![]()
![]()
![]()
Prev--Wisconsinan Mammalian Faunas || Next--Grass-opal phytoliths as climatic indicators
The central Great Plains is the heart of the North American grasslands province. Using published and unpublished pollen and other botanical evidence, an outline of the late Quaternary vegetational history for the region is presented. Pre-Wisconsinan pollen records indicate that grasslands were characteristic of the central Great Plains during several interglacial or interstadial climatic episodes but were greatly reduced or even absent during glacial maxima. During the Farmdalian mid-Wisconsinan interstadial, grassland vegetation extended northward at least to the Sandhills of Nebraska, and eastward to Missouri and Iowa where it merged with open coniferous forest. At the onset of the Woodfordian (ca. 22,000 yrs B.P.), the regional vegetation rapidly changed toward greater forest cover. Spruce forest invaded as far south as northeastern Kansas and the Missouri Ozarks. To the west and south the forest was broken into a mosaic of spruce, mixed spruce-deciduous, and aspen with extensive prairie openings. Much of eastern Kansas may have been aspen forest or parkland. Few data are available for the Woodfordian vegetation of western Kansas and Nebraska and eastern Colorado. An open parkland dominated by grasses and sage, with pine and aspen.confined to protected sites, is hypothesized. During the Woodfordian-Holocene transition (12,000-9,000 yrs B.P.), mixed spruce-deciduous forest may have briefly expanded, perhaps even touching the Black Hills. By 9,000 yrs B.P., grasslands had reinvaded the entire region, leaving a few disjunct arboreal populations scattered over the region. Changes in the central Great Plains Holocene climate and vegetation have proved more subtle and less discernible in the pollen record.
Over the last five years (1980-85), 19 late Quaternary paleobotanical records in the central Great Plains have come to light (fig. 1; table 1). Much of the data, occurring as single macrofossil, charcoal, or pollen samples, has not been available in recent syntheses (Bradbury, 1980; Wright, 1981; Watts, 1983; Axelrod, 1985; Baker and Wain, 1985). Other analyses of more promising localities are yet underway. In this short review, we will present preliminary results from these unpublished localities and, in the context of previously published data, offer some hypotheses on the chronology and geography of late Quaternary vegetation. The data will be presented chronologically, from oldest to most recent, as follows: pre-Wisconsinan, early Wisconsinan (Altonian and Farmdalian), Woodfordian (late Wisconsinan), Woodfordian-Holocene transition, and Holocene.
We thank H. E. Wright and J. Groeger for their comments and for allowing us to report previously unpublished data. Additional unpublished data was generously given by J. B. Swinehart, W. J. Wayne, and L. D. Martin. We also thank Linea Sundstrom for her diligent editing.
Before the data are reviewed, several factors in the interpretation of Great Plains pollen assemblages warrant consideration. Any interpretation of pollen assemblages as vegetational reconstruction, no matter how statistically sophisticated, must be based upon appropriate analog studies of modern vegetation and pollen. In the central Great Plains, pre-Holocene climatic and historic biogeographic contexts theoretically may have resulted in the encroachment of a variety of forest or steppe vegetational associations (Axelrod, 1985). In this region, the appropriate modern analogs for fossil assemblages, if any, are not always readily apparent. Some useful studies of modern pollen and vegetation in and peripheral to the region include Potter and Rowey (1960), Potter (1967), and Hall (1985) in the Southwest; McAndrews (1966), Janssen (1966, 1967, 1984), Mott (1969), and Lichti-Federovitch and Ritchie (1968) along the boreal to eastern-deciduous forest transition; Webb and McAndrews (1976), Peterson (1978), Webb et al. (1981), and Delcourt et al. (1984) in the eastern deciduous forests to the Great Plains . border; and Kapp (1965) and McAndrews and Wright (1969) in the Great Plains. Even with adequate modern vegetational and pollen data for comparison, intuitive interpretations of fossil data may be misleading (Davis, 1963; Janssen, 1970). In this paper, in which preliminary percentage data from a variety of depositional environments are being compared, much of the vegetational reconstruction is still conjecture.
Additionally, in the central Great Plains region where ideal wet depositional sites are rare, differential pollen preservation is a problem. Modern analogs are a basis for late Quaternary environmental reconstruction only where pollen deterioration has not significantly biased the information content of the fossil pollen assemblage (Delcourt and Delcourt, 1980, p. 215). In Kapp's (1965, 1970) analysis of pre-Wisconsinan Great Plains localities with poor pollen preservation, pollen assemblages were either reported as raw counts or prefaced with a warning that reported percentages were questionable. As other studies of pollen preservation have shown, Kapp's concern was well founded (Cushing, 1967; Delcourt and Delcourt, 1980; Hall, 1981). The fact that not all pollen taxa deteriorate uniformly is firmly and empirically established (Sangster and Dale, 1964; Havinga, 1967; Holloway, 1981). Differential preservation has been shown to be responsible for tremendous over representation of Pinus (Hall, 1985) in some situations, while elsewhere rendering Populus invisible (Cushing, 1967) in the pollen record. Poor pollen preservation is the limiting factor for many of the late Quaternary records reviewed in this paper.
Over the last decade several revelations have changed the way in which pre-Wisconsinan Great Plains chronology and stratigraphy are approached. First, the Pearlette ash was recognized in fact to represent multiple ash falls spaced over more than a million years (Boellstorf, 1978). Secondly, using the deep-sea stable-oxygen-isotope record as a model, the sequence of major climatic perturbations potentially affecting the region is far more complex than the four-stadial model traditionally presumed. The pre-Wisconsinan loess and paleosol sequence documented at the Eustis ash pit, Nebraska, is believed to reflect this climatic complexity (Fredlund, Johnson, and Dort, 1985).
The primary palynological data for pre-Wisconsinan central Great Plains vegetation and climate were published prior to these changes in stratigraphic and chronological assumptions (Kapp, 1965, 1970). The chronologie and stratigraphic placement of these pre-Wisconsinan pollen records has not been reconciled with the new chronological model. Nevertheless, Kapp's research remains a landmark. Kapp's work first established that major phytogeographic shifts from forest to steppe conditions were not unique to the Wisconsinan-Holocene transition (Kapp, 1970).
Of the sites investigated by Kapp (1965, 1970), those which produced the best preserved pollen seem to represent glacial conditions with cooler temperatures, more effective precipitation, and open coniferous forest. These sites include Berends Draw and Doby Springs in northwestern Oklahoma and the Adams locality (Butler Springs sites), Meade County, Kansas (fig. 1, table 2). At both of the Oklahoma sites, Pinus was the most abundant pollen taxon, typically comprising about 50% of the pollen sum. Picea pollen also occurred consistently in significant amounts (2-10%) at these sites. The Doby Springs site also contained a small but significant number of Pseudotsuga pollen. Poaceae and Asteraceae types, excluding Artemisia and Ambrosia, were the most important nonarboreal pollen (NAP) taxa. Collectively, the NAP typically contributed more than 50% of these pollen assemblages. The pollen assemblages from the Adams locality differed slightly. At this site Artemisia pollen was consistently more common (ca. 20%), while Pinus percentages were lower (ca. 30%). At this site, like the others, Pinus and Picea pollen frequencies seem to indicate regional presence of these taxa. Based on the occurrence of Pseudotsuga at the Doby site and the geographical context of the sites, Kapp (1965, 1970) hypothesized that open, western needle-leaf forest extended into the southern High Plains during Illinoian time.
Figure 1--Location map of late Quaternary pollen botanical sites in the central Great Plains; letter codes of sites refer to table 1.
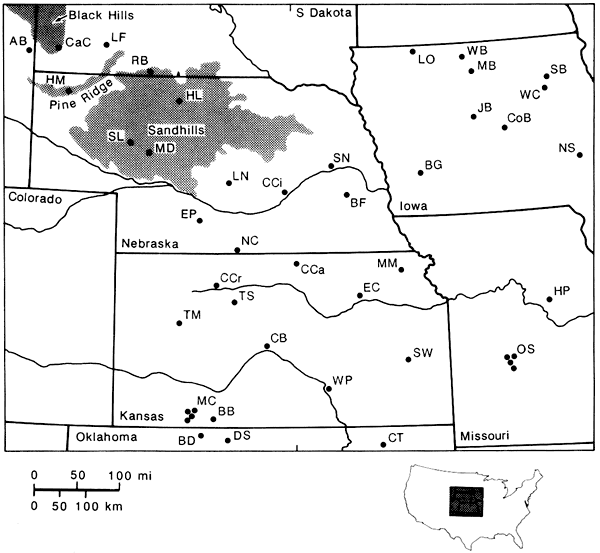
Table 1--Key to map of late Quaternary botanical/pollen sites in the central Great Plains.
