![]()
![]()
![]()
Prev--Pre-Meeting Field Trip 1 || Next--Post-meeting Field Trip 3
1University of Iowa, 2University of Wisconsin-Milwaukee, 3Kansas State University, 4Kansas Geological Survey, 5University of Kansas
The morning will be devoted to observing plants and animals at Konza Prairie, one of the largest prairie preserves left in the United States. Gene Towne will discuss some of the current ecological projects at the site. Participants will observe and hear about the dynamics of prairie vegetation. including the effects of fire and bison. Bill Johnson will present results of recent isotopic research at Konza after a tour along the "Bison Loop." He will consider topographic variation in δ13C of modern soils as an aid to interpreting isotopic signals in buried soils, and biochemical and geochemical markers (e.g., 137Cs) as indicators of relative surface stability.
In the afternoon, we will drive northeast into Nebraska, where we will examine paleoecologic evidence for what the vegetation was like during the late Wisconsinan and Holocene. We will look at three large cutbanks on the South Fork of the Big Nemaha River in southeastern Nebraska. Detailed studies of the alluvial stratigraphy of the sites by Rolfe Mandel and Art Bettis have provided the stratigraphic and chronologic setting, and the paleoecology is based on studies by Dick Baker and Glen Fredlund. The Farwell locality has exposures of mostly Holocene-age alluvium, plus one mid-Wisconsinan outcrop, and has been analyzed for plant macrofossils, pollen, phytoliths, and stable carbon isotopes. The Old Bridge site has exposures of early Holocene alluvium overlain by late Holocene valley fill and has been analyzed for plant macrofossils, phytoliths, and pollen. The Miles Fan site is late Wisconsinan in age, and pollen and plant macrofossils have been studied. On our way back to Lawrence, we will stop briefly at Muscotah Marsh, studied by Johanna Gruger, 1973, and re-studied by Patrick Moss (unpublished data).
Kansas State University
The Konza Prairie Biological Station is a 3,487-ha tall grass prairie preserve located in the Flint Hills of northeastern Kansas (fig. 2-1). Relatively steep slopes and shallow, clayey soils overlying limestone and shale layers make the area more suitable for grazing than cultivation. Consequently, the area has a long history of fire and ungulate grazing. To achieve its primary mission of ecological research, Konza Prairie is divided into 52 watersheds that provide large, replicated experimental units subjected to various fire regimes. In addition, there is a resident bison herd of over 300 animals within a 992-ha enclosure. This tour will focus on changes in vegetation composition and structure resulting from different burn frequencies and different burn seasons, and the interactive effects of fire and large ungulate grazing. We will also talk about the bison herd, its management, and how the bison impact tallgrass prairie compared to cattle.
Figure 2-1--Map of Kansas showing the location of the Konza Prairie (map courtesy of The Nature Conservancy).
University of Kansas
The relatively undisturbed nature of the Konza Tallgrass Prairie Long Term Ecological Research (LTER) site provides a reference landscape for evaluation of biogeochemical signatures within grassland soils. Our first study is evaluating the radioisotope 137Cs as an erosion indicator in this loess-mantled, limestone terrain. Once the Cs inventory and its variability within the natural landscape at Konza have been characterized and modeled, we will apply the model to Fort Riley, an adjacent military reservation. in order to assess the rate of soil loss. While the climate and geology are identical to Konza, the reservation has experienced accelerated erosion caused by mechanized infantry. The second study is examining the vertical (soil profile) and horizontal (landscape) variability in stable C isotopes. Patterns observed are providing modern analog data that can be applied to the interpretation of horizontal and vertical isotopic data sequences from buried soils within the region.
A grid of sample points 50 m apart within a 660 m by 690 m area (fig. 2-2) was defined using a GIS, and specific points for core collection were selected with stratified random sampling. Sample points were then located on the ground using high-resolution GPS. Reference markers were placed, aerial photography flown, and a high-resolution DEM constructed (precision: 30 cm horizontal. 45 cm vertical). For Cs analysis, single cores, 30 cm long, were collected by hand at each of the 188 sample points. To evaluate micro-scale variability, 12 randomly selected points were sampled for a cluster of 12 additional samples each (fig. 2-2), thereby producing a total of 332 core samples. For stable C isotope analyses, soil surface samples (0-2 cm) and associated grass samples were collected at each of the 188 points. Also, representative upland sites were selected for extraction of 1-m to 2-m-deep soil cores to characterize the vertical distribution of stable C.
Figure 2-2--Landscape grid sampled for this study, including the scheme for micro-scale sampling at the 12 selected points.

The 332 30-cm-long cores were dried, ground, homogenized, and sent to J. Kaste and A. Heimsath at Dartmouth College for 137Cs and 239Pu assays. Sub-samples were also analyzed for elemental concentrations, including 133Cs using an ICP-MS. Other analyses of the short cores include clay mineralogy (XRD), particle size (laser diffraction), carbon content (LECO CN analyzer), and rock magnetic analysis. Soil and grass samples were assayed for carbon isotopic content using a system consisting of a CE elemental analyzer, GasBench II, Conflo II interface, CombiPal autosampler, and ThermoFinnigan Delta Plus mass spectrometer.
The theoretical basis of using 137Cs as an indicator of soil-surface stability is that H-bomb testing created a 137Cs reservoir in the atmosphere, which is delivered to the soil in precipitation. 137Cs then attaches to soil particles and concentrates within the uppermost part of the soil, with smaller amounts being transported down the soil profile. Generally, the more 137Cs in the upper soil profile, the less erosion that has occurred since bomb testing began in the 1950's. The two research questions focus on defining (1) mechanisms that 137Cs uses to move through soil and (2) scale of variation in 137Cs soil inventories within a natural landscape.
137Cs is thought to be strongly adsorbed onto clay minerals and organic matter in soil, resulting in very low mobility (Ritchie and McHenry, 1990), and the small size of 137Cs plumes emanating from mining and milling waste piles or other low-level radioactive waste discharges have been attributed to nearly irreversible sorption of 137Cs onto the clay fraction in soils (Brady et al., 2002). Sorption onto clays may occur onto high-affinity or low-affinity sites on illite (Poinssot et al., 1999), creating sub-reservoirs of what may be lower-mobility and higher-mobility 137Cs. Presence of other clay minerals with higher ion-exchange coefficients, pH variations, or organic matter could lead to higher mobility (Chappell, 1999), as could changes in the redox state of the soil (Thomson et al., 2002). In addition, because ion exchange is competitive, a change in the dominant cation (especially an increase in the dissolved Na content) has the potential to mobilize sorbed 137Cs (Poinssot et al., 1999). The distribution coefficient for 137Cs is apparently not strongly affected by the amount of humic acid present, suggesting that complexation by dissolved organic carbon is not an important transport mechanism (Langmuir, 1997). Previous work delineating potentially mobile reservoirs of 137Cs have suggested that Chernobyl-sourced 137Cs is probably affiliated minimally with clay minerals in exchangeable sites, and more with organic matter (or possibly fuel particles that liberate 137Cs during the step that liberates material associated with oxidizable phases), and with phases dissolved using 7 M nitric acid at 80°C (Salbu et al., 1994).
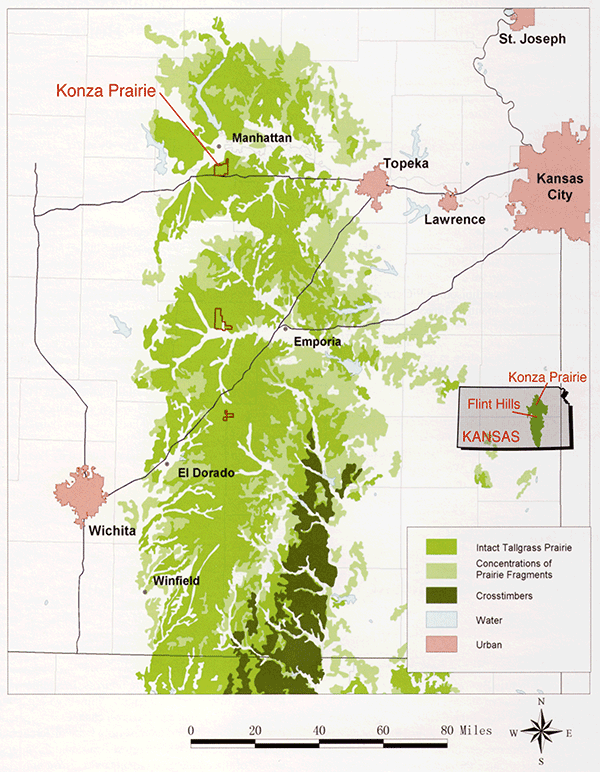
At Konza, 137Cs (half life of 30.2 years) and 239Pu (half-life of 24,000 years) radioisotope activities show similar exponential declines with depth (fig. 2-3), which supports the idea that the decrease in the activity of 137Cs with depth is not a function of its short half life. Preliminary findings suggest that Cs at Konza (stable Cs, mass 133, and, by proxy, radioactive Cs, mass 137) has an affinity for SOC. Most (95%) of the stable Cs resides in the weathering-resistant portion of the soil, as isolated using a sequential-extraction technique modified from Tessier et al. (1979). Most of the remaining mobile stable Cs is affiliated with oxidizable phases, probably mostly SOC (fig. 2-4). Enrichment in Cs concentration in SOC with depth occurs, as demonstrated by normalizing Cs concentration to Cs concentration in the oxidizable fraction of the soil and to the shallowest sample (fig. 2-4). Cs content in the other extractable reservoirs (reducible, acid-soluble, and easily exchangeable fractions) shows much less change with depth (fig. 2-4). An exponential decline in 137Cs and SOC with depth contrasts with an exponential increase with depth of stable Cs affiliated with oxidizable phases (SOC).
Figure 2-3--137Cs and 239Pu radioisotope distributions in a soil profile from the upland of the landscape grid.
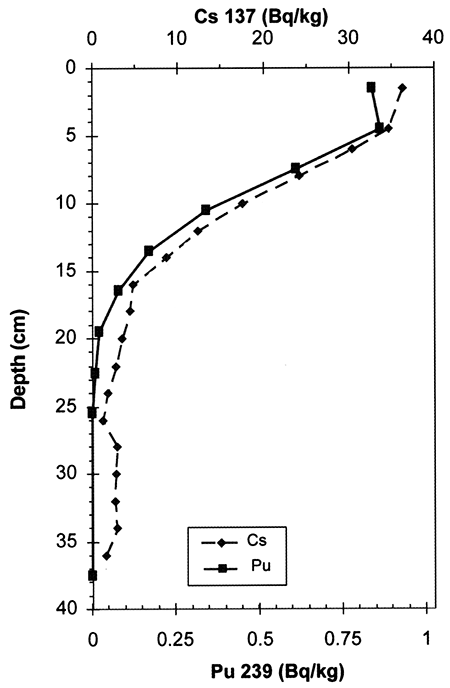
Figure 2-4--Cs distribution with depth. A) Cs concentration with depth; B) Cs content normalized to shallowest sample and to the extract reservoir (% SOC for oxidizable soil components; % clay for exchangeable Cs); C) log-normal distributions of 137Cs activity normalized to shallowest sample, SOC normalized to shallowest sample, and stable Cs concentration normalized to shallowest sample and to SOC.
