![]()
![]()
![]()
Prev--Wisconsinan Mammalian Faunas || Next--Late Quaternary Palynology and Paleobotany
Pleistocene faunal provinces in North America correspond to modern biomes. These Pleistocene biomes were antecedent to but different from modern biomes. Community structure within the Ice Age biomes was more complex and niche width would have been less. The simplification of these biomes at the end of the Pleistocene may have been a major factor in the extinction at the end of the Pleistocene.
The study of faunal provincialism is dependent on the generation of maps depicting the distribution of species comprising the fauna. Such maps are uncommon in vertebrate paleontology and very little serious work has been done on the interpretation of fossil-vertebrate distributions. Vegetational data are much more likely to be summarized, but usually for a very restricted region, and until quite recently, enormous regions lacked data. This severely handicapped studies of even Pleistocene vegetational distributions. In the west, the addition of vegetational data collected from woodrat middens of Pleistocene age (Wells, 1976; Wells and Hunziker, 1979; VanDevender and Mead, 1976) has filled in much that was missing, and recent studies in the central Great Plains have made it possible to provide a vegetational reconstruction for North America during the Wisconsinan glacial maximum. Wells (unpublished) prepared such a reconstruction and made it available to Martin and Neuner (1978) who were then able to compare the distribution of Wisconsinan mammals with the vegetational reconstruction.
We have benefited from conversations with W. Johnson, W. Dort, Jr., and J. D. Stewart. M. A. Klotz drafted the figures, and M. Slade typed the manuscript.
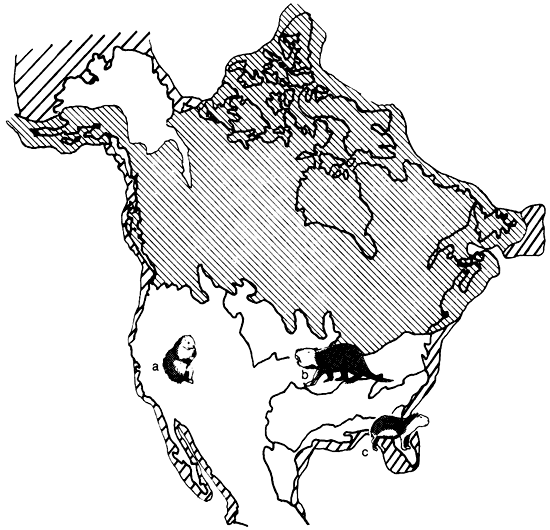
Martin and Neuner (1978) recognized distinct assemblages of Pleistocene mammals that tended to co-occur with each other and found that their regions of co-occurrence corresponded very closely to mapped areas of roughly uniform vegetational types (fig. 1). These regions of co-occurring plants and animals correspond closely to the classical "biome" concept of ecologists (fig. 2). However, following standard practice in invertebrate paleontology, Martin and Neuner (1978) called these regions faunal provinces and gave them names based on characteristic animals that occurred in them. This was done in part to emphasize that they are not defined by their geographic location, but by biota, and might change size, shape, and location during geological time. Only regions south of the continental ice were included and Beringia, which was intimately connected with Siberia at that time, was not considered. The Wisconsinan flora of Beringia seems to have been in part a unique sort of grassy tundra called Steppe Tundra by Guthrie (1968). A number of animals inhabited this region that are not known to have occurred south of the continental ice sheet, including the yak Bos (Poephagus) grunniens and the so-called steppe antelope Saiga tatarica. We have since termed this the "Saiga faunal province" (Martin et al., 1985).
Figure 1--North American Wisconsinan Faunal Provinces (based on Martin and Neuner, 1978). Narrow hatching indicates continental ice and wide hatching indicates areas of exposed continental shelf.