| Map Symbol |
Site name and state | Depositional Environment |
Time range | Oldest and youngest C14 dates in yrs B.P. and material dated |
Reference |
|---|---|---|---|---|---|
| AB | Agate basin, WY | alluvial/paleosol | early Holocene | 11,450±110, 10,430±670, charcoal | Beiswenger, 1982 |
| BB | Big basin/Little basin, KS | karst sink, colluvium | late Holocene | 535±130, organic sediments | Shumard, 1974 |
| BD | Berends Draw, OK | karst sink, lacustrine | pre-Wisconsinan | Kapp, 1965, 1970 | |
| BF | Barteck farm, NB | prairie fen | pre-Wisconsinan | Fredlund et al., in press | |
| BG | Brayton gravel pit, IA | alluvial | Woodfordian | 12,420±420, wood and plant fragments | Baker et al., 1980 |
| CB | Cheyenne Bottoms, KS | lacustrine/marsh | Wisconsinan? | Fredlund. unpublished field notes | |
| CaC | Capes' Cave, SD | alluvial | late Holocene | 230±160, modern charcoal | Fredlund, Weston, and Mandel, 1985 |
| CCa | Courtland canal, KS | loess/colluvial | Woodfordian | 14,450±140, charcoal | Wells and Stewart, this vol. |
| CCi | Central City, NB | loess/colluvial | Woodfordian | 19,640±230/240, charcoal | Martin, p.c., 1986 |
| CCr | Coon Creek, KS | colluvial/loess? | Woodfordian | 17 ,930±550 bone collagen | Wells, 1983 |
| CoB | Colo bog, IA | bog | Woodfordian, Holocene | 13,775±300, 3,100±300, peat | Brush, 1967 |
| CT | Cross Timbers, OK | rock shelters, alluvial/colluvial | late Holocene | 1,980±75, 70±55, charcoal, organics | Hall, 1982 |
| DS | Doby Springs, OK | karst/springs | pre-Wisconsinan | Kapp, 1965, 1970 | |
| EC | Elbow Creek, KS | alluvial/paleosol | late Holocene | 1,580±70, charcoal | Kurmann, 1985 |
| EP | Eustis pit, NB | loess/paleosol | pre-Wisconsinan to Holocene | Fredlund, Johnson, and Dort, 1985 | |
| HL | Hackberry Lake, NB | interdunal lake | Holocene | 5.040±95, 1,110±75 gyttja | Sears, 1961 |
| HM | Hudson-Meng, NB | colluvial | early Holocene | 9,820±160, 8,990±190, charcoal and bone | Agenbroad, 1978 |
| HP | Hinkson-Perche, MO | pond/alluvial | late Woodfordian | 12,430±80, wood | Schmits, 1985 |
| JB | Jewell bog, IA | bog | Woodfordian, Holocene | 10,640±270, 9,570±180 peat | Brush, 1967 |
| LF | Lange-Ferguson, SD | marsh/colluvial | late Woodfordian, early Holocene | 10,730±530, bone collagen | Fredlund, 1985 |
| LN | Litchfield, NB | alluvium | Farmdalian (?) | Rogers, in Voorhies and Corner, 1985 | |
| LO | Lake Okoboji, IA | lacustrine | Woodfordian, historic | 13,990±13'5, 390±55, gyttja | Van Zant, 1979 |
| MB | McCulloch bog, IA | bog | Woodfordian, Holocene | 14,500±340, 3,170±190, peat | Brush, 1967 |
| MC | Meade County, KS (Adams Spring, Cragin Quarry, Hart Draw, Mount Scott, Jinglebob) | karst sinks, springs/alluvium | pre-Wisconsinan except Jinglebob Altonian (?) | Hibbard, 1970; Kapp, 1965, 1970 | |
| MM | Muscotah Marsh, Arrington Marsh, KS | bogs/marshes and alluvial | Farmdalian, Holocene | 24,500±800, 5,100±250, wood, peat | Grueger, 1973 |
| MD | McPherson Co. Drill Hole, NB | lake? | Farmdalian | 32,130±280/1,520, diatomaceous peat | Swehart; p.c., 1986; Wright, p.c., 1986 |
| NC | North Cove, NB | spring deposits | Woodfordian | 14,770±100, wood | Johnson et al., 1986 |
| NS | Nichols Silt, IA | alluvial | late Woodfordian | 11,800±200, wood | Baker et al., 1980 |
| OS | Ozark Springs, MO (Boney Spring, Jones Spring, Kirby Spring, Koch Spring, Trolinger Spring) | springs on alluvial terrace | Farmdalian, Woodfordian, some late Holocene | 39,020±2,600, 1,900±80, wood, peat, charcoal | Mehringer et al., 1970; King, 1973 |
| RB | Rosebud site, SD | interdunal marsh, alluvium | Woodfordian, Holocene | 12,630±160, 12,580±160, peat, plant fragments | Watts and Wright, 1966 |
| SB | Sumner bog, IA | bog | early Holocene | 9,270±90, 5,520±70,peat | Baker et al., 1980 |
| SL | Swan Lake, NB | interdunal lake | early Holocene to present | 8,950±160, 3,680±70, gyttja | Wright et al., 1985 |
| SN | Schuyler, NB | loess, colluvial | Woodfordian (?) | Wayne, p.c., 1984 | |
| SW | Sanders's well, KS | spring-fed bog | Farmdalian, Woodfordian | 23,740±340/350, 12,820±220, muck | Fredlund and Johnson, 1985; Fredlund and Jaumann, 1986 |
| TM | 12 Mile Creek, KS | alluvium | early Holocene | 10,435±260, 10,245±335, bone apatite, collage | Rogers and Martin, 1984 |
| TS | Trapshoot site, KS | loess/paleosol | Woodfordian (?) | Stewart and Rogers, 1984 | |
| WB | Woden bog, IA | bog | late Woodfordian, Holocene | 11,570±330, 2,830±(?), gyttja | Durkee, 1971 |
| WC | Wapsipinicon cutbank, IA | bog | Farmdalian, early Woodfordian | 34,500±2,000/2,760, 20,850±450, peat | Mundt and Baker, 1979 |
| WP | Wichita peat, KS | bog(?)/alluvial | Woodfordian | 19,340±200/210, peat | Rogers and Martin, 1985; Jaumann et al., 1985 |
The interpretation of pollen from sites believed to be Sangamon in age (Cragin quarry, Hart Draw, and Mount Scott localities, Meade County, Kansas) is somewhat hampered by poor preservation (Kapp, 1965, 1970). At Hart Draw where preservation was best, Kapp reported high NAP (greater than 70%) and low Pinus (less than 15%). He noted that the primary difference between these fossil assemblages and modern grassland assemblages is that Chenopodiaceae/Amaranthaceae pollen is significantly lower in the fossil record. The signal from other "Sangamon" samples is less consistent. It was found that as pollen preservation and abundance decreased, Pinus percentages tended to increase, reaching over 90% in some Mount Scott and Cragin quarry samples. Kapp (1970, p. 153) attributed this to differential pollen degradation and easy recognition of the distinctive bisacate Pinus morphology (cf. Hall, 1981, 1985). Where pollen was well preserved, Kapp's analysis clearly indicates a "Sangamon" grassland steppe.
An additional pre-Wisconsinan record comes from the Bartek Brothers farm locality in eastern Nebraska (Fredlund et al., n.d.), where fossil-bearing sedge-peat beds lie above a weathered till and below multiple loesses, paleosols, and alluvial deposits. The pollen assemblages from this peat are dominated by NAP taxa (greater than 70%), with Poaceae, Petalostemum, Ambrosia, and other Asteraceae types being among the most significant. Quercus and an assortment of other deciduous taxa dominate the AP assemblage, while Pinus and Picea occur relatively infrequently. This mid-Pleistocene pollen record is interpreted as an open grassland similar to that of the eastern Great Plains today.
Sparse but consistent evidence indicates that the Altonian, Farmdalian, and Woodfordian substages of the Wisconsinan were accompanied by major climatic and biogeographical shifts in the central Great Plains. Recently Baker and Waln (1985) used pollen and paleontological data to argue for mid-Wisconsinan (Farmdalian) grasslands in the region. Outside the Central Plains, in south-central Illinois (Grueger 1972a, 1972b), and in the Southeast (Delcourt, 1980), parallel palynological evidence for climatic substages of the Wisconsinan exists. The few previously unreported records offered here support this hypothesis.
The only Great Plains record spanning all three of these substages comes from the Llano Estacada to the south of our area of focus (Hafsten, 1961, 1964; Oldfield and Schoenwetter, 1975). The earliest Wisconsinan (Terry Pluvial or pre-Tahoka) records from this area indicate that open pine and spruce forest characterized the southern High Plains during much of this period. From about 33,500 to 22,500 yrs B,P., open Artemisia steppe replaced these forests. During the late Wisconsinan (Tahoka Pluvial), pine and spruce forest again became the dominant regional vegetation. The exact physiognomy of these southern High Plains Wisconsinan forests--whether savannas, parklands, woodlands, or full forests--is not clear (Bryant and Holloway, 1985).