Carbon consists of two naturally occurring stable isotopes, 12C (98.89%) and 13C (1.1 %), which are usually expressed in ratio form (13C/12C) or as δ13C (‰). Plants discriminate against 13C during photosynthesis such that the tissue is depleted in 13C relative to the atmosphere (O'Leary, 1981). Grasses fix carbon from atmospheric CO2by one of three pathways, the Calvin-Benson (C3), Hatch-Slack (C4), and Crassulacean acid metabolism (CAM). Because CAM is employed by desert plants, it is not widespread in the Great Plains. Each of these photosynthetic pathways results in differing levels of discrimination against 13C (Smith and Epstein, 1971). Values of δ13C from C4 plants produce a range from about -17&permill; to -10&permill; with an average of -13&permill; whereas C3 plants range from -32&permill; to -20&permill; and average -27&permill; (Ceding et al., 1989; Boutton, 1991). SOC represents an integration of the isotopic inputs from the various sources within a plant community (Troughton et al., 1974; Balesdent and Mariotti, 1996), the value of which approximates the photosynthetic pathway of the dominant species (Stout and Rafter, 1978; Nadelhoffer and Fry, 1988). After removal of carbonates using IN HCl (16 hours @ room temperature), samples were assayed for stable carbon and carbon content using a ThermoFinnigan Delta Plus mass spectrometer and elemental analyzer system.
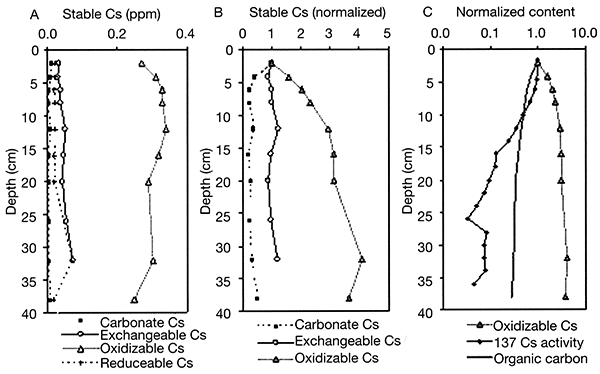
Konza Tallgrass Prairie is dominated by C4grasses, which translate their characteristic isotopic signal to the soil. Soil surface samples from the landscape grid averaged -17.5&permill;, None of the variance yet appears to be explained, however, by slope and aspect. Senescent grass samples averaged -15.8&permill;, reflecting a clear C4signal. The isotopic profile of an upland soil indicates surface values of about -18&permill;, but with an increase to about -14&permill; at 10-20-cm depth (fig. 2-5). Such 13C depletion in modern surface soils has been widely recognized (e.g., Kelly et al., 1991; de Freitas et al., 2001; Torn et al., 2002) and is due in part to historic, fossil-fuel-induced 13C depletion of the CO2in the atmosphere (Boutton, 1996). Consequently, the larger value at 10-20 cm is more representative of the long-term average of the stable grassland vegetation. Likewise, isotopic depletion of soil-surface SOC within the landscape grid likely explains the disparity between the mean of the soil surface and grass sample δ13C. Isotopic values decrease downward in the core to a low of about -24.0&permill; (circa 1.6 m), reflecting cool-season C3plant domination of the late-Wisconsinan central Great Plains environment. Increase in δ13C values at the core base reflects presence of a middle-Wisconsinan buried soil, the Gilman Canyon Formation soil, one characterized regionally by its C4grass-dominated environment (Johnson and Willey, 2000).
Figure 2-5--Distribution of δ13C and SOC within an upland soil profile.
Balesdent, J., and Mariotti, A., 1996, Measurement of soil organic matter turnover using 13C natural abundance; in, Mass Spectrometry of Soils, T. W. Boutton and S. Yamasaki, eds.: New York, Marcel Dekker, Inc., p. 83-111.
Boutton, T. W., 1991, Stable carbon isotope ratios of natural materials--I. Sample preparation and mass spectrometric analysis; in, Carbon Isotope Techniques, D. C. Coleman and B. Fry, eds.: New York, Academic Press, p. 155-171.
Boutton, T. W., 1996, Stable carbon isotope ratios of soil organic matter and their use as indicators of vegetation and climate change; in, Mass Spectrometry of Soils, T. W. Boutton and W. Yamasaki, eds.: New York, Marcel Dekker, Inc., p. 47-82.
Brady, P. V., Jove-Colon, C. F., Carr, G., and Huang, F., 2002, Soil radionuclide plumes; in, Geochemistry of Soil Nuclides, P.-C. Zhang and P. V. Brady, eds.: Soil Science Society of America, Special Publication No. 59, p. 165-190.
Cerling, T. E., Quade, J., Wang, Y., and Bowman, J. R., 1989, Carbon isotopes in soils and paleosols as ecology and paleoecology indicators: Nature, v. 341, p. 138-139.
Chappell, Adrian, 1999, The limitations of using 137Cs for estimating soil redistribution in semi-arid environments: Geomorphology, v. 29, p. 135-152.
de Freitas, H. A., Pessenda, L. C. R., Aravena, R., Gouveia, S. E. M., and Ribeiro, A. D. S., 2001, Late Quaternary vegetation dynamics in the southern Amazon basin inferred from carbon isotopes in soil organic matter: Quaternary Research, v. 55, p. 39-46.
Johnson, W. C., and Willey, K. L., 2000, Isotopic and rock magnetic expression of environmental change at the Pleistocene-Holocene transition in the central Great Plains: Quaternary International, v. 67, p. 89-106.
Kelly, E. F., Amundson, R. G., Marino, B. D., and DeNiro, M. J., 1991, Stable carbon isotopic composition of carbonate in Holocene grassland soils: Soil Science Society of America Journal, v. 55, p. 1,651-1,658.
Langmuir, Donald, 1997, Aqueous environmental chemistry: Upper Saddle River, New Jersey, Prentice-Hall, Inc., p. 539.
Nadelhoffer, K. J., and Fry, B., 1988, Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter: Soil Science Society of America Journal, v. 52, p. 1,633-1,640.
O'Leary, M., 1981, Carbon isotope fractionation in plants: Phytochemistry, v. 20, p. 553-567.
Poinssot, C., Baeyens, B., and Bradbury, M. H., 1999, Experimental and modeling studies of caesium sorption on illite: Geochimica et Cosmochimica Acta, v. 63, no. 19/20, p. 3,217-3,227.
Ritchie, J. C., and McHenry, J. R., 1990, Application of radioactive fallout cesium-137 for measuring soil erosion and sediment accumulation rates and patterns--A review: Journal of Environmental Quality, v. 19, p. 215-233.
Salbu, B., Oughton, D. H., Ratnikov, A. V., Zhigareva, T. L., Kruglov, S. V., Petrov, K. V., Grebenshakikova, N. V., Firsakova, S. K., Astasheva, N. P., Loshchilov, N. A., Hove, K., and Strand, P., 1994, The mobility of 137Cs and 90Sr in agricultural soils in the Ukraine, Belarus, and Russia, 1991: Health Physics, v. 67, no. 5, p. 518-528.
Smith, B. N., and Epstein, S., 1971, Two categories of 13C/12C ratios for higher plants: Plant Physiology, v. 47, p. 380-384.
Stout, J. D., and Rafter, T. A., 1978, The 13C/12C isotopic ratios of some New Zealand tussock grassland soils; in, Stable Isotopes in the Earth Sciences, B. W. Robinson, ed.: Wellington, New Zealand Department of Scientific and Industrial Research, p. 75-83.
Tessier, A., Campbell, P. G. C., and Bisson, M., 1979, Sequential extraction procedure for the speciation of particulate trace metals: Analytical Chemistry, v. 51, no. 7, June 1979, p. 844-851.
Thomson, J., Dyer, F. M., and Croudace, I. W., 2002, Records of radionuclide deposition in two salt marshes in the United Kingdom with contrasting redox and accumulation conditions: Geochimica et Cosmochimica Acta, v. 66, no. 6, p. 1,011-1,On
Tom, M. S., Lapenis, A. G., Timofeev, A., Fischer, M. L., Babikov, B. V., and Harden, J. W., 2002, Organic carbon and carbon isotopes in modem and l00-year-old soil archives of the Russian steppe: Global Change Biology, v. 8, p. 941-953.
Troughton, J. H., Stout, J. D., and Rafter, T. A., 1974, Longterm stability of plant communities: Carnegie Institute, Washington Yearbook, v. 73, p. 838-845.
1University of Iowa, 2University of Winconsin-Milwaukee, 3Kansas Geological Survey, 4University of Queensland
The paleoecological record of much of North America is largely based on studies of fossil pollen from lakes and wetlands. Such depositional basins are widespread, especially in glaciated regions, but pollen sites are much less common on the Great Plains beyond the Wisconsinan glacial margin. Of 328 pollen sites listed for eastern North America (Webb et al., 1993), only one (Swan Lake in the Nebraska Sandhi lis) is shown for the region from Nebraska to southern Oklahoma. In these regions, other types of paleoecological information have been developed from different depositional environments. Plant macrofossils, where present, can yield species-level identifications and definite local presence of those plants. Phytoliths provide information of useful subgroups of grasses that are indistinguishable from pollen, and δ13C analyses can be transformed into percentages of C4 vs. C3 plants. Both these techniques can be applied to either soils or alluvial sediments, and both can be helpful in distinguishing prairie from forest and distinguishing among various types of prairie. The δ13C can also be analyzed from cave speleothems, if caves are nearby, giving very high time-resolution. This trip provides the opportunity to examine the potential of stream cutbanks as sources of these paleoecological data and to discuss how these recent paleoecological tools correlate with one another.