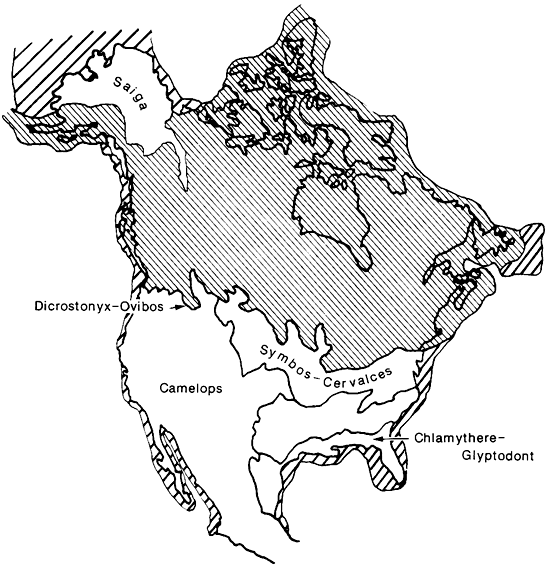
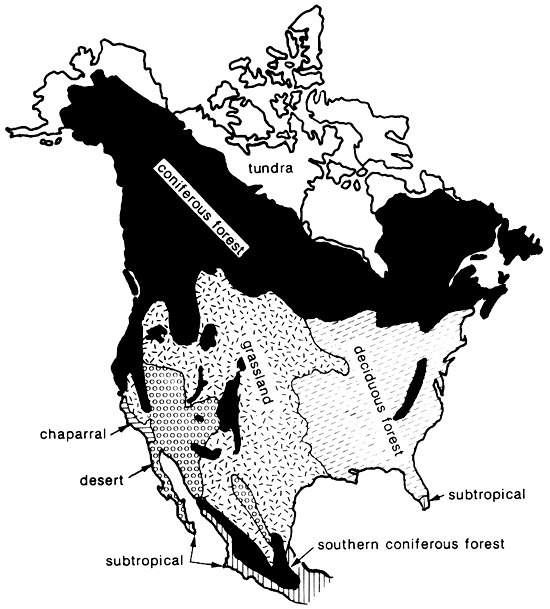
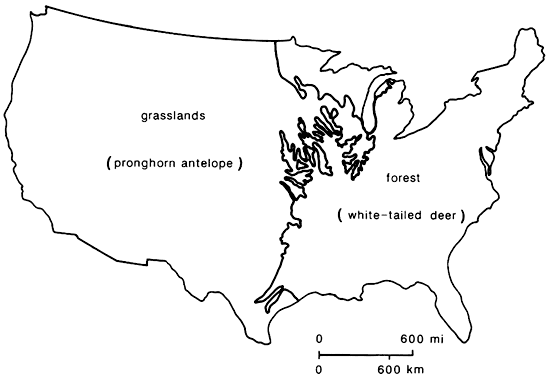
Figure 2--Schematic map of major biomes in North America, based on Shelford (1963). The woodland biome of the Southwest is not represented for reasons of clarity and scale, but it occurs at intermediate elevations between grassland/ desert and coniferous forest.
In parts of Beringia, as well as along the southern glacial margins and in regions of high elevations, true tundra like the modern high arctic or alpine occurred, along with the tundra muskox (Ovibos), caribou (Rangifer), and collared lemming (Dicrostonyx). This assemblage characterizes the Ovibos faunal province and still exists although in a very different location from its Pleistocene distribution.
South of the continental ice sheet, the tundra was bordered on the south by a boreal forest largely dominated by spruce (Picea). This forest contained a highly characteristic fauna including the American mastodon (Mammut), woodland muskox (Bootherium) (= Symbos), extinct stagmoose (Cervalces), and the giant beaver (Castoroides). The above association defines the Symbos-Cervalces faunal province. South of the southeastern half of this faunal province was a large region of mixed boreal and deciduous forest that in fact seems to have included the primary refugia for the taxa that now dominate the eastern deciduous woodlands. It contains an interesting fauna largely dominated by more widely ranging forms. Even though no taxa are endemic, certain ones had their principal distributions here; we term this the Odocoileus-Pitymys faunal province.
Farther south the region that included Florida and the Gulf Coast seems to have had a remarkably tropical aspect even during periods of glacial advance. Wells mapped this region as mostly southern deciduous woodlands (although evergreen broadleafed trees and shrubs also were present), and the associated animals are forms that now have their closest relatives in Central and South America, including glyptodonts (Glyptotherium), chlamytheres (Holmesina), capybaras (Hydrochoerus), and the spectacled bear (Tremarctos).