A very similar early Wisconsinan (Altonian?) record comes from the Jinglebob site, Meade County, Kansas (Kapp, 1970). High percentages (ca. 50%) of Pinus pollen indicate that pine was at least regionally present. Also reported are a variety of eastern deciduous-tree pollen taxa in trace amounts (Hibbard, 1955, 1970; Kapp, 1965, 1970). The age of this locality is based solely on stratigraphy and vertebrate fossils.
From McPherson County in the Sandhills of Nebraska, a diatomaceous-peat sample recovered from a depth of approximately 200 ft (60 m) below the modern surface of an interdunal site has been radiocarbon dated to 32,130±1,280/1,520 yrs B.P. (James Swinehart, Conservation and Survey Division, Nebraska, personal communication, 1986). A pollen analysis of this peat by Linda Shane of Limnological Research Center, University of Minnesota, indicates a grassland (H.E. Wright, personal communication, 1986). NAP contributed 88% of the nonaquatic pollen sum in this sample. Poaceae, Artemisia, and Cheno-Ams were the dominant pollen taxa. Pinus and Picea pollen comprised only about 7%, indicating a distant source. This is at present the only radiometrically dated early Farmdalian pollen assemblage from the region.
Rogers (in Voorhies and Corner, 1985) offers a preliminary pollen assemblage from the Litchfield vertebrate faunal site of central Nebraska. Although poor pollen preservation prevented reliable quantitative interpretation, the presence of Ambrosia, Poaceae, Pinus, and three other AP taxa were noted. Absent are Picea or other boreal indicators expected in later, Woodfordian sites. The absolute age of this site is yet to be determined. Voorhies and Corner (1985), noting a diversity of reptiles and amphibians and relatively few individuals of boreal-indicative small mammals, suggested an environment less severe than the Woodfordian full-glacial. Certainly Rogers's pollen work supports this. We hypothesize here that the Lichfield site may date either to the Farmdalian or Farmdalian-Woodfordian transition.
The best indications that the middle Wisconsinan climate and vegetation were significantly different from the Woodfordian comes from records which span the Farmdalian-Woodfordian boundary (24,000 to 21,000 yrs B.P.). The Ozark Springs records (Mehringer et al., 1970; King, 1973) indicate that jack-pine (Pinus banksiana) parkland or savanna was overrun by spruce (Picea glauca) forest during this climatic shift. Farmdalian-Woodfordian transitional pollen records from east-central Iowa show a similar shift from jack-pine with spruce to spruce forests (Mundt and Baker, 1979; Hallberg et al., 1980; VanZant et al., 1980).
At the Muscotah and Arrington marshes in northeastern Kansas, this transition is not as well documented (Grueger, 1973). Below the 24,500-yrs-B.P. radiocarbon-dated level at Arrington, NAP, especially Poaceae and Asteraceae types, increases significantly while Picea decreases to 30%. At Muscotah, Picea percentages drop substantially below the 23,000-yrs-B.P. radiocarbon-dated level. Most enlightening is the pollen assemblage recovered from organic-rich sands below those reported on the Muscotah diagram (Grueger, personal communication, 1986). This assemblage believed to represent Farmdalian conditions has an AP:NAP ratio of aproximately 50:50. In this sample, Picea was almost absent, but the percentage of Quercus was significantly higher than in full-glacial samples. Because this pollen assemblage came from sands, rather than peat, the age and affinity were uncertain. Current investigations at Sanders's well (discussed below) support the interpretation of this as a Farmdalian assemblage. The Muscotah Farmdalian assemblage is hypothesized to represent a mosaic of deciduous forest and prairies in northeastern Kansas.
One of the most significant Wisconsinan sites now under investigation is Sanders's well, Coffey County, Kansas. As at Muscotah, the Farmdalian record (radiocarbon-dated at 23,740 yrs B.P.) indicates a mosaic of prairie and oak-hickory forest. NAP characteristic of prairie (Poaceae, Ambrosia, and other Asteraceae types) comprises more than 70% of the pollen sum exclusive of local, wet-site taxa. Quercus, Carya, Ulmus, and Populus are the most abundant AP taxa in the Farmdalian zone of the Sanders's well record. The overlying Woodfordian pollen zone indicates that an aspen parkland replaced the Farmdalian grassland at this site. This pollen record is presented more fully in the Woodfordian discussion below.
These few Farmdalian pollen records are further supported by the geomorphological, paleosol, and phytolith record from the Eustis ash pit in south-central Nebraska (Fredlund, Johnson, and Dort, 1985). In this loess record is a mid-Wisconsinan period during which eolian deposition slowed or halted (cf. Frye et al., 1968; Ruhe, 1983). In central Nebraska, this zone, the Gilman Canyon Formation (Dreeszen, 1970), is a minor Farmdalian paleosol development in early Wisconsinan loess. Opal phytolith assemblages from this Farmdalian paleosol contain tremendous quantities of panicoid forms suggesting an abundance of moist, temperate-adapted tall grasses.
Several newly discovered sites with Woodfordian macrofossil and pollen evidence have caused us to modify our working hypothesis of regional vegetation during this period (Fredlund and Jaumann, 1986). Prior to our investigations at these sites we assumed that white-spruce (Picea glauca) forest characterized the late-Wisconsinan vegetation in the central Great Plains. Certainly, Muscotah and Arrington marshes (Grueger, 1973), the Ozark Springs sites (Mehringer et at., 1970; King, 1973), and numerous sites in Iowa (Brush, 1967; Baker et al., 1980) indicated that spruce-dominated coniferous forests were the rule to the north and east of the region during the last glacial maximum (figs. 1 and 2). Also well documented is that the range of white spruce during this time extended as far south as northern Louisiana and possibly Texas (Delcourt and Delcourt, 1985; Holloway and Bryant, 1984; Bryant and Holloway, 1985) and as far west as the Nebraska Sandhills, if not farther (Watts and Wright, 1966). Therefore with some surprise and consternation we discovered two Woodfordian sites in eastern Kansas in which Picea pollen frequencies were much lower than anticipated.
Figure 2--Comparison of late Wisconsinan (Woodfordian) pollen assemblages from the central Great Plains; + trace (less than 1% ), * pollen taxa present, insufficient pollen preservation for quantitative analysis. Group A generalized from previously published data; group B from previously unpublished data.
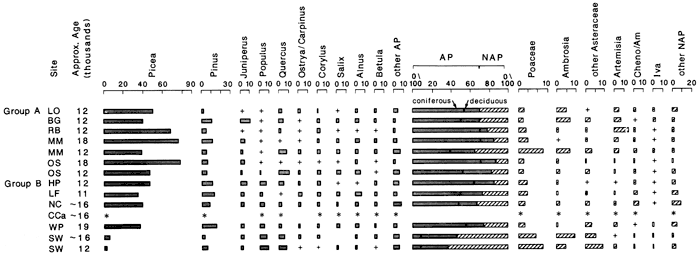
Table 2--Identified pollen taxa and possible representative modern species and their common names.