The purpose of the afternoon portion of the trip is to show you two sections near Du Bois, Nebraska, that we have studied and to discuss the results. The deposits were sampled from cutbanks along the river. We will examine the lithologic units, look at the fossiliferous deposits, screen them to show you examples of the macrofossils, and discuss their interpretation. At first glance, these deposits would seem to be of questionable use, if not downright useless. They are generally discontinuous spatially, separated by sediments barren of fossils; they are discontinuous snapshots in time; and they would seem particularly prone to reworking. However, by choosing the right sample sites, these problems can largely be overcome, and the benefits can be substantial. First, while it is true that most of the sediments are barren of plant macrofossils, if the creek is permanent and not in a stretch that loses water (in karst terrain, for example), organic-rich deposits near or even below creek level may be present. Second, though the deposits generally do represent a very short time interval, in the streams we have worked on, different ages are exposed in the stream's cutbanks over distances of tens of meters to a few kilometers upstream and downstream. We have put together a series of snapshots representing much of the Holocene. Third, it is true that reworking is possible, but with careful work and site selection, we have determined that there is no evidence of wholesale reworking, and only a few cases of reworking of individual plant remains were recorded. Preservation of very delicate plant remains effectively rules out reworking in many cases. One fairly common exception is spruce needles, which are extremely durable. We occasionally do find needles in middle Holocene deposits, but they are broken and corroded, in the midst of other well-preserved delicate plant macrofossils. Furthermore, we try to minimize reworking by sampling fairly close to the headwaters of the stream; thus, if reworking did occur, it would not involve transport from a distant biome.
There are several advantages to using these organic-rich alluvial deposits. They do contain pollen (Chumbley et al., 1990), and comparisons with the closest lake deposits indicate that the alluvial pollen signal correlates well with the lacustrine record (Baker et al., 1992). A big advantage is that the alluvial deposits contain a much more diverse plant macrofossil record than is found in lacustrine and wetland sediments (Baker et al., 1996; Baker et al., 2002). These fossils represent the whole range of habitats in the drainage basin, including upland, floodplain, weedy disturbed habitats, wetlands that occur on the floodplain, and in the stream (fossils of aquatic plants) (Baker, 2000; Baker et al., 1996; Baker et al., 2000; Baker et al., 2002). In many cases there are well over 200 taxa, and the preservation is often excellent, allowing specific identification. Wood or other upland macrofossils are dated from each section to provide the chronology. Thus, these deposits give a picture of the landscape that has very high ecological resolution. The other big advantage is that the phytolith and isotopic records test and refine the pollen and plant macrofossil interpretations. The climatic record derived from these data is then compared with the stream activity derived from the alluvial stratigraphy and lithology (see Mandel, 1996; Mandel and Bettis, 2001).
On the way back to Lawrence, we will make a short stop at Muscotah Marsh, an important locality cored and analyzed in the early 1970's by Gruger (1973). We will compare and contrast the results from the study sites in the South Fork of the Big Nemaha River with the paleobotanical record at Muscotah.
We collected ca. 1-liter samples in resealable plastic bags from several organic-rich zones near the base of three large cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska. These were subsampled by Baker for plant macrofossils and by Fredlund for phytoliths, pollen, and stable carbon isotopes. Samples were prepared for phytolith analysis by (1) removal of carbonates with HCl, (2) removal of clays with 0.1 N sodium pyrophosphate, (3) oxidation of the residue with 30% hydrogen peroxide, and (4) isolation of biogenic silicates using heavy-liquid (ZnBr2 at 2.35 specific gravity) as described in Fredlund and Tieszen (1997). Pollen samples were processed using heavy liquid flotation (Johnson and Fredlund, 1985). Pollen and phytoliths were identified using reference materials at the Geography Department, University of Wisconsin-Milwaukee. Plant macrofossils were washed through 0.5- and O.1-mm sieves, picked by hand, and identified using the reference collection at the Geoscience Department, University of Iowa.
The stable carbon isotope analyses of bulk organic matter were carried out at the Isotope Ratio Mass Spectrometer facility in the Biology Department at Augustana College, Sioux Falls, South Dakota, as described in Fredlund and Tieszen (1997). Pretreated samples were combusted in a Carlo Erba CHN analyzer coupled to an SIRA-10 Isotope Ratio Mass Spectrometer fitted with a special triple trap to isolate cryogenically and purify the CO2. Lab standard and random replicates were run to ensure precision better than 0.2&permill;, Isotopic ratios are expressed as per-mil deviations from the PDB standard.
This site (Stop 1 in fig. 2-6) is an entrenched channel on the South Fork of the Big Nemaha River, about 1.5 km east of the town of Du Bois (ca. 40° 01' 15" N. Lat., ca. 96° 01' 15" W. Long.). It is within about 20 km of its source in northeastern Kansas. At this stop, we will park on a bridge and view Holocene and mid-Wisconsinan alluvial sediments exposed in numerous cutbanks along a stretch of the river (Mandel, 1996; Mandel and Bettis, 2001). The Holocene sediments are contained in four members of the DeForest Formation: Gunder (oldest), Roberts Creek, Honey Creek. and Camp Creek (fig. 2-7). The DeForest Formation is a lithostratigraphic unit containing all the fine-grained Holocene alluvium in Iowa (Bettis, 1995). Recent studies have extended the DeForest Formation into eastern Nebraska (Dillon. 1992; Mandel, 1994; Mandel and Bettis, 1995, 2001). At Farwell, the DeForest Formation is laterally inset against the Severance Formation (fig. 2-8). The Severance formation consists of oxidized, sandy alluvium and is Wisconsinan in age (Mandel and Bettis, 2003). The DeForest and Severance Formations are distinguished on the basis of their lithology and geomorphic position, and their chronology has been determined by radiocarbon dating. Plant macrofossils associated with the alluvial fills typically occur at or near water level.
Figure 2-6--Map of field-trip stops.
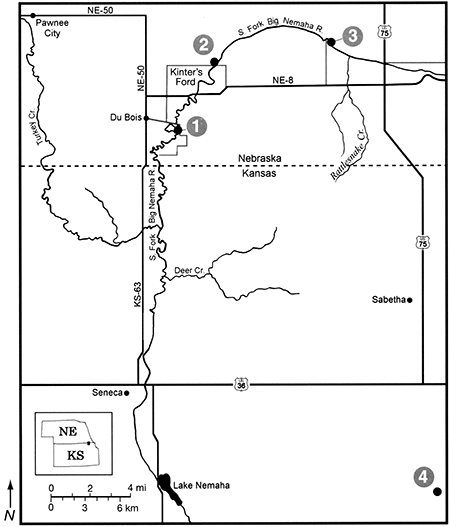
Figure 2-7--Stratigraphic and temporal relationships of Wisconsinan formations and units of the Holocene DeForest Formation along the South Fork Big Nemaha River (from Mandel and Bettis, 2001).
The vegetation of this part of its drainage basin was tall grass prairie prior to European settlement, with some riparian forest along the stream. Although the prairie has largely been lost to cultivation, its composition was probably typical diverse prairie vegetation somewhat similar to what we saw at Konza Prairie. The riparian forest that remains includes Acer negundo (boxelder), Acer saccharinum (silver maple), Fraxinus pennsylvanica (green ash), Populus deltoides (cottonwood), Salix spp. (willow), and Ulmus americana (American elm).
Pollen sites from central Nebraska (Wright et al., 1985) and the northern Great Plains (Barnosky et al., 1987; Grimm, 2001) indicate that prairie was present throughout the Holocene, but that there were significant fluctuations. In the early Holocene, open deciduous forest on the northeastern Great Plains gave way to prairie, and there is strong evidence of recurring drought in the mid-Holocene (Grimm, 2001; Laird et al., 1998). The only pollen site on the eastern edge of the central Great Plains is Muscotah Marsh (Gruger, 1973), our last stop, where the Holocene was compressed and not well dated.
Our work in the South Fork of the Big Nemaha River valley provides pollen, plant macrofossil, phytolith, and carbon-isotopic data to reconstruct the Holocene vegetation. Because of the coarse time resolution, we cannot resolve short-term climatic changes, but the cutbank sediments give a far better ecological resolution of a snapshot of the past vegetation than marsh or lake sediment samples.
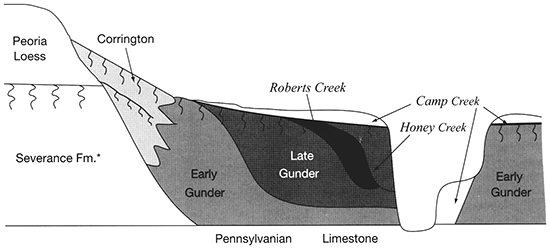
The Farwell site (fig. 2-8) is part of a sequence of sites along the South Fork of the Nemaha River that was analyzed for the plant macrofossil dataset. Uncalibrated radiocarbon ages for these sites are shown in table 2-1. Figure 2-9 shows the pollen percentages from these sites, and figs. 2-10 through 2-12 show macrofossils of upland trees, riparian trees, prairies, and selected weeds. Macrofossils representing aquatic, wetland, other weed, forest floor herb, and shrub and vine habitats are plotted and discussed in Baker (2000). Figure 2-13 presents the phytolith and δ13C data. All are arranged vertically by radiocarbon age of each site, and the macrofossils are grouped by plant habitat and physiognomy. Five zones are recognized, mainly on the basis of macrofossils; all ages are expressed in uncalibrated radiocarbon years. The basal zone (Zone 1, 9000-8500 yrs B.P.) has a diverse mix of macrofossils of deciduous tree taxa including hickories, white ash, butternut type, walnut, ironwood, and bur oak. Riparian trees including boxelder, elms, willows, and mulberry. Despite the diverse mix of forest trees, prairie also is well represented, with the major tall prairie grasses and important legumes, as well as several other taxa. Pollen of deciduous trees is relatively low, and typical prairie pollen types including grass, chenopods, and ragweed are all high. The δ13C values are highly negative, but they increase upwards, and the highest percentage of C3 phytoliths occurs in this zone, although both C3 and C4 types are present.
Figure 2-8--View of the cutbank at the Farwell site (Stop 1). Radiocarbon ages were determined on plant macrofossils. The white line marks the boundary between the DeForest and Severance Formations.

Figure 2-9--Pollen spectra for sediment samples collected from cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska. The pollen analysis was conducted by Glen Fredlund.
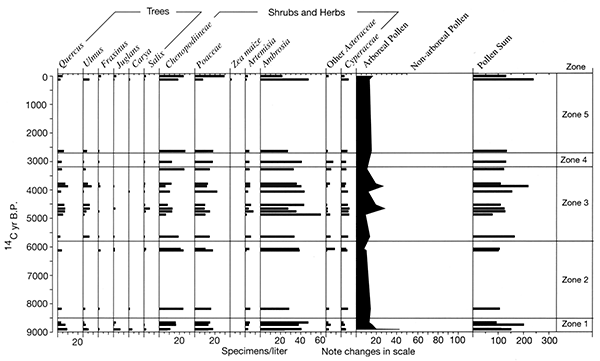
Figure 2-10--Summary of plant macrofossils derived from trees. The macrofossils were recovered from Holocene sediments exposed in cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska.