The lowlands of the western United States, including most of the region that is now desert and steppe, were occupied by a variety of conifer parklands. The trees that dominated these parklands varied but the structure of the vegetation (scattered stands of trees with large areas of intervening grass) may have been consistent over most of this region. This is the area where we find most of the large grazers and the large pursuit predators that hunted them, including the American camel (Camelops), the extinct antilocaprid (Capromeryx), most of the records of the imperial mammoth (Mammuthus), the short-faced bear (Arctodus simus), and the American lion (Panthera atrox). This fauna had grassland aspects but many of the constituent species seem to also have browsed and may have been tied to the C3 grasses that were abundant in this region during the Pleistocene.
One thing which seems evident when we compare the reported floras from these regions with the faunas occupying them is that the individual species associations did not control the animal distributions as much as the general structure of the vegetational community. The presence or absence of large open spaces is especially important. The types of trees (conifer or deciduous) also must strongly influence the associated fauna, and even the carbon pathway used by the grasses (C3 or C4) affects the quality of summer and winter forage and the time of the year when it would have been desirable to have young.
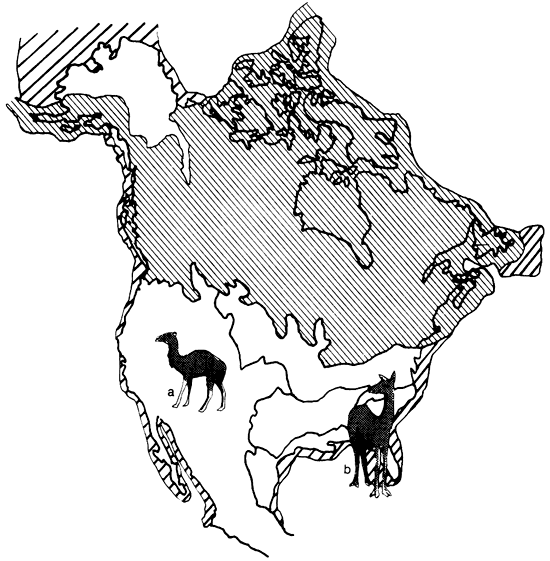
The occurrence of such distinct vegetational regions during the Pleistocene resulted in diversification so that each area may be characterized by its particular species. The most obvious example may be found among the Ovibovinae (fig. 3). The Ovibos-Dicrostonyx faunal province is dominated by the tundra muskox Ovibos, the Symbos-Cervalces faunal province by the forest muskox Bootherium, and the Camelops faunal province by the shrub-ox Euceratherium. Among the large cats of the genus Panthera, we find the American lion (P. atrox) mostly in the Camelops faunal province while the American jaguar P. onca augusta occurred mainly in the Chlamythere-Glyptodont faunal province. The American camels (Camelops, Hemiauchenia) are restricted to the Camelops faunal province while the llama (Palaeolama) is found in the Chlamythere-Glyptodont faunal province (fig. 4).
Figure 3--Bovids characteristic of different faunal provinces: a) shrub-ox Euceratherium; b) tundra muskox Ovibos; c) woodland muskox Bootherium, (= Symbos); d) yak Bos (Poephagus),
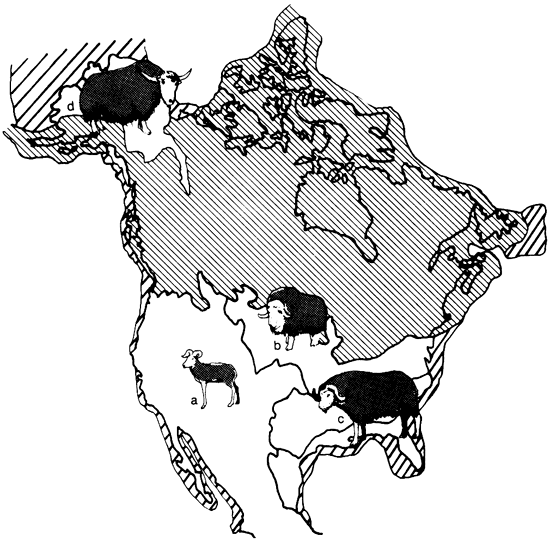
Figure 4--Camelids characteristic of different faunal provinces: a) American camel Camelops; b) stout-legged llama Palaeolama.