| Pollen taxa | Representative species | Common name |
|---|---|---|
| AP (trees and large woody shrubs) | ||
| Picea | P. glauca (Moench.) Voss. | white spruce |
| P. mariana (Mill.) BSP. | black spruce | |
| Abies | A. balsamea (L.) Mill. | balsam fir |
| Larix | L. laricina (DuRoi) K. Koch | tamarack |
| Pinus | P. ponderosa Laws | ponderosa pine |
| P. flexilis James | limber pine | |
| P. banksiana Lamb | jack pine | |
| Juniperus | J. virginiana L. | red cedar |
| J. communis L. | common juniper | |
| J. horizontalis Moench. | creeping juniper | |
| Populus | P. tremuloides Michx. | quaking aspen |
| P. balsamifera L. | balsam poplar | |
| P. deltoides Marsh | cottonwood | |
| Salix | S. discolor Muhl. | pussy-willow |
| S. candida Fluegge | sage-leaf willow | |
| S. interior Rowlee | sand-bar willow | |
| Betula | B. papyrifera Marsh | paper birch |
| B. glandulosa Michx. | swamp birch | |
| B. nigra L. | river birch | |
| Corylus | C. cornuta Marsh | beaker hazelnut |
| C. americana Walt. | hazelnut | |
| Alnus | A. rugosa (DuRoi) Spreng | speckled alder |
| A. serrulata (Ait.) Willd. | smooth alder | |
| Myrica | M. gale L. | sweet gale |
| M. cerifera L. | wax-myrtle | |
| Quercus | Q. macrocarpa Michx. | bur oak |
| Q. alba L. | white oak | |
| Juglans | J. cinerea L. | butternut |
| J. nigra L. | black walnut | |
| Fraxinus | F. nigra Marsh | black ash |
| F. pennsylvanica Marsh | green ash | |
| Acer | A. saccharinum L. | silver maple |
| A. saccharum Marsh | sugar maple | |
| A. negundo L. | boxelder | |
| Ostrya/Carpinus | O. virginiana (Mill.) Koch. | ironwood |
| C. caroliniana Walt. | hornbeam | |
| Ulmus | U. rubra Muhl. | slippery elm |
| U. americana L. | american elm | |
| U. thomasi Sarg. | rock elm | |
| Carya | C. tomentosa (Poir.) Nutt | mockernut |
| C. glabra (Mill.) Sweet | pignut hickory | |
| Rhus | R. glabra L. | smooth sumac |
| R. radicans L. | poison oak | |
| Comus | C. drummondii C. A. Mey | rough-leaved dogwood |
| C. racemosa Lam. | gray dogwood | |
| Shepherdia | S. canadensis (L.) Nutt. | buffalo berry |
| Elaeagnus | E. commutata Bernh. | silver berry |
| Sambucus | S. canadensis L. | common elder |
| NAP (nonarboreal pollen) | ||
| Ambrosia | A. trifida L. | giant ragweed |
| A. tomentosa Nutt. | bursage | |
| Artemisia | A. filifolia Torr. | sand sagebrush |
| A. tridentata Nutt. | big sagebrush | |
| Iva | I. xanthifolia Nutt. | marsh elder |
| Helianthus | H. annuus L. | sunflower |
| Asteraceae | (other members of the aster family) | asters, goldenrod, thistle, groundsel |
| Poaceae | (the grass family) | grass |
| Cheno/Am | (the Chenopodiaceae and Amaranthaceae families) | goosefoot, lamb's quarters, amaranth |
| Petalostemum | Dalea purpurea Vent. | prairie clover |
One of these sites is in the Arkansas River floodplain near Wichita, Kansas. Bones of Pleistocene fauna (Rogers and Martin, 1985), spruce wood, and balls of woody peat containing Picea glauca needles, twigs, cone fragments, and other macro- and microfossils are being recovered from a depth of more than 30 ft (9 m) below the surface. One of these spruce-bearing peat samples has yielded a radiocarbon date of 19,340±200/210 yrs B.P. Although the stratigraphy of this site is not yet fully known, we believe these Pleistocene materials to be of local origin. Pollen of aquatic plants indicating open, still-water deposition is common in the peat samples. Although alluvial transport and mixing in this floodplain depositional environment remain a possibility, we argue that the pollen and macrofossils contained in the peat samples accurately reflect the local and regional Woodfordian vegetation of the site.
Characteristic pollen assemblages of the Wichita Woodfordian peat are high in AP (fig. 2). Although Picea is the most common AP taxa in this peat, its relative frequency (35%) is far less than that found at the Ozark Springs or Muscotah Marsh localities during the approximate same time. Pinus percentages seem to indicate a limited presence of the taxon. A diversity of deciduous AP taxa is also identified, including Populus, Quercus, Salix, Ostrya/Carpinus, Fraxinus, Alnus, Acer, Betula, Corylus, Ulmus, Carya, and Comus. The frequency of occurrence for any one of these taxa seems relatively inconsequential; however, taken together they represent a significant constituent of the fossil-pollen assemblage. We interpret this arboreal-dominated pollen spectrum as a mixed, coniferous-deciduous forest in the mesic Arkansas River valley. The substantial NAP portion of the Wichita pollen record, principally Poaceae, Ambrosia, Artemisia, Iva, and Petalostemum, gives the record an open-forested aspect.
During the 1936 excavation of a well in a small, upland, spring-fed bog, Mr. Albert Sanders unearthed the partial remains of a mammoth. Recent reinvestigation of this site, positioned along a first-order tributary above the Neosho River, Coffey County, Kansas, documents a 2.6-m (8.6-ft)-thick organic-rich layer of muck underlain by sands. The late Pleistocene portion of this section lies below an apparent erosional unconformity 1.6 m (5.3 ft) below the surface. Two radiocarbon dates, 12,820±220 and 23,740±330/350 yrs B.P., bracket this 1-m (3-ft) portion of the section.
Pollen preservation in these muck deposits is exceptional. Estimates of pollen concentration are typically 200,000 to 400,000 grains per cm3. Changes in relative frequencies of pollen taxa reflect good zonation within the Wisconsinan portions of the section. The lowest pollen zone, representative of Farmdalian conditions, has been discussed above. Like the Farmdalian zone, the Woodfordian zone is dominated by NAP. The most important of these NAP taxa are Poaceae (22%), Ambrosia (26%), and a variety of other Asteraceae types (14%). Both Cheno-Am types and Artemisia are relatively rare in these sediments. Quercus (8%) and Populus cf. tremuloides (7%) are among the most common AP taxa. Pinus and Picea occur relatively less frequently (each less than 5%) and indicate distant, rather than local, presence of these trees. Myrica pollen is also common in the Woodfordian zone. The unusually high amounts (up to 24%) of Myrica indicate a local presence of this wet-loving shrub. This record indicates aspen parkland or groveland was the dominant Woodfordian vegetation of the uplands surrounding the Sanders's well locality.
The Sanders's well record is unique. Although the site lies between the documented occurrences of spruce at Wichita and Muscotah Marsh, spruce seems to be absent in the vicinity of Sanders's well. In view of the Sanders's well record, the relatively high NAP and Populus pollen percentages at Wichita probably also represent aspen parklands. Two factors are hypothesized to be responsible for the observed differences among Woodfordian sites in eastern Kansas. Most important is the unique topographic situation of the Sanders's well site. It is located in gently rolling, interfiuvial uplands, rather than within a river valley. Fire protection and a more mesic micro-climate allowed mixed spruce and deciduous forest to dominate in the river valleys, while the exposed, edaphically poorer uplands were burned frequently enough to prevent succession to spruce forests.
The second hypothesized reason for the differences among central Great Plains Woodfordian pollen records is climate. It has been proposed that major Woodfordian airmass boundaries were centered over the region (Bryson, 1966; Bryson and Wendland, 1967; Bryson et al., 1970). Such a boundary would create a northeast-southwest moisture and temperature gradient across the central Great Plains analogous with that currently in the Manitoba and Minnesota region (Borchert, 1950). The evidence from Wichita and other localities south and west of Muscotah all indicate a more open and mixed forest during the Woodfordian.
The Courtland Canal in north-central Kansas provides one of these mixed spruce-deciduous forest records (figs. 1 and 2). The Courtland Canal locality deposition is reported to be loess, with some colluviation possible. Picea charcoal from the site has been dated to 14,450±140 B.P. (Wells and Stewart, this volume). Pollen has been recovered from a sample of the dated horizon, but unfortunately is poorly preserved. Only one out of every four or five grains was identifiable; however, Picea and a variety of AP taxa have been identified (fig. 2). The diversity of pollen types with the presence of Picea qualitatively resembles the assemblages recovered from the Wichita peat and the North Cove locality discussed below.
The North Cove site, exposed along the Harlan County Lake on the loess plain of south-central Nebraska, has produced a variety of paleoenvironmental data (Johnson et al., 1986; Wells and Stewart, this volume; Stewart, this volume). Persistently flowing springs seem to be responsible for the excellent preservation of botanical macrofossils and pollen at this locality. A sample of spruce wood has yielded a radiocarbon date of 14,770±100 yrs B.P. Organic-rich sediments from this dated level have produced an assemblage of pollen very similar to that recovered at Wichita (fig. 2). Picea pollen and diverse deciduous AP comprise 68% of the pollen assemblage. Populus cf. tremuloides is again among the most common deciduous AP. Pollen of woody shrubs including Elaeagnus, Shepherdia canadensis, and Sambucus occurs in very low relative frequencies.
The Woodfordian vegetation of eastern Nebraska is not yet fully documented. So far only a few charcoal-bearing sites are known. Picea charcoal from a Peorian loess section near Central City in Merrick County, Nebraska, has produced a 19,640±230/240 yrs B.P. date (Larry D. Martin, personal communication, 1986). A similar section from near Schuyler in Colfax County also has produced Picea charcoal (William Wayne, personal communication, 1984). A radiocarbon date from this locality is still pending. Our attempts to extract pollen from these localities have been unsuccessful. It is anticipated that when found, full-glacial Woodfordian pollen from eastern Nebraska will be Picea-dominated with less deciduous diversity than documented at the North Cove site. Both these spruce-charcoal sites and the macrobotanical and pollen sites from central Nebraska and Kansas indicate that some forest cover was present in the region during Peorian loess deposition.
One hypothesized source of Woodfordian (Peorian) loess-forming silts is the Sandhills of central Nebraska (Fredlund, Johnson, and Dort, 1985). The Woodfordian vegetation of the Sandhills themselves is not documented. The major period of dune formation is generally accepted to have occurred during the Woodfordian with later (Holocene) sand movement being secondary (Wright et al., 1985). The development and perpetuation of this massive dunefield would have severely retarded the vegetational succession (Wright, 1970; Bradbury, 1980; Wright et al., 1985); however, we believe that this does not necessarily translate into a treeless landscape. Within the boreal forest today, active, albeit smaller, dune fields encroach on interdunal areas of open-coniferous forest (e.g., Raup and Argus, 1982). Pollen evidence for the Woodfordian vegetation both south and east of the Sandhills indicates that the regional climate would have supported open spruce or mixed spruce-deciduous forest probably with large areas of aspen parkland. Until Pleistocene botanical fossil-bearing interdunal deposits are found, the Woodfordian vegetation of the Nebraska Sandhills will remain uncertain.
