages of the Western Interior Cretaceous Kauffman (1969, p. 238) postulated a depth range of from 50 to 500 feet. For small Cretaceous pycnodonts, apparently including forms like those in the Jetmore Member, he (Kauffman, 1969, p. 238) postulated depths ranging from 200 to 500 feet. He regarded the most common Niobrara oyster, known generally as Ostrea congesta, as small pycnodonts, but Stenzel (1971, p. N1131) referred these to the genus Pseudoperna. The Pfeifer oysters are also Pseudoperna. If Kauffman's interpretation is correct, the small oysters in the Jetmore and Pfeifer Members represent a deepwater environment, probably the greatest represented in the Greenhorn.
Species of Inoceramus are common to abundant throughout the stratigraphic section extending from the upper half of the Graneros Shale to the middle part of the Blue Hill Shale Member of the Carlile Shale. This distribution is parallel with that of ammonites and is accepted as evidence that these species of inoceramids were inhabitants of marine waters of normal salinity. Many of the species recognized in the Greenhorn of Kansas are known also from distant parts of the globe. Examples include Inoceramus flavus Sornay, described originally from Madagascar, Mytiloides labiatus (Schlotheim), described originally from Europe but having essentially worldwide distribution, and Inoceramus cuvieri Woods, described originally from the British Chalk. Presence of these widespread species suggests free communication of the Western Interior Sea with the world ocean. This observation is supported by presence in the Greenhorn of several ammonite species that likewise are known from distant areas.
The only close living relative of the inoceramids is the byssate genus Isognomon which bears only general resemblance to the Greenhorn inoceramids and which has a mode of fixation to solid substrates that is not demonstrable in any Greenhorn species. Highly inequivalved, probably byssate inoceramids are represented by a few specimens of Inoceramus rutherfordi? at the base of the Lincoln Member at one locality, but other Greenhorn species apparently were not attached forms. Many of the latter are preserved in isolated positions within shaly chalk beds that afforded no apparent solid substrate for byssal attachment. Chalky limestone beds containing the same species do not represent former hardgrounds and thus offered no general attachment surface for these bivalves. Most Greenhorn inoceramids were neither semi-infaunal nestlers nor burrowers because none have been observed in a vertical position. Rather, all whole specimens are preserved lying concordant with stratification, suggesting that they merely lay on the carbonate muds with the plane of commissure parallel to the sediment-water interface. The three principle Greenhorn species Inoceramus prefragilis, I. cuvieri, and Mytiloides labiatus all have large, relatively flat or only moderately biconvex valves in the large adult stage of growth and thus were adapted well to a free existence on relatively soft substrates. juvenile forms of these species were likewise adapted to the soft substrate, I. cuvieri by virtue of highly inflated valves, I. prefragilis and M. labiatus by virtue of moderate inflation and closely spaced, prominent concentric undulations. All three species served at times as hosts for small epizoans, I. prefragilis for Exogyra aff. E. boveyensis, M. labiatus for Pycnodonte sp. A., and I. cuvieri and M. labiatus for Pseudoperna bentonensis and small cirripeds. Presence of these epizoans seems to have been linked to energy conditions on the sea floor because in beds lacking current-accumulated concentrations of skeletal debris epizoans are rare or lacking.
The Greenhorn section in Kansas contains many shaly chalk beds in which inoceramids are the only conspicuous bottom-dwelling macroinvertebrates and these may be abundant where no other benthonic species are preserved. Among species of the Greenhorn benthos only inoceramids could tolerate the lowest oxygenation levels reached on the Kansas floor of the Western Interior Sea.
Prev Page--Petrology || Next Page--Summary
Kansas Geological Survey, Geology
Placed on web June 5, 2010; originally published May 1975.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/209/07_paleo.html