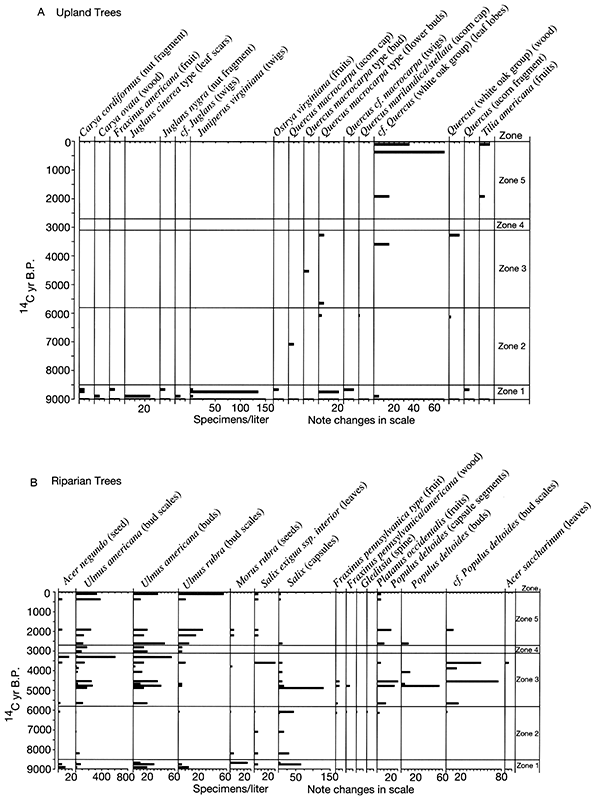
Figure 2-11--Summary of plant macrofossils derived from prairie species. The macrofossils were recovered from Holocene sediments exposed in cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska.
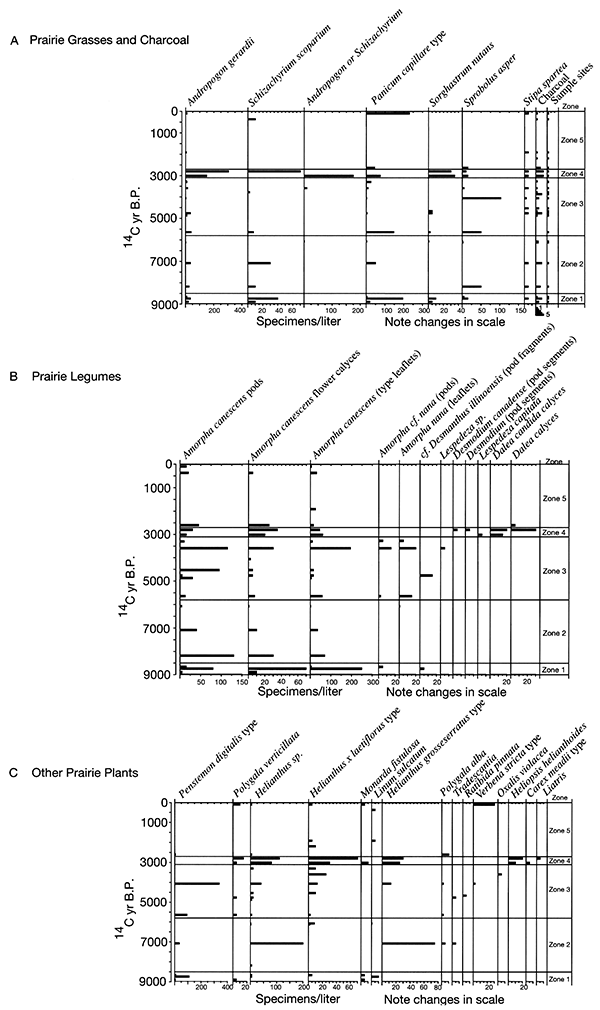
Figure 2-12--Summary of plant macrofossils derived from weeds. The macrofossils were recovered from Holocene sediments exposed in cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska.
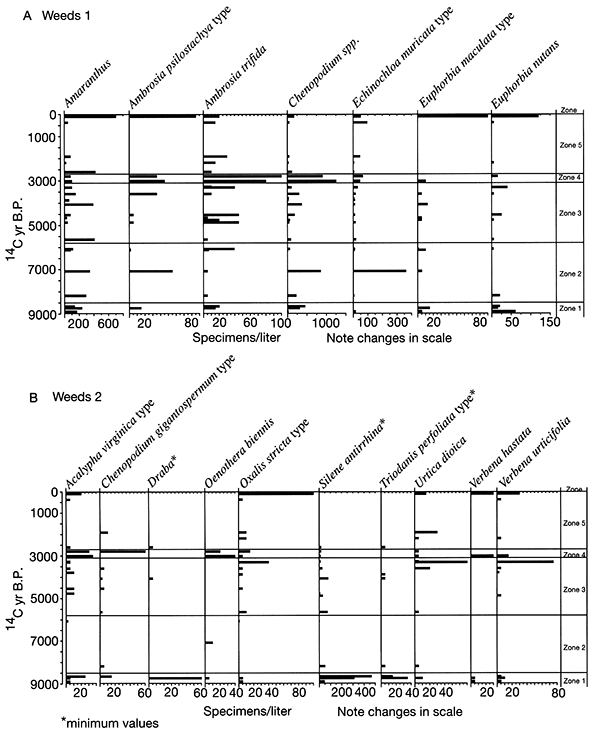
Figure 2-13--Phytolith and stable carbon isotope records for sediment samples collected from cutbanks along the South Fork of the Big Nemaha River near Du Bois, Nebraska.
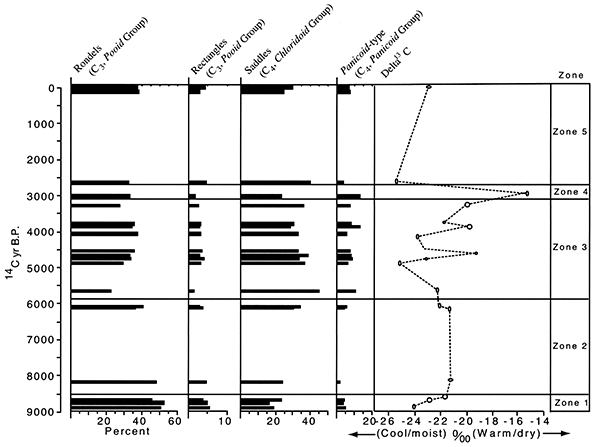
Table 2-1. Radiocarbon ages for study sites in the South Fork Big Nemaha River valley (from Mandel and Bettis, 2001).
| Stop No. and Locality |
Stratigraphic Unit |
Material Assayed |
Depth (m) |
Uncorrected Age (yrs B.P.) |
δ13C | Corrected Age (yrs B.P.) |
Lab No. |
|---|---|---|---|---|---|---|---|
| STOP 1, Farwell | |||||||
| 25PW63 | Late Gunder Mbr. | Wood | 6.50-6.70 | NR | -28.0 | 4060±70 | ISGS-3416 |
| 25PW63 | Late Gunder Mbr. | Wood | 7.30-7.40 | 4,460±60 | -27.5 | 4420±60 | Tx-9017 |
| 25PW63 | Late Gunder Mbr. | Charcoal | 3.23-3.28 | 3.150±60 | -25.1 | 3140±60 | Tx-9015 |
| 25PW63 | Severance Fm. | Plant frag. | 8.72-8.73 | NR | NR | 37.100±520 | CAMS-41681 |
| 25PW63 | Severance Fm. | Charcoal | 8.81-882 | NR | NR | 53,110±2870ˆ | CAMS-33983 |
| Floodplain- | Camp Creek Mbr. | Wood | 3.75-3.90 | NR | -28.6 | Modern | ISGS-3415 |
| 25PW64 | Early Gunder Mbr. | Charcoal | 1.30-1.70 | 4,460±70 | -26.0 | 4440±70 | Tx-8952 |
| 25PW64 | Early Gunder Mbr. | Charcoal | 1.30-1.70 | 3.920±70 | -25.0 | 3920±70 | Beta-95298 |
| 25PW64 | Late Gunder Mbr. | Wood | 7.69-7.90 | NR | -27.9 | 4660±70 | ISGS-3417 |
| 25PW64 | Late Gunder Mbr. | Wood | 7.75-7.98 | NR | -27.6 | 4530±70 | ISGS-3418 |
| 25PW65 | Late Gunder Mbr. | Wood | 8.14-8.30 | NR | -27.0 | 4760±70 | ISGS-3414 |
| 25PW65 | Late Gunder Mbr. | Charcoal | 1.60-1. 75 | 3,670±180 | -26.1 | 3650±180 | Tx-9016 |
| 25PW65 | Honey Creek Mbr. | Charcoal | 4.60-4.75 | 1,880±40 | -26.8 | 1850±40 | Tx-9013 |
| 25PW65 | Severance Fm. | Soil | 2.00-2.10 | NR | -23.0 | 15,110±110 | ISGS-4468 |
| STOP 2 | |||||||
| Old Bridge | Early Gunder Mbr. | Wood | 12.11-12.31 | NR* | -27.7 | 8890±70 | ISGS-3427 |
| Old Bridge | Early Gunder Mbr. | Wood | 12.11-12.25 | NR | -26.7 | 8890±70 | ISGS-3426 |
| Old Bridge | Early Gunder Mbr. | Wood | 12.59-12.70 | NR | -25.6 | 8740±70 | ISGS-3430 |
| Old Bridge | Early Gunder Mbr. | Wood | 12.05-12.27 | NR | -26.9 | 8630±70 | ISGS-3436 |
| Old Bridge | Early Gunder Mbr. | Wood | 11.72-11.93 | NR | -28.2 | 8180±70 | ISGS-343I |
| Old Bridge | Honey Creek Mbr. | Wood | 9.22-9.33 | NR | -28.0 | 3870±70 | ISGS-3429 |
| Old Bridge | Honey Creek Mbr. | Wood | 10.78-10.95 | NR | -27.0 | 2630±70 | ISGS-3437 |
| Old Bridge | Honey Creek Mbr. | Wood | 8.55-8.66 | NR | -26.6 | 1910±70 | ISGS-3424 |
| STOP 3 | |||||||
| Miles Fan | Corrington Mbr. | Soil | 1.30-1.40 | 2,920±60 | -12.9 | 3120±50 | Tx-8945 |
| Miles Fan | Corrington Mbr, | Soil | 3.60-3.70 | 10,250±120 | -12.9 | 10,450±120 | Tx-8944 |
| Miles Fan | Peat | 9.90-10.0 | 23,530±310 | -27.0 | 23,490±310 | Tx-8946 | |
| Miles Fan | Peat | 10.3-10.4 | 24.690±410 | -27.3 | 24,640±410 | Tx-8947 | |
| Miles Fan | Peat | 10.6-10.7 | 27.620±550 | -27.4 | 27.580±550 | Tx-8948 | |
| Miles Fan** | Wood | ca. 10.0 | NR | -25.1 | 19,920±240 | ISGS-468I | |
| Miles Fan** | Wood | ca. 10.0 | NR | -24.5 | 20,290±120 | ISGS-4680 | |
| Other Localities | |||||||
| DuBois Quarry | Severance Fm. | Soil | 2.36-2.46 | 18.780±140 | -22.4 | 18,830±140 | Tx-9308 |
| DuBois Quarry | Severance Fm. | Soil | 4.16-4.26 | 25,730±360 | -28.5 | 25,670±360 | Tx-9309 |
| DuBois Quarry | Severance Fm. | Wood | 7.62-7.82 | NR | NR | 33,257±1096 | ISGS A-0020 |
| Kinter's Ford | Early Gunder Mbr. | Soil | 2.90-3.00 | 6,930±70 | -16.5 | 7070±70 | Tx-8943 |
| Kinter's Ford | Early Gunder Mbr. | Wood | 7.05-7.20 | NR | -28.6 | 6120±70 | ISGS-3405 |
| Kinter's Ford | Early Gunder Mbr, | Wood | 7.15-7.38 | NR | -28.5 | 6070±70 | ISGS-3400 |
| Kinter's Ford | Early Gunder Mbr. | Wood | 6.70-6.90 | NR | -28.3 | 5640±70 | ISGS-3401 |
| Kinter's Ford | Early Gunder Mbr. | Wood | 4.44-4.60 | NR | -27.0 | 4870±70 | ISGS-3398 |
| Kinter's Ford | Early Gunder Mbr. | Soil | 1.80-1.90 | 4.630±110 | -13.3 | 4780±110 | Tx-8942 |
| Kinter's Ford | Honey Creek Mbr. | Wood | 5.22-5.44 | NR | -27.3 | 3780±70 | ISGS-3402 |
| Kinter's Ford | Honey Creek Mbr. | Wood | 6.48-6.80 | NR | -26.4 | 3300±70 | ISGS-3446 |
| Kinter's Ford | Roberts Creek Mbr. | Wood | 3.49-3.70 | NR | -27.5 | 3270±70 | ISGS-3399 |
| Kinter's Ford | Honey Creek Mbr. | Wood | 6.56-6.70 | NR | -28.3 | 3010±70 | ISGS-3404 |
| Kinter's Ford | Honey Creek Mbr. | Wood | 6.53-6.64 | NR | -25.9 | 2800±70 | ISGS-3403 |
| Kinter's Ford | Honey Creek Mbr. | Wood | 7.23-7.55 | NR | -26.4 | 2190±70 | ISGS-3452 |
| Kinter's Ford | Camp Creek Mbr. | Wood | 6.56-6.79 | NR | -27.5 | 370±70 | ISGS-3453 |
| 25PW83 | Late Gunder Mbr. | Wood | 8.30-8.50 | NR | -26.4 | 3590±70 | ISGS-3450 |
| 25PW83 | Late Gunder Mbr. | Charcoal | 200-205 | 2.290±70 | -26.2 | 2270±70 | Tx-9014 |
| *NR = Not Reported. ˆAge rejected. **Sample was collected ca. 100 m downstream from the Miles Fan. ~Sample was collected on the west side of the South Fork of the Big Nemaha River, immediately west of site 25PW63. |