In south-central South Dakota on the northern edge of the Sandhills, the Rosebud locality provides the critical record of late Woodfordian (ca. 13,000-12,000 yrs B.P.) vegetation (Watts and Wright, 1966). Picea was by far the most abundant AP taxon recorded. A diversity of deciduous AP taxa was present but lower in overall relative frequency than at the North Cove site (fig. 2). Another difference between these two localities is the significant occurrence of Artemisia pollen at Rosebud. None of the other Woodfordian central Great Plains records discussed above included high frequencies of this NAP taxon. It is hypothesized that the unique edaphic situation of the Sandhills may be responsible for the higher Artemisia percentages at Rosebud (Watts and Wright, 1966).
That area of the region least understood is western Kansas and Nebraska and eastern Colorado. We currently do not have evidence for the western limits of Picea glauca or any of the deciduous arboreal taxa present in eastern Kansas and central Nebraska during the Woodfordian. Based on paleontological data, the western Kansas and eastern Colorado region has been hypothesized to have been an open-pine savanna or parkland during the last glacial maximum (Guilday, 1964; Hoffmann and Jones, 1970; Martin and Neuner, 1978; Graham, 1979; Martin and Hoffmann, this volume). Unfortunately, Woodfordian botanical data from this region are scarce. The palynological evidence from the Llano Estacado (Oldfield and Schoenwetter, 1975), as well as that from the few recently investigated localities, supports the conifer-parkland hypothesis.
J. D. Stewart and P. V. Wells (Wells, 1983; Wells and Stewart, this volume) recently recovered needles of Pinus flexilis from highly calcareous loess or colluviated loess deposits at the Coon Creek paleontological site, Graham County, Kansas. Bone collagen from this deposit has produced a date of 17,930±550 yrs B.P. Our first attempt to extract pollen from these sediments yielded mixed results. Pinus pollen was present but too poorly preserved to identify to subfamily or genera. Also present were a large number of insect-pollinated taxa, which further confused the quantitative results. These insect-pollinated types are suspected to have been emplaced by burrowing bees or other insects and probably are not contemporaneous with the other fossils at this site.
Palynological investigations at the Trapshoot site, Rooks County, Kansas, another Wisconsinan paleontological site, have produced similar results (Stewart and Rogers, 1984). Probably because of poor preservation, only four pollen taxa were identified from this site: Pinus, Poaceae, Asteraceae types (primarily Ambrosia), and Cheno-Am types. From the presence of 18% Pinus in this assemblage, Stewart and Rogers argued that pines probably were present in limited stands in the region of the site; however, poor preservation makes such quantitative interpretations questionable.
All the data accrued so far indicate that pine was present in the western reaches of the central Great Plains during the Woodfordian. The documented occurrence of limber pine (P. flexilis) suggests that western (montane) conifer forests were in part the source for the flora of this western Great Plains Woodfordian vegetation (Wells and Stewart, this volume). The exact physiognomy and geographical extent of this vegetation zone remain unknown. None of the records available so far indicate closed forest or even savanna. Instead, more open vegetation, with conifers and probably aspen confined to escarpments and other fire-protected sites, can be hypothesized.
One Wisconsinan site currently under investigation may help in understanding the nature of the transition from the western parklands to the open, mixed spruce-deciduous forest of eastern Kansas. With the cooperation of the Kansas Geological Survey, a 15-m (50-ft) core of lacustrine sediments from Cheyenne Bottoms in central Kansas has been obtained and is currently being analyzed at the University of Kansas Palynological Laboratory. This record should add significantly to our understanding of the history of central Great Plains vegetation.
Between about 12,000 and 9,000 yrs B.P., the climate and vegetation of central North America underwent dramatic changes (Wright, 1970; Watts, 1983; Webb et al., 1983). The pollen record from Muscotah Marsh presents the most complete record of this critical3,000-yr period in the eastern central Great Plains (Grueger, 1973). Around 12,000 yrs ago relative frequencies of Picea pollen began to fall sharply, indicating the rapid demise of the spruce forests. A diversity of deciduous AP taxa (Corylus, Salix, Quercus, Ulmus, Ostrya/Carpinus, and Fraxinus) and NAP taxa (principally Poaceae and Ambrosia) increased simultaneously. This assemblage is very similar to the open, mixed spruce-deciduous assemblages now documented for the mid-Woodfordian sites southeast of Muscotah (fig. 2). The Muscotah record indicates that by 10,500 yrs B.P., spruce had all but disappeared from the region. During this final spruce decline, deciduous trees apparently increased until about 9,000 yrs B.P. when grasslands in the region expanded significantly.
The late Woodfordian pollen assemblages from Missouri indicate that mixed spruce-deciduous forest replaced the coniferous forest early in the transition period. At Boney Springs, one of the Ozark Springs sites, this mixed forest was present by about 13,500 yrs B.P. (Mehringer et al., 1970; King, 1973). A very similar pollen assemblage (fig. 2) from the Hinkson-Perche archeological site near Columbia, Missouri, has an associated radiocarbon date of 12,430±80 yrs B.P. (Schmits, 1985). Neither of these localities has the complete late Woodfordian-early Holocene succession of vegetation.
The late Woodfordian-early Holocene transition in northern Iowa at Lake Okoboji (Van Zant, 1979) and Woden Bog (Durkee, 1971) generally parallels that recorded at Muscotah Marsh. The Iowa records, however, exhibit better zonation of deciduous AP taxa during this period. At Lake Okoboji, as Picea pollen declines, the deciduous pollen taxa that show increases are Fraxinus nigra, followed by Betula and Alnus, and finally Quercus and Ulmus prior to the 9,000-yrs-B.P. rise in grasslands. The zonation present at these Iowa localities reflects both the proximity of the deciduous taxa refugia and the differential rates of their dispersion (Wright, 1970).
No continuous records of this Woodfordian-Holocene transition exist for the western half of the central Great Plains. Even the Rosebud record is apparently broken by an unconformity (Watts and Wright, 1966). Although as a rule archeological sediments are not ideal depositional environments for pollen preservation, a number of early Holocene archeological sites have produced significant palynological records. By piecing these chronologically short archeological records with other data, an outline of the Woodfordian-Holocene transition in the western part of the region has begun to emerge. As yet many unexplained discrepancies exist among these records.
The presence of disjunct populations of Picea glauca and a variety of eastern deciduous trees indicates that these late Pleistocene forests probably extended westward at least to the Black Hills (Buttrick, 1914; Watts and Wright, 1966). Pollen extracted from late Woodfordian-early Holocene sedge-marsh sediments from the Lange-Ferguson paleoindian mammoth-kill site, along the Pine Ridge escarpment in the White River Badlands of South Dakota, supports this hypothesis (Fredlund, 1985). Although pollen preservation was mixed and pollen concentration relatively low (3,000 grains/gram sediment), the pollen diversity and consistency with other records warranted quantitative representation and interpretation of the data (fig. 2). The pollen indicates that at the time of the mammoth kill (circa 11,000 yrs B.P.), the vegetation was an open-spruce forest with a diverse understory of small trees and shrubs including Populus, Juniperus, Alnus, Betula, Corylus, Salix, Shepherdia, and Ostrya/Carpinus. All of these taxa except Alnus are extant in the Black Hills approximately 80 km (48 mi) northeast of the site. This record is similar to many of the Pleistocene-Holocene transition sites of the northern and eastern portions of the central Great Plains (fig. 2).
How widespread such forests were in the late Woodfordian western Great Plains is unclear. Recently published data from the Powder River basin, Wyoming, west of the Black Hills, indicate that steppe vegetation and a semi-arid climate have persisted in the area since 13,000 yrs B.P. (Markgraf and Lennon, 1986). At the Hudoon-Meng early Holocene archeological site on the Pine Ridge escarpment just south of the Black Hills, fossil-pollen assemblages show no similarity to the Lange-Ferguson record (Kelso, reported in Agenbroad, 1978). The Hudson-Meng pollen record, dated at approximately 9,000 yrs B.P., indicates a grassland and Artemisia-steppe regional vegetation. No pollen evidence exists for either open spruce forests or the ponderosa pine for which this escarpment is known today. These other regional records suggest that the hypothesized open mixed spruce-deciduous forest bridge to the Black Hills was not widespread, but an ephemeral late Wisconsinan phenomenon confined to favorable topographical and edaphic situations, such as the north-facing Pine Ridge escarpment.
A palynological investigation of the Agate Basin archeological site just southwest of the Black Hills was less successful (Beiswenger, 1982). Inadequate pollen preservation, low concentration, and recycling of older fossil palynomorphs precluded quantitative interpretation.