The prairie dogs (Cynomys) were characteristic of the Camelops faunal province while the giant beaver (Castoroides) and the capybara (Hydrochoerus) were characteristic of the Symbos-Cervalces and Glyptodont-Chlamythere faunal provinces respectively (fig. 5).
Figure 5--Rodents characteristic of different faunal provinces: a) prairie dog Cynomys; b) giant beaver Castoroides; capybara Hydrochoerus.
One of the characteristics of the modern depauperate large-mammal fauna is that many of the surviving forms actually seem to have been more wide-ranging in the Pleistocene than their extinct relatives and continued to occupy very extensive ranges in the Holocene. Examples include the bison (Bison bison), wapiti (Cervsu elaphus), and white-tailed deer (Odocoileus virginianus). Some, however, were (and are) more restricted; the pronghorn (Antilocapra) and mountain sheep (Ovis canadensis) to the Camelops faunal province and the moose (Alces alces) to the Symbos-Cervalces faunal province and its taiga-forest successor.
While the influence of regional climate may determine the overall character of any particular faunal province, the actual boundaries between faunal provinces must be controlled by local factors of microclimate, soils, and topography. These factors usually are integral parts of the local physical geography such as elevation change, major substrate change, or drainages. The geographical positions of these local factors may be geologically constant so that we might expect them also to influence the position of boundaries between past, as well as present, biomes. This seems to be the case, and many of the vegetational areas proposed by Wells share similar boundaries with major changes in modern vegetations. The very close correspondence between fossil-mammal distributions and the boundaries of the Pleistocene vegetations supports the validity of his procedure.
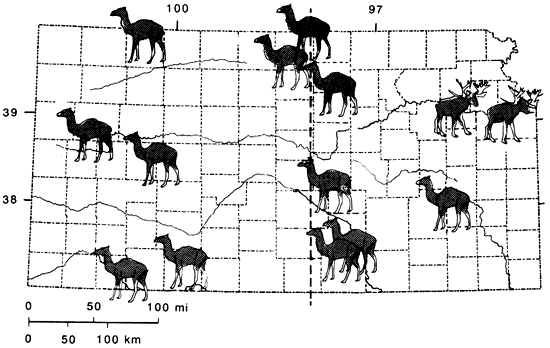
A very striking case occurs with the Pleistocene fauna in Kansas where the boundary between the Camelops faunal province and the Symbos-Cervalces faunal province falls along the Tatschl line proposed by Kuchler (1970; fig. 6). This line seems to mark a major division between many species and subspecies of plants and animals, and some of the speciation related to this line might possibly have involved Pleistocene biogeography.
Figure 6--Distribution of Wisconsinan camels (Camelops) and stag-moose (Cervalces) in Kansas. The vertical line indicates the approximate modern position of the Tatschl line.
As noted above, the term "faunal province" is not usually employed by biogeographers in dealing with modern biota. However, the concept of the "biome" is the modern analog. Biomes of North America have been described in great detail by Shelford (1963), and while there is some variation in usage by different authorities (cf. Kendeigh, 1961; Udvardy, 1975), there is general comparability (fig. 2). The Pleistocene faunal provinces, delineated by Martin and Neuner (1978) and employed in this paper, are far from identical with any modern biomes, yet they serve as antecedent and, in part, ancestral communities to modern biomes (fig. 1). The Saiga faunal province, with its steppe-tundra vegetation and fauna, has no modern analog, but the Ovibos faunal province has been succeeded by the tundra biome. Similarly, the Symbos-Cervalces faunal province has been succeeded by the boreal-coniferous forest, or taiga, biome. The Chlamythere-Glyptodont faunal province is represented today in North America only by relict fragments of subtropical/tropical-forest biome in southern peninsular Florida and along the coastal Mexican lowlands (fig. 2). These last three biomes now occupy areas that are far removed from the areas covered by their antecedent Pleistocene faunal provinces, the first two being more northerly now, while the last is more southerly. This is strong evidence for a marked decrease in mid-latitude climatic equability in Holocene time, compared with late Pleistocene conditions.