|||||||
The pollen record for Zone 1 suggests that prairie was prominent on flat uplands, but the combined records indicate that a mesic forest was still widespread on valley walls, in ravines, and probably on the floodplain. Riparian elements would likely be confined to areas close to the river. The macrofossils indicate that C4 grasses are abundant in this zone (fig. 2-11), yet the strong presence of C3 grasses is indicated by phytoliths (fig. 2-13), and the δ13C values indicate mixed C3 and C4 grasses (fig. 2-13).
Only a few sites represent Zone 2; nevertheless, a noteworthy change from Zone 1 is apparent. Macrofossils of most upland and riparian trees have disappeared. Arboreal pollen also declined. Elements of prairie continue and some weeds increase in abundance. The δ13C values increase about 2 per mil and remain steady. Phytoliths of C4 grasses increase at the top of this zone.
These changes suggest a distinctly drier climate between 8500 and 5800 yrs B.P. Studies of the 1930's droughts show that the prairie vegetation became much less dense and more patchy, giving room for weeds to become established (Weaver, 1968; Tomenek and Hulett, 1970). Streams were generally undergoing aggradation early in this zone, becoming stable towards the end, and alluvial fans were aggrading throughout Zone 2.
A major change in Zone 3 (5800-3100 yrs B.P.) is that there is a marked increase in the abundance of riparian trees, including boxelder, elm, willow, green ash, and cottonwood. Pollen and plant macrofossils of prairie and weedy plants continue to be present in Zone 3, and only oak was present as a macrofossil. Phytoliths of C4 plants are slightly more prominent than in Zone 2, and the δ13C values fluctuate significantly during this interval.
Zone 4 (3100-2700 yrs B.P.) marks another distinct interval. Upland trees remain absent, and all riparian trees disappear except elm. In contrast, most other elements increase strikingly. These include many prairie, weed, wetland, and aquatic species. Charcoal is abundant in this zone, and δ13C values are at their peak, indicating an abundance of C4 grasses. Phytoliths of the chloridoid group of C4 plants (which include grama grasses and buffalo grass of the midgrass-shortgrass prairie) decline, but those of panicoid C4 plants increase.
The increased abundance of many macrofossil taxa and the increase in charcoal suggests that fires were more common at ca. 3100-2700 yrs B.P., allowing more runoff and increased deposition of plant remains. The disappearance of all riparian elements except elm, which is a late-successional tree, suggests that climatological and hydrological factors prevented the establishment of early succession on sandbars of cottonwood and willow for a few hundred years. One hypothesis is that flow was generally low in spring and early summer, allowing spring establishment on sandbars. Occasional large floods may have occurred in Zone 3, providing fresh sand forto colonize reworked sandbars. In Zone 4, floods may have been smaller and droughts more frequent, favoring late-successional elms. The absence of new sandbars would make it more difficult for willow and cottonwood to germinate. The loss of most tree species and the increase in diversity and abundance of prairie plants indicate a warmer and/or drier climate at this time. Valley-floor surfaces were stable at this time, but alluvial fans were aggrading. willow, cottonwood, and other early successional plants
The return of oak and basswood, along with forest-floor herbs and riparian trees, and the decline of prairie and weed species occurs in Zone 5 (2700 yrs B.P.-present). Values of δ13C decline sharply at the base of this zone, but no other analyses in this interval were done except for a post-settlement sample. Chloridoid-group phytoliths increased and panicoid decreased in this basal sample. These data suggest that climate had become cool and moist enough to support some trees that provide enough shade for some forest-floor herbs.
The Old Bridge Site is about 8 km downstream from Stop 1 (Stop 2 in fig. 2-6). A 400-m-long cutbank on the east side of the South Fork of the Big Nemaha River exposes a complex sequence of alluvial deposits beneath the T-1 terrace (fig. 2-14). Three members of the DeForest Formation are represented at this locality: Gunder, Honey Creek, and Camp Creek (fig. 2-15).
Figure 2-14--View of the cutbank at the Old Bridge site.

Figure 2-15--Outcrop along the east bank of the South Fork of the Big Nemaha River (Stop 2).
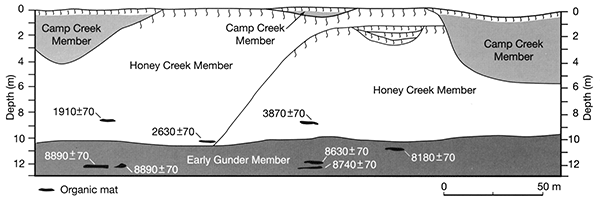
The Old Bridge Site displays important early Holocene and late Holocene sections (ca. 8000-9000 and ca. 3900-1900 uncalibrated 14C yrs B.P.) that contribute to our overall paleoecological reconstruction along the South Fork of the Nemaha River. These sections have been analyzed for pollen, phytoliths, stable carbon isotopes, and a diverse macrofossil assemblage that reflects what was growing on the landscape. Other sections nearby fill in some of the gaps in time. Table 2-2 shows the variety of plant parts that have been identified just from the Old Bridge site, including buds, bud scales, leaf fragments, leaflets, nut fragments, wood (identified by Laura Strickland), seeds, and fruits. Most samples contain between 200 and 600 macrofossil types picked from just 200 ml of sediment.