An early Holocene (ca. 10,300 yrs B.P.) pollen assemblage relatively high in Pinus pollen has been reported from the 12 Mile Creek bison-kill site in Logan County, Kansas (Rogers and Martin, 1984). Although dominated by NAP, Pinus comprised approximately 37% of the assemblage. Quercus and Populus were reported only in trace amounts. These data were interpreted as most likely indicating the persistence of pine, possibly limber pine (Pinus flexilus), along escarpments or other protected situations in the vicinity of the site at the time of the bison kill (cf. Wells, 1965, 1970).
The southernmost central Great Plains late Woodfordian-early Holocene pollen record comes from the Domebo archeological site in central Okalahoma (Wilson, 1966). This NAP-dominated record roughly spans the 11,000-9,000-yrs-B.P. transitional period. The ca. 11,000- yrs-B.P. assemblages are dominated primarily by Poaceae (ca. 45%) and Asteraceae (ca. 25%). These increase even more after 10,000 yrs B.P. Pinus is relatively unimportant (5%) when compared to the 12 Mile Creek record. Picea pollen occurs even less frequently. Several deciduous pollen taxa, Quercus, Carya, Ulmus, and Rhus, are present but rare. Most significant, especially considering its typically low production and susceptibility to deterioration, is Populus (4-5% ). This Populus pollen may represent either local cottonwoods (P. deltoides) or the southern extent of the aspen (P. tremuloides) parklands hypothesized for the eastern Kansas Woodfordian. The Domebo pollen record indicates that grasslands dominated the vegetation of central Oklahoma in the earliest Holocene.
Changes in vegetation and climate in the central Great Plains since 9,000 yrs B.P. are expected to have been less dramatic and to have resulted in more subtle changes in the pollen record. In the Midwest, pollen evidence for Holocene vegetation change resulting from species migration as well as climatic fluctuations has been mapped in detail (Webb et al., 1983). With only a few continuous records available, these types of analyses are not yet possible for the central Great Plains. This is in part due to lack of suitable sites. Only along the eastern side of the region, where a few lakes and bogs are present and changes in forest-prairie border are detectable, is there pollen evidence for climatically significant Holocene vegetation change.
In northern Iowa, at both Woden Bog (Durkee, 1971) and Lake Okoboji (VanZant, 1979), an increase in prairie forbs occurred beginning about 7,500 yrs B.P. (Webb et al., 1983). Between 5,000 and 3,000 yrs B.P., the relative frequencies of these prairie-indicative pollen taxa decreased somewhat and the prairie-forest mosaic characteristic of the region in historic times (Kuechler, 1964) began to develop.
A similar record occurs at the Muscotah Marsh in northeastern Kansas (Grueger, 1973). Although the Holocene portions of the Muscotah Marsh profile are broken by unconformities and lack close-interval radiocarbon dates, the data clearly indicate that a mid-Holocene prairie expansion and contraction occurred in northeastern Kansas.
A late Holocene prairie-forest fluctuation has been shown in the Cross Timbers of northeastern Oklahoma (Hall, 1982). Hall employed a variety of differing proxy data sets to build a convincing argument for late Holocene climate change in the southern Great Plains. Based on snails, small mammals, and pollen, especially the record from the Big Hawk rockshelter, a more mesic period from 2,000 to 1,000 yrs B.P. followed by a significantly drier episode is hypothesized. During the wetter episode Carya pollen increased, while Poaceae pollen was relatively more frequent during the later drier period. We plan to replicate Hall's pollen record independently in the upper Holocene portions of the Sanders's well section from the Osage Hills of Kansas.
Capes's Cave, a late Holocene alluvial pollen record from the grassland-pine forest ecotone in the Black Hills, documents the historic or protohistoric (ca. 200 yrs B.P. to present) expansion of the ponderosa-pine forest (Fredlund, Weston, and Mandel, 1985). The encroachment of the pine forest into grassland steppe was probably due to a combination of climatic change and historical land-management changes resulting in control and containment of fire.
These climatically induced changes in vegetation have not been documented in the region away from sensitive grassland-forest ecotones. Even the best central Great Plains Holocene pollen records, those from the interdunal lakes in the Sandhills of Nebraska, show no climatically significant change. Sears' (1961) investigation of Hackberry Lake produced only a discontinuous, 5,000 yrs B.P. to present, record of pollen. More recently, Wright et al. (1985) have published.,a continuous 9,000 yrs B.P. to present pollen record from Swan Lake, Garden County, Nebraska. However, in both of these records variation in pollen percentages is most easily explained by local, site-specific edaphic changes. No clear, climatically induced signal in these grassland-pollen records has been identified.
Big basin and Little basin, karst sinkholes in Clark County, Kansas, also have been demonstrated to have considerable continuous Holocene deposits (Shumard, 1974). A 535±130-yrs-B.P. radiocarbon date was obtained from the base of a 3.2-m (10.6-ft) core used for pollen analysis. The minor fluctuations in the percentage pollen record do not seem to have regional climatic or vegetational significance.
Measuring climatic changes within the Holocene grasslands may prove more challenging than mapping the Pleistocene vegetation of the region. On the central Great Plains Holocene grasslands, major climatic perturbations may result only in a change of grass and forb species. To detect such subtle changes, more sensitive measures of the environment are needed. One approach to this problem is synthetic where a variety of data sets are pooled to test for significant changes (e.g. Hall, 1982). Within pollen analysis, finer resolution in prairie forb classification may also help, especially in the Asteraceae. Another approach is to combine pollen and phytolith analysis. Kurmans's (1985) investigation of the Elbow Creek paleosol is an example of the usefulness of these combined techniques. Phytolith analysis may enable the discrimination of Poaceae sub-families, which are indistinguishable in the pollen record (Twiss, this volume).
This review of central Great Plains late Quaternary pollen and botanical data has proceeded chronologically from the pre-Wisconsinan to the Holocene. The only new pre-Wisconsinan locality record confirms Kapp's (1965, 1970) conclusion that the grassland steppes of central North America are not unique to the Holocene. The general late Quaternary hypothesis holds that the grasslands expanded during the interglacials and contracted or even disappeared during the glacial maxima. Surprisingly, this effect was felt even during the Farmdalian interstadial which is often perceived as a minor climatic perturbation relative to the Holocene or Sangamon. Botanical evidence for Farmdalian grasslands is found as far east as eastern Kansas and Iowa (Baker and Waln, 1985) and as far north as the Sandhills of Nebraska.
Woodfordian data now available from the eastern side of the central Great Plains indicate that boreal forest overran the Farmdalian grasslands about 24,000-22,000 yrs B.P. and persisted until the Holocene. These hypothesized forests were a mosaic of spruce, mixed spruce-deciduous, and aspen, with open parkland more common to the south and west. If the Sanders's well record is representative, the larger part of eastern Kansas, the interfluvial uplands, were open aspen parkland or grovelands throughout the Woodfordian.
Woodfordian vegetation of the western portions of the region is poorly documented. The data do indicate that limber pine was present but in unknown amounts. Pines and other western conifers may have been confined to escarpments or other protected sites. Scattered groves of aspen probably also were present throughout western Kansas and eastern Colorado during this time. The best current hypothesis is an open parkland physiognomy, with grass or Artemisia steppe being arealy far more important than forest.
Vegetational change during the central Great Plains Holocene has been given less attention than that of the Pleistocene. The few recent investigations of localities away from grassland-forest ecotones have not been able to detect hypothesized Holocene climate variations. New tools sensitive to subtle changes in grass and forb communities must be developed to test for these Holocene climatic changes.
A great potential exists for late Quaternary palynological research in the central Great Plains, both Holocene and Pleistocene. Although the number of investigated localities in the region has more than doubled in the past five years, the area has only just begun to be explored. The pool of potentially productive palynological localities in the central Great Plains is anything but exhausted.
Agenbroad, L. D., 1978, The Hudson-Meng site--an Alberta bison kill in the Nebraska High Plains: Washington, D.C., University Press of America, 225 p.
Axelrod, D. I., 1985, Rise of the grassland biome, central North America: Botanical Review, v. 51, no. 2. p. 163-201.
Baker, R. G., and Waln, K. A., 1985, Quaternary pollen records from the Great Plains and central United States; in, Pollen Records of Late Quaternary North American Sediments, V. M. Bryant, Jr. and R. G. Holloway, eds.: Dallas. Texas, American Association of Stratigraphic Palynologists Foundation. p, 191-203.
Baker, R. G., VanZant, K. L.,and Dulian, J. J., 1980, Three late-glacial pollen and plant macrofossil assemblages from Iowa: Palynology, v. 4. p. 197 203.
Beiswenger, J. M., 1982. Pollen analysis; in, The Agate Basin Site. A Record of the Paleoindian Occupation of the Northwestern High Plains, G. C. Frison and D. J. Stanford, eds.: San Francisco, California, Academic Press, p. 349-352.
Boellstorf, J., 1978, Chronology of some late Cenozoic deposits from the central United States and the Ice Ages: Transactions of the Nebraska Academy of Sciences, v. 6. p. 35--49.