In contrast to this situation, the ecotone between Symbos-Cervalces and Chlamythere-Glyptodont faunal provinces, which we herein name as the Odocoileus-Pitymys faunal province, contained the principal species that now occupy the temperate deciduous-forest biome. This Holocene biome is centered on the Pleistocene ecotone and has essentially developed in situ by expansion of deciduous and evergreen broadleafed trees and emigration north and south respectively of those boreal and subtropical elements that formerly comprised the ecotone. Similarly, the Camelops faunal province gave rise, by segregation and differentiation of vegetational elements, not only to the Holocene woodland biome, but also to the temperate grassland and desert biomes of western North America. These too evolved basically in situ, and the modern biomes occupy the same area covered by the lowland portion of the Camelops faunal province.
The ecotone between Camelops, Symbos-Cervalces, and Odocoileus-Pitymys faunal provinces was fairly narrow and well-defined (fig. 1) The same is true of the ecotone between the grassland and deciduous-forest biomes today (fig. 2). Moreover, both ecotones are in the same position, approximately along the Tatschl line (fig. 6). This also may be seen in the geographic ranges of certain modern ungulate species that survived the Pleistocene/Holocene megafauna! extinction (fig. 7). The pronghorn (Antilocapra americana), which was a rare member of the Camelops faunal province, became a common species of the grassland and desert biomes. The mule deer (Odocoileus hemionus) manifests a similar Pleistocene and Holocene distribution. In contrast, the closely related white-tailed deer (O. virginianus) has a Pleistocene record "...concentrated in the central and eastern parts of the continent" (Kurten and Anderson, 1980, p. 310), though it also has been found in late Pleistocene sites in Nevada and New Mexico. Modern populations also are concentrated in the temperate-deciduous biome, but not restricted therein.
Figure 7--Generalized distribution of pronghorn, a grassland species, and white-tailed deer, a deciduous-forest species, related to the distribution of riparian forests across the Midcontinent. Eastward dispersal of pronghorns across the upper Missouri River was possible because the riparian forest barrier thinned to the north.
While these two species of Odocoileus represent sister species, their divergence is at least early Pleistocene. In contrast, certain sister taxa of small mammals possess genetic and behavioral characters that suggest much more recent divergence, perhaps as recent as the beginning of the Holocene. A recently studied example involves those populations of Plains pocket gophers usually referred to Geomys bursarius. Careful analysis (Heaney and Timm, 1983; Burns et al., 1985) has shown that in Kansas and Nebraska there are actually two distinct morphs, the western lutescens and the eastern bursarius (fig. 8). These two taxa show no evidence of intergradation in morphological characters except in one restricted locality in north-central Nebraska, where a narrow (2-km [1.2-mi]) contact zone occurs (fig. 9). However, here there is morphological, chromosomal, and biochemical evidence of gene flow between the two taxa. These results are open to different interpretations; Burns et al. (1985, p. 102) regard the gene flow as evidence that "...the two taxa should be regarded as subspecies of G. bursarius rather than distinct species," while Heaney and Timm (1983) hold the opposite viewpoint. Of broader biogeographic and evolutionary significance is the fact that the present boundary between these two taxa also is roughly along the Tatschl line. This suggests that this long-standing ecotonal region may be continuing to play a role in the evolutionary divergence of populations.
Figure 8--Distribution of Geomys in the central and northern Great Plains. Solid circles = G. bursarius bursarius, open circles = G. b. illinoensis, open squares = G. b. wisconsinensis, solid squares = G. lutescens lutescens, solid triangles = G. l. major, and open triangles = G. breviceps sagittalis (Heaney and Timm, 1983).
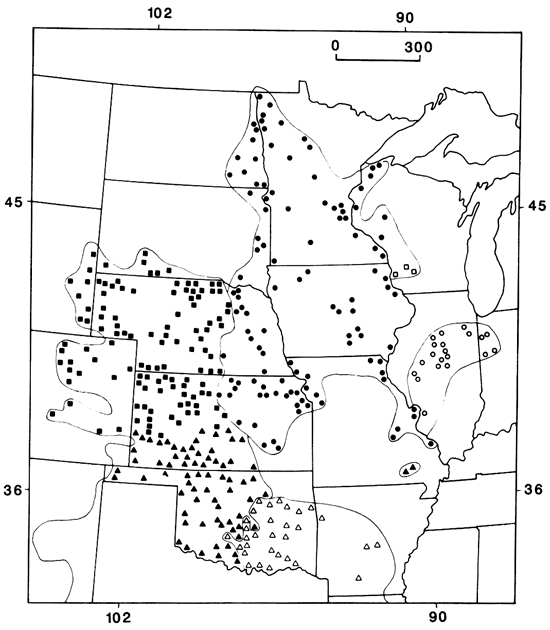
Figure 9--Map of central and northern Great Plains showing levels of similarity of populations of adult female Geomys based on cluster analysis (Heaney and Timm, 1983). Numbers indicate location of populations sampled.