Table 2-2--Plant macrofossils collected from the Old Bridge site (Stop 2). The data are presented by habitat.
| site # / info. date (yr. B.P.) |
Profile 1 (upper) 1910±70 |
Profile 4 2630±70 |
Profile 2 (upper) 3870±70 |
Profile 5 | Profile 6 | Profile 3 8180±70 |
Profile 2 (middle) 86660±70 |
Profile 2 (lower) 8740±70 |
Profile 1 (lower) 8990±70 |
|---|---|---|---|---|---|---|---|---|---|
| Trees / fossil type | |||||||||
| Acer negundo / seed | 1 | 1 | 2 | ||||||
| Acer negundo / fruit wing | 1 cf. | 1 cf. | |||||||
| Carya cordiformus / nut frag. | 1 | 1 | |||||||
| Fraxinus americana / fruit | 1 | ||||||||
| Juglans nigra / nut frag. | 1 | ||||||||
| Juniperus virginiana / twigs | 1 | 27 | 1 | ||||||
| Morus rubra / seed | 1 | 1 | 5 | ||||||
| Osirya virginiana / fruit | 1 | ||||||||
| Populus deltoides / capsule segments | 4 | 2 | |||||||
| Populus deltoides / buds | 2 | ||||||||
| cf. Populus deltoides / bud scales | 2 cf. | 3 | |||||||
| Populus cf. Populus / wood (LES) | 5 | 1 | |||||||
| Quercus macrocarpa type / flower bud scales | 4 | ||||||||
| Quercuscf. macrocarpa / twigs | 2 | ||||||||
| cf. Quercus / leaf lobes (white oak group) | 3 | 1 | |||||||
| Quercus / proximal tip (under cap) | 1 | ||||||||
| Salix exiguassp. / leaf equivalents interior type |
1 | 1 | 1 | ||||||
| Salix / capsules | 2 | 6 | 1 | 13 | |||||
| Salixsp. / wood (LES) | 6 | 1 | |||||||
| cf. Salix / buds | 1 | ||||||||
| Tilia americana / fruits | 1 | 1 | |||||||
| Ulmus americana / seeds / fruits | 3 | 4 | |||||||
| Ulmus americana / bud scales | 46 | 22 | 8 | 18 | 8 | 11 | 1 | 15 | 4 |
| Ulmus americana / buds | 4 | 9 | 3 | 1 | 2 | 6 | 4 | ||
| Ulmus americana / wood (LES) | 6 | 2 | 2 | ||||||
| Ulmus rubra / bud scale | 7 | 3 cf. | 1 cf. | 3 cf. | |||||
| Ulmus rubra / wood (LES) | 1 | 1 cf. | |||||||
| other bud scales / bud scales | 30 | 11 | 14 | 60 | 2 | 28 | 31 | 16 | |
| Thick-bottomed, pointed bud scales | 2 | 1 | 1 | ||||||
| Other buds / buds | 8 | 6 | 1 | 3 | 2 | 3 | |||
| Large-leaf fragments | 8 | ~5 | 7 | 3 | 5 | ||||
| Forest floor herbs / Fossil type | |||||||||
| Campanula americana / seeds | 1 | 2 | |||||||
| Laportea canadensis / fruits | 2 | ||||||||
| Salvia lyrata / seeds | 1 | 1 | |||||||
| Shrubs | |||||||||
| Cornus / nutlet fragments | 1 cf. | 1 | |||||||
| Rubus / fruits | 1 | ||||||||
| Sambucus canadensis /seeds | 1 | 1 | 1 | 2 | |||||
| Sambucus / seeds | 1 | ||||||||
| Staphylea trifolia / seeds | 2 | ||||||||
| Vitis / seeds | 2 | 1 | |||||||
| Aquatics | |||||||||
| Heterantheracf. reniformis / seeds | 1 | ||||||||
| Wetlands | |||||||||
| Bidens cernua / fruits | 1 | 9 | |||||||
| Bidens comosa / fruits | 4 | ||||||||
| Bidens frondosa type / fruits | 3 | 2 | 9 | ||||||
| Boehmeria cylindrica / fruits | 1 | 1 | 3 | ||||||
| Carex conoidea / fruits | 1 | ||||||||
| Cyperus aristatus / fruits | 1 | 1 | |||||||
| Cyperus esculentus / fruits | 1 | ||||||||
| Cyperus odoratus / fruits | 1 | 1 | |||||||
| Echinodorus rostratus / seeds | 1 | ||||||||
| Eleocharis palustris / fruits | 2 | ||||||||
| Hemicarpa micrantha / fruits | 3 | ||||||||
| Juncus / seeds | 14 cf. | 2 | |||||||
| Leersia oryzoides / florets | 4 | 3 | |||||||
| Lippia lanceolata / seeds | 1 | ||||||||
| Ludwigia type alterniflora / seeds | 1 | ||||||||
| Lycopus americanus / seeds | 2 | 3 | |||||||
| Penthorum sedoides / seeds | 1 | 1 | 1 | 5 | |||||
| Pilea pumila / fruits | 2 | ||||||||
| Polygonum hydropiperoides / fruits | 1 | ||||||||
| Polygonum lapathifolium / fruits | 3 | 4 | 12 | 1 | 12 | ||||
| Polygonum pensylvanicum / fruits | 2 | 1 | 1 | 1 | |||||
| Polygonum punctatum / fruits | 1 | 2 | |||||||
| Rorippa palustris / seeds | 1 | ||||||||
| Rumex altissimus / fruits with perianth | 1 | 2 cf. | |||||||
| Sagittaria brevirostra (= engelmanniana / fruits |
1 | ||||||||
| Scirpus americanus / fruits | 1 | ||||||||
| Scirpus atrovirens type / fruits | 1 | ||||||||
| Scutellaria / seeds | 1 | 1 | |||||||
| cf. Spartina pectinata / partial floret | 1 | ||||||||
| cf. Suaeda depressa / inner seed coat | 1 | ||||||||
| Prairie plants | |||||||||
| Amorpha canescens / pods | 9 | 9 | 26 | 3 | 16 | 1 | |||
| Amorpha canescens / flower calyces | 5 | 7 | 6 | 14 | 2 | ||||
| Amorpha canescens type / leaflets | 5 | 3 | 50 | 14 | 1 | 50 | |||
| Amorphacf. nana / pods | 1 | ||||||||
| Amorphacf. nana / leaflets | 2 | 2 | 2 | ||||||
| cf. Amorpha / calyx fragments | 4 | ||||||||
| Andropogon gerardii / florets | 1 | 19 | 5 | 2 | 7 | 4 | |||
| Andropogon (Schizachyrium) scoparius / florets | 5 | 2 | 8 | 2 | |||||
| Androsace occidentalis / seeds | 1 | 2 | 22 | 1 | |||||
| Ceanothus americanus type / seed | 1 | ||||||||
| Ceanothus americanus type / fruit | 12 | ||||||||
| Dalea / calyx base | 1 cf. | 1 | |||||||
| cf. Dalea / calyx | 1 | ||||||||
| cf. Dalea / leaflet | 1 | ||||||||
| Daleacf. purpurea / calyces | 3 | ||||||||
| Desmanthus illinoensis / pod fragment | 1 | 1 cf. | |||||||
| Desmanthus illinoensis / leaflet | 1 | 1 cf. | |||||||
| Helianthus grosseserratus type / fruits | 1 | ||||||||
| Helianthus laetiflorus type / fruits | 1 | 1 | 1 | 1 | |||||
| Helianthussp. / fruits | 1 | 4 | 1 | 1 | 1 | ||||
| Linum sulcatum / seeds | 1 | 2 | |||||||
| Monarda fistulosa / seeds | 1 | 1 | |||||||
| Monarda pectinata / seeds | 1 | 1 | |||||||
| Oenothera biennis type / seeds | 1 | ||||||||
| Panicum capillare type / florets | 9 | 1 | 39 | 4 | |||||
| Penstemon digitalis type / seeds | 1 | 2 | 22 | 1 | |||||
| Polygala alba / seeds | 2 | ||||||||
| Polygala verticillata / seeds | 1 | ||||||||
| "popped seeds" / seeds/fruits | 1 | ||||||||
| Ratibida pinnata / fruits | |||||||||
| Schrankia nuttallii / leaflets | 1 | 1 cf. | |||||||
| Sorghastrum nutans / florets | 2 | 2 | 1 | ||||||
| Sporobolus asper / caryopses (maybe not !) - see note |
3 | 10 | 1 | 3 | |||||
| cf. Sporobolus vaginiflorus / caryopses (maybe not) |
13 | ||||||||
| Stipa spartea / awns (fragments) | 1 | 1 | 1 | 1 | |||||
| Vernonia baldwini type / fruits | 2 | ||||||||
| cf. Vernonia / fruits | 1 | ||||||||
| Weeds | |||||||||
| Acalypha virginica type (incl. A. rhomboidea) / seeds |
1 | 1 | 5 | 1 | 1 | ||||
| cf. Acalypha / seeds | 1 | ||||||||
| cf. Amaranthus / seeds | 27 | ||||||||
| Amaranthus / seeds | 16 | 84 | 12 | 6 | 14 | 58 | 47 | 33 | |
| Ambrosia psilostachya type / fruits | 1 | 3 | |||||||
| Ambrosia trifida / fruits | 6 | 2 | 1 | 4 | 1 | 4 | 3 | ||
| Asclepias verticillata type / seeds | 9 | ||||||||
| Chenopodium berlandieri type / fruits | 4 | 2 | |||||||
| Chenopodium bushianum / fruits | 5 | ||||||||
| cf. Chenopodium capitatum / fruits | |||||||||
| Chenopodium gigantospermum type / fruits | 2 | 1 | 1 | 3 | |||||
| Chenopodiumspp. / fruits | 4 | 22 | 4 | 3 | 45 | 89 | 62 | 3 | |
| Descurainia pinnata (combine) / seeds | 1 | ||||||||
| Draba / seeds | 1 | 14 | |||||||
| Echinochloa muricata type / florets | 9 | 1 | 2 | 6 | 9 | 1 | 3 | ||
| Euphorbia glyptosperma / seeds | 3 | ||||||||
| Euphorbia maculata type / seeds | 3 | 1 | |||||||
| Euphorbia maculata type / perianth parts | 4 | 1 | |||||||
| Euphorbia mutans / seeds | 3 | 4 | 4 | 3 | 12 | ||||
| Euphorbiacf. mutans / perianth parts | 1 | 4 | |||||||
| Euphorbia serpens / seeds | 5 | 1 | 1 | 1 | |||||
| Euphorbiacf. serpens / perianth paris | 1 | ||||||||
| Euphorbia spathulata / seeds | 2 | ||||||||
| Euphorbia sp. / locules | 1 | 2 | |||||||
| Lepidium virginicum type / seeds | 4 | 5 | |||||||
| Lepidium / siliques | 4 | 5 | |||||||
| Mollugo verticillata / seeds | 1 | ||||||||
| Oxalis stricta type / seeds | 2 | 1 | 1 | 1 | 1 | ||||
| Phyla lanceolata / seeds | 1 | 2 | |||||||
| Polygonum aviculare type / fruits | 1 | ||||||||
| Polygonum erectum / fruits | 1 type | ||||||||
| Polygonum scandens / fruits | 1 | ||||||||
| Portulaca oleracra / seeds | 1 | 1 | 4 | 1 | 4 | ||||
| Silene antirrhina / seeds | 3 | 1 cf. | 10 | 93 | 62 | 9 | |||
| Triodanis perfoliata type / seeds | 1 | 1 | 1 | 3 | 7 | ||||
| Urtica dioica / seeds | 6 | 2 | 1 | 2 | |||||
| Verbena hastata / seeds | 1 | 1 | |||||||
| Verbena urticifolia / seeds | 1 | 1 | 2 | 1 | |||||
| Verbena cf. urticifolia / seeds | 3 | ||||||||
| Other | |||||||||
| cf. Ammania robusta | 3 | 1 | 4 | ||||||
| Amorpha sp. / pod fragment | 1 | ||||||||
| Carex (biconvex) / fruits | 2 | 1 | 1 | 4 | 1 | 1 | 7 | 16 | |
| Carex (trigonous) / fruits | 2 | 1 | 1 | 2 | 3 | 4 | |||
| Cirsium / fruits | 1 | ||||||||
| Composites / fruits | 2 | 1 | 1 | 3 | 3 | 17 | 1 | ||
| other crucifer / seeds | 1 | 1 | |||||||
| Cyperus / fruits | |||||||||
| Eupatorium purpureum type (incl. E. maculatum and E. fistulosum) |
1 | ||||||||
| Eupatorium sp. / fruits | 1 | 1 | |||||||
| medium grass florets | 3 | ||||||||
| long. narrow grass / caryopses | 12 | ||||||||
| medium grass / caryopses | 1 | 1 | 5 | ||||||
| small mottled cf. grass / caropyses | 1 | 5 | |||||||
| misc. other caryopsis-like forms | 1 | ||||||||
| large grass (prairie) florets (damaged or incomplete) |
2 | 4 | 2 | 48 | 11 | 35 | |||
| other legume calyces | 3 | 3 | |||||||
| large legume seed (charred) | 1 | ||||||||
| small leaflet bases / tips | 6 | 1 | 14 | ||||||
| medium-sized leaflets / leaves | 8 | ||||||||
| monocot leaf tips | 2 | ||||||||
| Panicum / florets | 6 | 5 | |||||||
| Polygonum sp. / fruits | 1 cf. | 2 | 1 | 1 | |||||
| cf. Scrophulariaceae / seeds | 38 | ||||||||
| Solanaceae / seed, | 1 | 1 | |||||||
| legume pod fragments | 1 | ||||||||
| misc. small leaves / leaflets | 1 | 14 | |||||||
| tiny "flowers" | 3 | ||||||||
| umbel / fruit | 3 | ||||||||
| Viola / seeds | 1 | 2 | 1 | 2 | 1 | ||||
| Totals | 229 | 240 | 57 | 340 | 140 | 325 | 285 | 594 | 192 |
| unknowns / indeterminate | 20 | 11 | 11 | 44 | 13 | 35 | 20 | 68 | 38 |
| unidentifiable wood | 3 | 5 | 2 | ||||||
| insects | common | uncommon | present | present | uncommon | ||||
| bryophytes | rare | 1 | none | rare | |||||
| charcoal | rare (woody when present) |
rare | herbaceous charcoal common, woody rare (1 woody chunk saved) |
||||||
| bones | rare | 1 cf fish vertebrae |
|||||||
Table 2-2 is organized by habitat. Each group represents a specific habitat that must have been present at or near the site. The habitat groups are trees, shrubs, forestf-loor herbs, aquatics, wetlands, prairie plants, weeds, and "other" (not sufficiently identifiable to assign to a habitat).