Borchen, J. R., 1950, Climate of the central North American grassland: Annals of the Association of American Geographers, v. 40, p. 1-39.
Bradbury, J. P., 1980, Late Quaternary vegetation history of the central Great Plains and its relationship to eolian processes in the Nebraska Sandhills: U.S. Geological Survey, Professional Paper 112Q-C, p. 29-36.
Brush, G. S., 1967, Pollen analysis of late-glacial and postglacial sediments in Iowa; in, Quaternary Paleoecology, E. J. Cushing and H. E. Wright, Jr., eds.; New Haven, Connecticut, Yale University Press, p. 99-115.
Bryant, V. M., Jr., and Holloway, R. G., 1985, A late Quaternary paleoenvironmental record of Texas--an overview of the pollen evidence; in, Pollen Records of Late Quaternary North American Sediments, V. M. Bryant, Jr. and R. G. Holloway, eds.: Dallas, Texas, American Association of Stratigraphic Palynologists Foundation, p. 39-70.
Bryson, R. A., 1966, Air masses, streamlines, and the boreal forest: Geographical Bulletin, v. 8, p. 228-269.
Bryson. R. A., and Wendland, W. M., 1967, Tentative climatic patterns for some late-glacial and post-glacial episodes in central North America: University of Wisconsin, Madison, Department of Meteorology, Technical Report 34, Nonr. 1202(07), 32 p.
Bryson, R. A., Baerreis, D. A., and Wendland, W. M., 1970, The character of late-glacial and post-glacial climatic changes; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr. and J. K. Jones, Jr., eds.: Lawrence, Kansas. University of Kansas Press, p. 53-74.
Buttrick, P. L., 1914, The probable origin of the forests of the Black Hills of South Dakota: Forestry Quarterly, v. 12, p. 223-227.
Cushing, E. J., 1967, Evidence for differential pollen preservation in late Quaternary sediments in Minnesota: Review of Paleobotany and Palynology, v. 4, p. 87-101.
Davis, M. B., 1963, On the theory of pollen analysis: American Journal of Science, v. 261, no. to, p. 897-912.
Delcourt, H. R., and Delcourt, P. A., 1985, Quaternary palynology and vegetational history of the southeastern United States; in, Pollen Records of Late Quaternary North American Sediments, V. M. Bryant, Jr., and R. G. Holloway. eds.: Dallas, Texas, American Association of Stratigraphic Palynologists Foundation, p. 1-37.
Delcourt, P. A., 1980, Goshen Springs--late Quaternary vegetation record for southern Alabama: Ecology, v. 61, no. 2, p. 371-386.
Delcourt, P. A., and Delcourt, H. R., 1980, Pollen preservation and Quaternary environmental history in the southeastern United States: Palynology, v. 4, p. 215-231.
Delcourt, P. A., Delcourt, H. R., and Webb, T., III, 1984, Atlas of mapped distributions of dominance and modern pollen percentages for important tree taxa of eastern North America: American Association of Stratigraphic Palynologists, Contribution Series, no. 14, 131 p.
Dreeszen, V. H., 1970, The stratigraphic framework of Pleistocene glacial and periglacial deposits in the central Plains; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr. and J. K. Jones, Jr., eds,; Lawrence, Kansas, University of Kansas Press, p. 9-22.
Durkee, L H., 1971, A pollen profile from Woden Bog in north-central Iowa; Ecology, v. 52, no. 5, p. 837-844.
Fredlund, G. G., 1985, A late Pleistocene-early Holocene pollen and phytolith record from the Lange-Ferguson site, White River Badlands, South Dakota: Sioux Falls, South Dakota, Augustana College, Archaeology Laboratory of the Center for Western Studies, manuscript on file.
Fredlund, G. G., Manin, L. D., Kreycik-Messinger, K., in press, Inconsistencies in paleoenvironmental indicators from a mid-Pleistocene site in eastern Nebraska: Nebraska Academy of Sciences, Institute for Tertiary-Quaternary Studies. TER-QUA Symposium Series. v. 2.Fredlund, G. G., Johnson, W, C., and Dort, W., Jr., 1985, A preliminary analysis of opal phytoliths from the Eustis ash pit, Frontier County, Nebraska; Nebraska Academy of Sciences, Institute for Tertiary-Quaternary Studies, TER-QUA Symposium Series, v. 1, p. 147-162.
Fredlund, G., Weston, T., and Mandel, R., 1985, Palynological evidence for a late prehistoric vegetation change in the southern Black Hills from Capes Cave (39FA205): Plains Anthropologist v. 30, no. 107, p. 9-23.
Fredlund, G. G., and Jaumann, P. J., 1986, The influence of topography and fire disturbance on late Wisconsinan vegetation in eastern Kansas (abs. ); American Quaternary Association. 9th Meeting, Champaign, Illinois. Program and abstracts. p. 81.
Fredlund, G. G., and Johnson, W. C., 1985, Palynological evidence for late Pleistocene vegetation from Sanders's well locality in east-central Kansas (abs.); Institute for Tertiary-Quaternary Studies, 4th Annual Symposium, Lawrence, Kansas, Program with Abstracts, p. I5.
Frye, J. C., Willman, H. B., and Glass. H. D., 1968, Correlation of Midwestern loesses with the glacial succession; in, Loess and Related Eolian Deposits of the World. C. B. Schultz and J. C. Frye, eds.: Lincoln, Nebraska, University of Nebraska Press, p. 3-22.
Graham, R. W., 1979, Paleoclimates and late Pleistocene faunal provinces in North America; in, Pre-Llano Cultures of the Americas--Paradoxes and Possibilities, R. L Humphrey and D. Stanford, eds.: Washington, D. C., Anthropological Society of Washington, p. 49-69.
Grueger, E., 1972a, Pollen and seed studies of Wisconsinan vegetation: Geological Society of America, Bulletin. v. 83. p. 2.715-2,734.
Grueger, E., 1972b. Late Quaternary vegetation development in south-central Illinois: Quaternary Research, v. 2. p. 217-231
Grueger, J., 1973, Studies on the late Quaternary vegetation history of northeastern Kansas: Geological Society of America, Bulletin, v. 84, p. 237-250.
Guilday, J. E., 1964, Differential extinction during late Pleistocene and Recent times; in, Pleistocene Extinctions-the Search for a Cause, P. S. Manin and H. E. Wright, eds.: New Haven, Connecticut, Yale University Press, p. 121-140.
Hafsten, U., 1961. Pleistocene development of vegetation and climate in the southern High Plains as evidenced by pollen analysis; in, Paleoecology of the Llano Estacada, F. Wendorf, ed.; Santa Fe, New Mexico, Museum of New Mexico Press, Fort Burgwin Research Center, p. 59-91.
Hafsten, U., 1964, A standard pollen diagram for the southern High Plains, U.S.A., covering the period back to the early Wisconsin glaciation: International Union Quaternary Research (INQUA), VI Congress, Warsaw, 1961, v. II, paleobotanical section, p. 407-420.
Hall, S. A., 1981, Deteriorated pollen grains and the interpretation of Quaternary pollen diagrams: Review of Paleobotany and Palynology, v. 32, p. 193-206.
Hall, S. A., 1982, Late Holocene paleoecology of the southern Plains: Quaternary Research, v. 17, p. 391-407.
Hall, S. A., 1985, Quaternary pollen analysis and vegetational history of the South west; in, Pollen Records of Late Quaternary North American Sediments. W. M. Bryant, Jr., and R. G. Holloway, eds.: Dallas, Texas, American Association of Stratigraphic Palynologists Foundation, p. 95-123.
Hallberg, G. R., Baker, R. G., and Legg, T., 1980, A mid-Wisconsinan pollen diagram from Des Moines County, Iowa: Proceedings of the Iowa Academy of Sciences, v. 87, no. 2, p. 41-44.
Havinga, A. J., 1967, Palynology and pollen preservation: Review of Paleobotany and Palynology, v. 2, p. 81-98.
Hibbard. C. W., 1955, The Jinglebob interglacial (Sangamon?) fauna from Kansas and its climatic significance: Contributions of the Museum of Paleontology, University of Michigan, v. 12, p. 179-228.
Hibbard. C. W., 1970, Pleistocene mammalian local faunas from the Great Plains and central-lowland provinces of the United States; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort. Jr., and J. K. Jones, Jr., eds.: Lawrence, Kansas, University of Kansas Press, p. 395-433.
Hoffmann, R. S., and Jones, J. K., Jr., 1970, Influence of late-glacial events on the distribution of recent mammals on the northern Great Plains; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: Lawrence, Kansas, University of Kansas Press, p. 355-394.
Holloway, R. G., 1981, Preservation and experimental diagenesis of the pollen exine: Ph. D. dissertation, Texas A & M University, College Station, Texas, 317 p.
Holloway, R. G., and Bryant, V. M., Jr., 1984, Picea glauca pollen from late-glacial deposits in central Texas: Palynology, v. 8, p. 21-32.