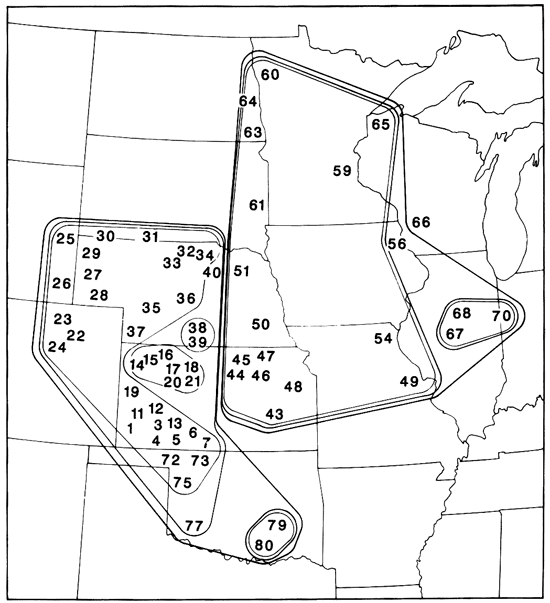
Several more examples may be cited. Big brown bats (Eptesicus fuscus) have a dark eastern subspecies E. f. fuscus in the temperate deciduous biome and a pale subspecies E. f. pallidus in the grassland biome. These two taxa are distinct physiologically as well as morphologically, because fuscus gives birth to two young, while pallidus produces only one. The nature of variation in litter size across the ecotone has not been reported. Other subspecies boundaries among grassland small mammals also show this geographic coincidence, for example in the thirteen-lined ground squirrel (Spermophilus tridecemlineatus) and the northern grasshopper mouse (Onychomys leucogaster). This is not to say, of course, that the boundaries of all species and subspecies fall within this ecotone, as a glance at any general source (e.g., Hall, 1981) will show. Nevertheless, a general concordance exists among a large number of taxon boundaries in this ecotone. If one regards well-defined subspecies as incipient species, then the Tatschl line may mark a zone of significant evolutionary divergence that has been of evolutionary importance for several million years.
The late Pleistocene faunal provinces of Martin and Neuner (1978) really correspond to Ice Age "biomes," which while analogous are not directly comparable to the Holocene biomes that replace them. The Pleistocene biomes contained a greater diversity of both plants and animals and may be thought of as being more complex, with more opportunities for coevolved systems, and probably with selection for narrower species niches. These complex biomes were maintained by climates of low seasonality (Graham and Lundelius, 1984). With the inception of higher seasonality in the Holocene, overall biotic diversity went down and the community structures simplified. This resulted in the disruption of a wide variety of adaptive relationships and finally to widespread extinction and range retraction. The new Holocene biomes were composed largely of surviving subsets of plants and animals that had previously occupied the areas of the antecedent Pleistocene biomes and to a smaller extent immigrants from other regions. The animals and plants that now dominate the Holocene biomes are often forms that had very restricted niches and seem to have been present in low densities during the Pleistocene. In many instances they may have been narrowly specialized to habitats that were limited in extent and importance during the Ice Age but became important constituents of Holocene biomes.
If we are correct in our speculation that mammal species in Pleistocene biomes usually possessed narrower ecological niches than those of Holocene biomes, then this would have permitted greater sympatry and large ranges, thus contributing to greater species richness at any one place. The causal chain may in fact be reversed, in that high species richness in an environment with low seasonality might lead to a higher level of interspecific competition, which would lead to compression of realized niche breadth.
Alternatively, intensified competition could result in allo- or parapatric distribution of species possessing limiting similarities of their fundamental niches. This would have led to geographic vicariance of similarly adapted species seen in some of the Pleistocene fauna, such as the Ovibovinae and others (see above, figs. 3-5).