The early Holocene macrofossils, prior to 8500 yrs B.P., had a good representation of mesic deciduous forest trees including oak, hickory, American elm, slippery elm, walnut, ironwood, and red cedar. During this time, prairie elements are also very abundant and diverse, including such indicator species of tallgrass prairies as big bluestern, little bluestem, indian grass, needle and thread, wild bergamot, and lead plant. Phytoliths and 813C values indicate a preponderance of C3 plants, although deciduous-tree pollen is never less than 20 percent. The forest elements in the macrofossil assemblage all but disappear by 8200 yrs B.P. Although sites on the eastern plains are few, data from pollen sites in the Dakotas suggest that deciduous forests were present for between about 11,000-10,000 yrs B.P. on the eastern Great Plains, but died out soon thereafter and were replaced by prairie. These exposures suggest that at least some deciduous trees were able to survive locally until after 8500 yrs B.P. Presumably prairies began to dominate on the drier uplands, and the trees hung on along valley walls and ravines, where cooler, moister conditions were still available.
Other plant communities represented include riparian forests (willow and boxelder), wet meadow/marsh habitats, and disturbed ground/weedy areas. All these habitats are commonly found along floodplains, where erosion and deposition cause disturbance and create oxbows and other wet areas.
By late Holocene (3870 to 1910 yrs B.P.), only riparian forests were present at the Old Bridge site, with cottonwood, willow, boxelder, and elm (a late-successional floodplain as well as an upland forest tree). There was apparently enough shade at 1910 yrs B.P. for some shade-loving forest-floor plants to be present. Macrofossils of prairie species, though less abundant and diverse, are still well represented, and prairie must have been the dominant upland plant cover. Wetland and weedy disturbed habitats were again common on the floodplain.
The Miles alluvial fan is located 8 km downstream from Stop 2 (Stop 3 in fig. 2-6). This fan developed at the mouth of an unnamed intermittent stream that flows into the South Fork of the Big Nemaha River. The channel of the Nemaha is along the north side of the valley wall and is cutting into the midsection of the fan. This has resulted in the development of a steep cutbank exposing a thick section of fan and floodplain deposits (fig 2-16). In addition, a Farmdalian peat is exposed at the bottom of the cutbank.
Figure 2-16--Photograph of the section at the Miles alluvial fan. Note the dark peat at the bottom of the section.

Based on its lithology, the Miles fan is composed of the Corrington Member of the DeForest Formation (fig. 2-17). The fan deposits are oxidized and consist of multiple upward-fining sequences. Two buried soils are developed in the fan: soil 2 at a depth of 130 cm and soil 3 at a depth of 360 cm (figs. 2-16 and 2-17). Decalcified organic carbon from the upper 10 cm of soils 2 and 3 yielded radiocarbon ages of 3130±50 yrs B.P. (Tx-8945) and 10,450±120 yrs B.P. (Tx-8944), respectively.
Figure 2-17--Stratigraphic section for the Miles alluvial fan.
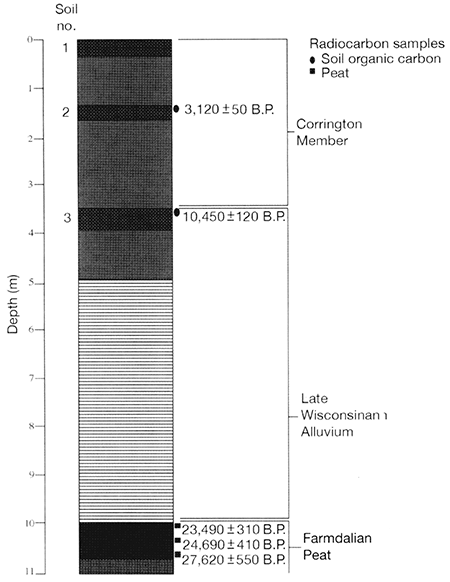
The alluvium below soil 3 is distinctly stratified and consists of fine-grained, late Wisconsinan floodplain deposits (figs. 2-16 and 2-17). A razor-sharp boundary separates these deposits from a 1-m-thick peat that is 9.9-10.7 m below the surface of the alluvial fan. Peat samples collected at depths of 9.9-10.1 m, 10.3-10.4 m, and 10.6-10.7 m yielded radiocarbon ages of 23,490±310 yrs B.P. (Tx-8946), 24,640±410 yrs B.P. (Tx-8947), and 27,580±550 yrs B.P. (Tx-8948), respectively. The peat grades downward into a 75-100-cm-thick unit of gray, organic-rich, pebbly clay loam.
At Stop 3, we will focus on the Fanndalian peat at the base of the fan, and the fossiliferous full-glacial alluvium 100 m downstream from the fan. The pollen record of Fredlund (fig. 2-18) covers only the peat section, and the macrofossil records (fig. 2-19) cover both sections, which are plotted together on diagrams.
We recognize two zones at this locality. The lower zone (ca. 28,000-22,000 yrs B.P.) is characterized by less than 10 percent spruce and very high percentages of non-arboreal pollen. especially grasses and sedges (fig. 2-18). There is also a rich aquatic and wetland macrofossil flora (figs. 2-18 and 2-19).
Figure 2-18--Miles Fan pollen data.

Figure 2-19--Miles Fan aquatic-plant data.
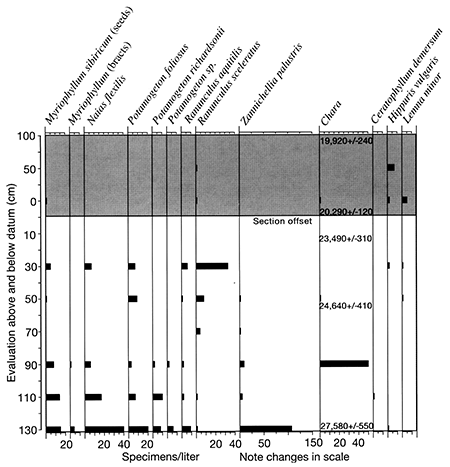
This assemblage indicates that the site was a small pond surrounded by sedges and other wetland plants. Aquatic plants are prevalent in the lower part of the zone but decline as wetland plants increase towards the top. This sequence suggests that the early wetland was deeper, supporting more aquatic plants, but gradually filled in or dried up as wetland elements prevailed. The upland vegetation is represented only by the pollen record and clearly is an open. probably prairie, environment. Such ponds characteristically receive very few macrofossils derived from upland habitats. This interpretation correlates well with the old segment from the Farwell site, with nearby Du Bois quarry, and with the Logan quarry site in southwestern Iowa (Baker et al., 2003, and unpublished data). all of which have prairie plant macrofossils from this interval.
Zone 2 is represented by two upper pollen counts from Miles fan, and by macrofossils from the site 100 m downstream. The pollen from the fan shows an abrupt rise in spruce pollen and a decline of non-arboreal pollen. Spruce macrofossils do not appear in the peat section, but they are very abundant at the downstream section, dating from 20,290 to 19.920 yrs B.P. Most aquatic and wetland macrofossils were not present at this site, but sedges are abundant.
Fredlund interpreted this zone with spruce pollen from the fan as representing local vegetation. However, spruce needles are absent there, whereas they are generally abundant where spruce is locally present. Spruce had clearly arrived in the region, however, because both pollen and macrofossils are abundant at about this time at Logan quarry (Baker, unpublished data) and Muscotah Marsh (Grüger, 1973). It apparently was not growing directly onsite here until about 20,000 yrs B.P. Peat deposition ceased when Wisconsinan alluvium was deposited on top of it. At the downstream section, sedimentation continued for a short time, and spruce was clearly growing on site. It was not possible to determine if it was black or white spruce, but the low-diversity assemblage of abundant spruce and sedge would be consistent with a black spruce wetland. This assemblage is also typical and widespread in the late-glacial sediments of the Midwest.