With the onset of rapid climatic change at the end of the Pleistocene, species with narrow niches, which also exhibited strong competitive interactions with other sympatric species, would have been particularly vulnerable to the breakdown of existing community structure. Those species that would have had the greatest probability of surviving would be of two sorts. One class of Holocene survivors would be those generalists with broad niches which, although perhaps scarce in any particular community, nevertheless were widespread. As communities were restructured across the Pleistocene-Holocene transition, these species had sufficient adaptability, because of their niche breadth, to succeed and eventually to prosper in the new communities, often without major changes in their distribution. Deer (Odocoileus sp.), wapiti (Cervus elaphus), and bison (Bison bison) may be examples of this sort of survivor.
A second class of survivor species may have been those which, though characterized by narrow niches and perhaps scarcity, were "fortunate" in that they were adapted to one of the reduced suite of niches that continued to exist in Holocene communities. Examples may be the tundra musk-ox (Ovibos moschatus), caribou (Rangifer tarandus), and moose (Alces alces) among large ungulates, and the taiga vole (Microtus xanthognathus) or round-tailed muskrat (Neofiber alleni) among small rodents. Such species must have "tracked" their niches through space and time, for they are often found far from their Pleistocene range today. They also may have become more abundant in the Holocene, as some of their competitors became extinct locally or globally, but some still persist only as relict species with limited distribution and numbers.
The depauperate Holocene fauna show particular reduction of the megafauna! component. This is in large part a phenomenon of scale. The larger the species, the larger its home-range requirements and the less likely that it would be able either to adapt to (class 1) or track (class 2) habitats that met its niche requirements during the Pleistocene-Holocene transition. This would have been particularly true of those large species with narrow niches and many congeners, such as North American Pleistocene horses (Equus). Thus not surprisingly the probability of an Equus surviving the restructuring of communities during the post-Pleistocene climatic shift would have been low. Species of smaller body size, with smaller home-range needs to satisfy niche requisites, would conversely have a greater probability of adapting to, or tracking, suitable habitats.
The result is a Holocene fauna with fewer large mammals, lacking certain groups (horses, camels, proboscidians, etc.) entirely, and with most surviving species being the only representative of their genus or higher taxonomic groups, but having large distributions and usually broad niches. Medium- to small-mammal species have mostly survived the transition but often with reduced distributions; niche breadth may have remained about the same. If congeners occur, distinct patterns of geographical vicariance are often seen. These may be inferred to be 1) competitive in ultimate cause, as for example in the two species of lynx (Lynx canadensis, L. rufus) or wolves (Canis lupus, C. niger); 2) a combination of competition and specific-niche adaptation, as among the flying squirrels (Glaucomys sabrinus, G. volans); or finally, 3) recency of species divergence, as in the Great Plains pocket gophers (Geomys) discussed above (see figs. 8 and 9) or in prairie dogs (Cynomys) or some ground squirrels (Spermophilus) and chipmunks (Tamias), where final species separation may date only to the Wisconsinan or early Holocene (Hoffmann, 1981).
Actual boundaries of both Pleistocene and Holocene biomes seem to be largely fixed by physical parameters such as rain-shadows, elevation changes, and river drainages. The position of the environmental effects produced by parameters may be relatively constant geologically, so that in many cases the boundaries of Pleistocene and Holocene biomes closely approximate each other. In the case of the Tatschl line across the Midcontinent, which presently delineates the approximate boundary between the western grassland biome and eastern deciduous-forest biome, the critical environmental effects seem to be the amount and frequency of precipitation and the probability of fire occurrence. Warm-season precipitation derives primarily from storms moving up the Mississippi Valley from the Gulf of Mexico, whereas winter precipitation is usually from storms that are Pacific in origin and often move up across the southern cordillera. The Tatschl line may be thought of as a "tension zone," east of which precipitation is sufficient to support a variety of temperate deciduous trees and to reduce the probability of natural fire. To the west, lower precipitation is less favorable to tree growth, while increasing the probability of fire, which further inhibits woody vegetation while encouraging grasses.
During the Wisconsinan, the same physical parameters (i.e., Gulf and Pacific storm tracks) were present, and while their relative magnitudes may have been modified by differences in general atmospheric circulation under Ice Age conditions, they nonetheless formed a tension zone in approximately the same position as the Tatschl line. The greater extent of woodland and savanna then, in what is now mostly grassland and desert, may be attributable to the greater relative importance of the Pacific storm track bringing more precipitation, especially in summer, to the Southwest and the Midcontinent. Fire must still have been of some importance in maintaining these communities, however, as it is today in Holocene savannas and woodlands.
If this proposed model of the establishment of Pleistocene and Holocene biome boundaries and of the nature of the post-Pleistocene faunal transition is correct, it should be applicable to other biomes and faunas. We hope to examine some of its predictions in future studies.
Burns, J. C., Choate, J. R., and Zimmerman, E. G., 1985, Systematic relationships of pocket gophers (genus Geomys) on the central Great Plains: Journal of Mammalogy, v. 66, p. 102-118.
Graham, R. W., and Lundelius, E., 1984, Coevolutionary disequilibrium and Pleistocene extinctions; in, Quaternary Extinctions-A Prehistoric Revolution, P. S. Manin and R. G. Klein, eds.: Universi1y of Arizona Press, Tucson, p. 223-249.
Guthrie, R. D., 1968, Paleoecology of the large-mammal community in interior Alaska during the late Pleistocene: American Midland Naturalist, v. 79, p. 346-363.
Hall, E. R., 1981, The mammals of North America: John Wiley, New York, second edition, v. 1, p. 1-600 + 90 p.; v. 2, p. 601-1,181 + 90 p.
Heaney, L. R., and Timm, R. M., 1983, Relationships of pocket gophers of the genus Geomys from the central and northern Great Plains: University of Kansas, Museum of Natural History, Miscellaneous Publication 74, p. 1-59.
Hoffman, R. S., 1981, Different voles for different holes-environmental restrictions on refugial survival of mammals; in, Evolution Today, G. G. E. Scudder and J. L. Reveal, eds.: Proceedings, 2nd International Congress on Evolutionary Biology, Vancouver, B.C.. p. 25-45.
Kendeigh, S. C., 1961, Animal ecology: Prentice-Hall, Englewood Cliffs, New Jersey, 468 p.
Kuchler, A. W., 1970, A biogeographical boundary--the Tatschl line: Transactions of the Kansas Academy of Science, v. 73, p. 298-301.
Kutten, B., and Anderson, E., 1980, Pleistocene mammals of North America: Columbia Universi1y Press, New York, 440 p.
Manin, L. D., and Neuner, A. M., 1978, The end of the Pleistocene in North America: Transactions of the Nebraska Academy of Science, v. 6, p. 117-126.
Manin, L. D.. Rogers, R. A., and Neuner, A. M., 1985, The effect of the end of the Pleistocene on man in North America; in, Environments and Extinctions--Man in Late Glacial North America, J. I. Mead and D. J. Meltzer, eds.: Center for the Study of Early Man, Orono, Maine, p. 15-30.
Shelford, V. E., 1963, The ecology of North America: Universi1y of Illinois Press, Urbana, 610 p.
Udvardy. M.D.F., 1975, World biogeographic provinces-map; in, A Classification of the Biogeographic Provinces of the World: International Union of Conservation of Nature and Natural Resources (IUCN), Occasional Paper 18.
VanDevender, T. R., and Mead, J. I., 1976, Late Pleistocene and modern plant communities of Shinumo Creek and Peach Springs Wash, lower Grand Canyon, Arizona: Journal of the Arizona Academy of Sciences, v. 11, no. 1, p. 16-22.
Wells, P. V., 1976, Macrofossil analysis of wood rat (Neotoma) middens as a key to the Quaternary vegetational history of arid America: Quaternary Research, v. 6, p. 223-248.
Wells, P. V., and Hunziker, J. H., 1979, Origin of the creosote-bush (Larrea) deserts of southwestern North America: Annals of the Missouri Botanical Garden, v. 63, p. 43-861.
Prev--Wisconsinan Mammalian Faunas || Next--Late Quaternary Palynology and Paleobotany
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version updated March 31, 2010. Original publication date 1987.
URL=http://www.kgs.ku.edu/Publications/Bulletins/GB5/Martin2/index.html