Muscotah Marsh is located on the floodplain of the Delaware River about 2.5 km south of the village of Muscotah (Stop 4 in fig. 2-6). The marsh is supported by an artesian spring and, as discussed previously, provides the only continuous Holocene pollen record on the eastern edge of the Great Plains (Grüger, 1973). The vegetation of the region prior to European settlement was tallgrass prairie, and the site is located close to a mosaic of deciduous forest and prairie to the east (northeastern Kansas and northwestern Missouri). The Arlington Marsh site, located 5 km south of Muscotah Marsh, also contains pollen-bearing sediments and provides a picture of late Wisconsinan vegetation (Grüger, 1973).
Grüger's (1973) initial analysis of the Muscotah and Arlington marsh sites provides a 25,000-yr record of vegetation change for the region. She identified five zones. The first zone from 25,000 to 23,000 yrs B.P. is composed of relatively open vegetation, with some pine, spruce, and birch, with local stands of alder and willow. It was suggested that the vegetation was similar to what is now found in southern Saskatchewan and southeastern Manitoba. The second zone from 23,000 to 15,000 yrs B.P. is dominated by spruce pollen and represents a spruce forest. There is a hiatus from the second to third zone between 15,000 to 11,000 yrs B.P. The third zone from 11,000 to 9,000 yrs B.P. was composed of a mix of prairie and deciduous oak forest, which was then replaced by prairie around between 9,000 to 5,000 yrs B.P. (fourth zone). The final zone (5,000 yrs B.P. to present) again represented a mix of deciduous oak forest and prairie.
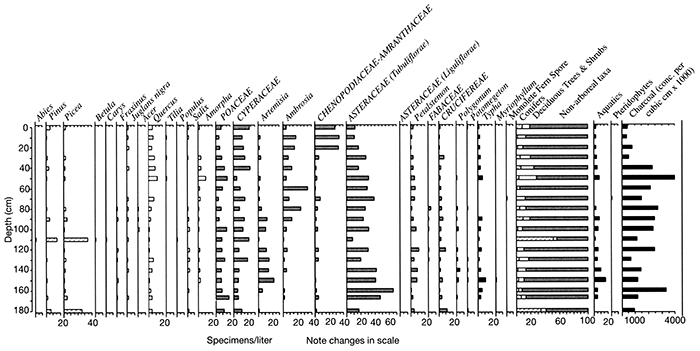
A new core (~10 m in length) was collected from Muscotah Marsh in September 2000 by Baker, Bettis, Mandel, and Moss to examine at higher resolution the Holocene environments of the eastern Great Plains. Moss (unpublished data) has produced a high-resolution (sample interval of 10 cm) pollen record for the first 180 cm of this core (fig. 2-20), which probably covers the late Holocene period. The results of the pollen analysis indicate that the vegetation around Muscotah Marsh was dominated by non-arboreal taxa (relative abundance between 60 to 70%). The dominant taxon from 180 to 30 cm was Asteraceae (Tubuliflorae) and probably reflects the presence of tallgrass prairie. Sagebrush (Artemisia) was also important from 160 to 100 cm. Ambrosia (ragweed) increases from 90 cm and may indicate increased human disturbance (European settlement) and the increased representation of Chenopodiaceae-Amranthaceae from 30 to 0 cm probably reflects intensive agricultural activity in the region. Grasses and sedges are important parts of the non-arboreal vegetation throughout the length of the record. Deciduous forest taxa (particularly oak) maintain consistently low values (5 to 10%) throughout the length of the record and reflect the regional presence of deciduous forest. Conifers also maintain low representation (less than 5%), except at 180 cm (~30%) and 110 cm (~40%), where spruce and pine peak; this may reflect contamination of the record or stronger westerly winds that were transporting these pollen types over long distances from the Rocky Mountains. The presence or absence of spruce needles at these depths in the record will hopefully resolve the cause of these conifer peaks. Carbonized particle analysis of the Muscotah Marsh record suggests relatively high fire regimes from 180 to 50 cm and with lower values from 40 to 0 cm, probably reflecting the imposition of fire suppression by EuroAmericans.
Figure 2-20--Pollen record for Muscotah Marsh.
Baker, R. G., 2000, Holocene environments reconstructed from plant macrofossils in stream deposits from southeastern Nebraska: The Holocene, v. 10, p. 357-365.
Baker, R. G., Bettis, E. A., III, and Huber, J. K., 2003, Vegetation on the eastern Great Plains, North America, during mid-Wisconsinan time---Late MIS Stage 3-early MIS Stage 2: Programs with Abstracts, XVI INQUA Congress, Reno, Nevada, p. 164
Baker, R. G., Maher, L. J., Chumbley, C. A., and Van Zant, K. L., 1992, Patterns of Holocene environmental change in the Midwest: Quaternary Research, v. 37, p. 379-389.
Baker, R. G., Bettis, E. A., III, Schwert, D. P., Horton, D. G., Chumbley, C. A, Gonzalez, L. A, and Reagan, M. K., 1996, Holocene paleoenvironments of northeast Iowa: Ecological Monographs, v. 66, p. 203-234.
Baker, R. G., Fredlund, G. G., Mandel, R. D., and Bettis, E. A., III, 2000, Holocene environments of the central Great Plains: Quaternary International, v. 67, p. 89-106.
Baker, R. G., Bettis, E. A., III, Gonzalez, L. A, Denniston, R. E, Strickland, L. E., and Krieg, J. R., 2002, Holocene paleoenvironments in southeastern Minnesota-Chasing the prairie-forest ecotone: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 177, p. 103-122.
Barnosky, C. W., Grimm, E. C., and Wright, H. E., Jr., 1987, Towards a postglacial history of the northern Great Plains: A review of the paleoecologic problems: Annals of Carnegie Museum, v. 56, p. 259-273.
Bettis, E. A., III, 1995, The Holocene stratigraphic record of entrenched stream systems in thick loess regions of the Mississippi River basin: Ph.D. thesis, University of Iowa, 149 p.
Chumbley, C. A, Baker, R. G., and Bettis, E. A., III, 1990, Midwestern Holocene paleoenvironments revealed by floodplain deposits in northeastern Iowa: Science, v. 249, p. 272-274.
Dillon, J. S., 1992, Holocene alluvial stratigraphy and landscape development in Honey Creek basin, Nemaha County, Nebraska: M.A. thesis, University of Nebraska at Omaha, 212p.
Fredlund, G. G., and Tieszen, L. L., 1997, Phytolith and carbon isotope evidence for late Quaternary vegetation and climate change in the southern Black Hills, South Dakota: Quaternary Research, v. 47, p. 206-217.
Grimm, E. C., 2001, Trends and palaeoecological problems in the vegetation and climate history of the northern Great Plains, U.S.A.--Biology and environment: Proceedings of the Royal Irish Academy, v. 101B, p. 47-64.
Grüger, J., 1973, Studies on the late Quaternary vegetation history of northeastern Kansas: Geological Society of America, Bulletin 84, p. 239-250.
Johnson, W. C., and Fredlund, G. G.,1985, A procedure for extracting palynomorphs (pollen and spores) from clastic sediments: Transactions of the Kansas Academy of Science, v. 88, p. 51-58.
Laird, K. R., Fritz, S. C., Cumming, B. E, and Grimm, E. C., 1998, Early Holocene limnological and climatic variability in the northern Great Plains: The Holocene. v. 8, p. 275-285.
Mandel, R. D., 1994. Holocene landscape evolution in the Big Blue and Little Blue River valleys, eastern Nebraska--Implications for archeological research; in, Blue River Drainage Intensive Archeological Survey. 1992-1993, Seward and Thayer Counties, Nebraska, Volume I, E. J. Lueck and R. P. Winham, eds.: Sioux Falls, South Dakota. Augustana College. Archeology Laboratory. Archeology Contract Series no. 84, p. H-I-H-79.
Mandel, R. D., 1996. Geomorphology of the South Fork Big Nemaha River valley, southeastern Nebraska; in, A Geoarchaeological Survey of the South Fork Big Nemaha Drainage. Pawnee and Richardson Counties, Nebraska, S. R. Holen, J. K. Peterson, and D. R. Watson. eds.: Nebraska Archeological Survey. Technical Report 95-02. University of Nebraska State Museum, p. 26-81.
Mandel, R. D., and Bettis, E. A., III, 1995, Late Quaternary landscape evolution and stratigraphy in eastern Nebraska; in, Geologic Field Trips in Nebraska and Adjacent Parts of Kansas and South Dakota, C. A. Flowerday, ed.: 29th Annual Meeting of the North-central and South-central Sections, Geological Society of America: Conservation and Survey Division, University of Nebraska, Guidebook No. 10, p. 77-90.
Mandel, R. D., and Bettis, E. A., III, 2001. Late Quaternary landscape evolution in the South Fork of the Big Nemaha River valley, southeastern Nebraska and northeastern Kansas: Midwest Friends of the Pleistocene 47th Field Conference, Guidebook No. II, Conservation and Survey Division, Institute of Agriculture and Natural Resources, University of Nebraska, Lincoln, 58 p.
Mandel, R. D., and Bettis, E. A., III, 2003, Characteristics of the Severance Formation--A new late Quaternary lithostratigraphic unit in the Eastern Plains of North America: Abstracts with Program, 37th Annual Meeting of the North-Central Section of the Geological Society of America. Kansas City. Missouri, p. 60.
Tomenek, G. W., and Hulett, G. K., 1970, Effects of historical droughts on grassland vegetation in the Central Great Plains; in, Pleistocene and Recent Environments of the Central Great Plains. W. Dort, Jr., and J. K. Jones. cds.: University Press of Kansas, Lawrence, p. 203-210.
Weaver, J. E., 1968. Prairie plants and their environment: University of Nebraska Press, Lincoln, 276 p.
Webb. T., III, Bartlein, P. J., Harrison. S. P., and Anderson. K. H., 1993, Vegetation, lake levels, and climate in eastern North America for the past 18,000 years; in, Global Climates Since the Last Glacial Maximum, H. E. Wright, Jr., 1. E. Kutzbach, T. Webb, 1II. W. F. Ruddiman, F. A. Street-Perrott, and P. J. Bartlein, eds.: Minneapolis, University of Minnesota Press. p.415-467.
Wright, H. E., Jr., Almendinger, J. c., and Grüger, J., 1985, Pollen diagram from the Nebraska Sandhills and the age of the dunes: Quaternary Research, v. 24, p. 115-121.
Prev--Pre-Meeting Field Trip 1 || Next--Post-meeting Field Trip 3
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version July 7, 2014. Original publication date 2006.
URL=http://www.kgs.ku.edu/Publications/Bulletins/TS21/field_trip2.html