Janssen, C. R., 1966, Recent pollen spectra from the deciduous and coniferous forest of northeastern Minnesota--a study in pollen dispersal: Ecology, v. 47, p. 804-825.
Janssen, C. R., 1967, A comparison between the recent regional pollen rain and sub-recent vegetation in four major vegetation types in Minnesota (U.S.A.): Review of Paleobotany and Palynology, v. 2, p. 331-342.
Janssen, C. R., 1970, Problems in the recognition of plant communities in pollen diagrams: Vegetatio, v. 20, p. 187-198.
Janssen, C. R., 1984, Modern pollen assemblages and vegetation in the Myrtle Lake peatland: Ecological Monographs, v. 54, p. 213-252.
Jaumann, P., Johnson, W., and Fredlund, G., 1985, Late Wisconsin full-glacial vegetation record from south-central Kansas (abs.): Institute for Tertiary-Quaternary Studies, 4th Annual Symposium, Lawrence, Kansas, Program with Abstracts, p. 18.
Johnson, W., Fredlund, G., Wells, P., Stewart, J., and Dort, W., Jr., 1986, Late Wisconsinan biogeography of south-central Nebraska-the North Cove site (abs.): American Quaternary Association, 9th Meeting. Champaign, Illinois, Program with Abstracts, p. 89.
Kapp, R. O., 1965, Illinoian and Sangamon vegetation in southwestern Kansas and adjacent Oklahoma: Contributions from the Museum of Paleontology, University of Michigan, v. 19, p. 167-255.
Kapp, R. O., 1970, Pollen analysis of pre-Wisconsin sediments; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: Lawrence, Kansas, University of Kansas Press, p. 143-155.
King, J. E., 1973, Late Pleistocene palynology and biogeography of the western Missouri Ozarks: Ecological Monographs, v. 43, p. 539-565.
Kuechler, A. W., 1964, Potential natural vegetation of the conterminous United States: American Geographical Society, Special Publication 36, map.
Kurmann, M. H., 1985, An opal phytolith and palynomorph study of extant and fossil soils in Kansas (U.S.A.): Palaeogeography, Palaeoclimatology, Palaeoecology, v. 49, p. 217-235.
Lichti-Federovitch, S., and Ritchie, J. C., 1968, Recent pollen assemblages from the Western Interior of Canada: Review of Paleobotany and Palynology, v. 7, p. 297-344.
Markgraf, V., and Lennon, T., 1986, Paleoenvironmental history of the last 13,000 years of the eastern Powder River basin, Wyoming, and its implications for prehistoric cultural patterns: Plains Anthropologist, v. 31, no. 111, p. 1-12.
Martin, L. D., and Neuner, A. M., 1978, The end of the Pleistocene in North America: Transactions of the Nebraska Academy of Sciences, v. 6, p. 117-126.
McAndrews, J. H., 1966, Post-glacial history of prairie. savanna, and forest in northwestern Minnesota: Torrey Botanical Club, Memoir, v. 22, p. 1-72.
McAndrews, J. H., and Wright, H. E., Jr., 1969, Modern pollen rain across the Wyoming basins and the northern Great Plains (U.S.A.): Review of Paleobotany and Palynology, v. 9, p. 17-43.
Mehringer, P. J., King, J. E., and Lindsay, E. H., 1970, A record of Wisconsin-age vegetation and fauna from the Ozarks of western Missouri; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: Lawrence, Kansas, University of Kansas Press, p. 173-183.
Mott, R. J., 1969, Palynological studies in central Saskatchewan, contemporary pollen spectra from surface samples: Geological Survey of Canada, v. 69, p. 1-13.
Mundt, S., and Baker, R. G., 1979, A mid-Wisconsinan pollen diagram from Black Hawk County, Iowa: Proceedings of the Iowa Academy of Science, v. 86, p. 32-34.
Oldfield, F., and Schoenwetter, J., 1975, Discussion of the pollen evidence; in, Late Pleistocene Environments of the Southern High Plains: Fort Burgwin Research Center, Publication 9, p. 149-178. [available online]
Peterson, G. M., 1978, Pollen spectra from surface sediments of lakes and ponds in Kentucky, Illinois, and Missouri: The American Midland Naturalist, v. 100, p. 330-340.
Potter, L. D., 1967, Differential pollen accumulation in water-tank sediments and adjacent soils: Ecology, v. 48, p. 1,041-1,043.
Potter, L. D., and Rowley, J., 1960, Pollen rain and vegetation, San Augustin Plains, New Mexico: Botanical Gazette, v. 122, p. 1-25.
Raup, H. M., and Argus, G. W., 1982, The Lake Athabasca sand dunes of northern Saskatchewan and Alberta, Canada. I. The land and vegetation: National Museum of Natural Sciences, Publication in Botany, no. 12, 96 p.
Rogers, R. A., and Martin, L. D., 1984, The 12 Mile Creek site--a reinvestigation: American Antiquity. v. 49, no. 4, p. 757-764.
Rogers, R. A., and Martin, L. D., 1985, Early projectile points and Pleistocene fauna from sandpits near Wichita, Kansas: Transactions of the Kansas Academy of Science, v. 88, p. 46-50.
Ruhe, R. V., 1983, Depositional environment of late Wisconsin loess in the midcontinental United States; in, Late Quaternary Environments of the United States, v. 1, The Late Pleistocene, S. C. Porter, ed.: Minneapolis, Minnesota, University of Minnesota Press, p. 130-137.
Sangster, A. G., and Dale, H. M., 1964, Pollen-grain preservation of underrepresented species in fossil spectra: Canadian Journal of Botany. v. 42. p. 437-449.
Schmits, L. J., 1985, Prehistory of lower Perche-Hinkson drainage. central Missouri; in, Archaeological Investigation at the Columbia Regional Waste Water Facility, L. J. Schmits, ed.: KESA Cultural Resource Management Report, no. 16, prepared for U.S. Environmental Protection Agency.
Sears, P. B., 1961, A pollen profile from the grassland province: Science. v. 134, p. 2,038-2,039.
Shumard. C. B., 1974, Palynology of a lacustrine sink-hole facies and the geologic history of a (late Pleistocene?) basin in Clark County, southwestern Kansas: M.S. thesis, Wichita State University, Wichita, Kansas, 155 p.
Stewart, J. D., and Rogers, R. A., 1984, Analysis of pollen from the Trapshoot local-fauna quarry (Rancholabrean) of Kansas: The American Midland Natumlist, v. 112, no. l, p. 198-200.
Van Zant, K. L., 1979, Late-glacial and post-glacial pollen and plant macrofossils from Lake West Okoboji, northwestern Iowa: Quaternary Research, v. 12, p. 358-380.
VanZant, K. L., Hallberg, G. R., and Baker, R. G., 1980, A Farmdalian pollen diagram from east-central Iowa: Proceedings of the Iowa Academy of Sciences, v. 87, p. 52-55.
Voorhies, M. R., and Corner, R. G., 1985, Small mammals with boreal affinities in late Pleistocene (Rancholabrean) deposits of eastern and central Nebraska: Nebraska Academy of Sciences, Institute for Tertiary-Quaternary Studies, TER-QUA Symposium Series, v. 1, p. 125-142.
Watts, W. A., 1983, Vegetational history of the eastern United States 25,000 to 10,000 years ago; in, Late Quaternary Environments of the United States, v. 1, The Late Pleistocene, S. C. Porter, ed.: Minneapolis, Minnesota, University of Minnesota Press, p. 294-310.
Watts, W. A., and Bright, R. C., 1968, Pollen, seed, and mollusk analysis of a sediment core from Pickerel Lake, northeastern South Dakota: Geological Society of America, Bulletin, v. 79, p. 855-876.
Watts, W. A., and Wright, H. E., Jr., 1966, Late Wisconsin pollen and seed analysis from the Nebraska Sandhills: Ecology, v. 47, p. 202-210.
Webb, T., III, and McAndrews, J. H., 1976, Corresponding patterns of contemporary pollen and vegetation in central North America: Geological Society of America, Memoir, v. 145, p. 267-299.
Webb, T., III, Howe, S. E., Bradshaw. R. H. W., and Heide, K. M., 1981. Estimating plant abundances from pollen percentages--the use of regression analysis: Review of Paleobotany and Palynology, v. 34, p. 269-300.
Webb. T., III, Cushing, E. J., and Wright, H. E., Jr., 1983, Holocene changes in the vegetation of the Midwest; in, Late Quaternary Environments of the United States, v. 2, The Holocene, H. E. Wright, Jr., ed.: Minneapolis, Minnesota, University of Minnesota Press, p. 142-165.
Wells, P. V., 1965, Scarp woodlands, transported grassland soils, and concept of grassland climate in the Great Plains region: Science, v. 148, no. 1, p. 246-249.
Wells, P. V., 1970, Post-glacial vegetational history of the Great Plains: Science, v. 167, p. 1,574-1,582.
Wells, P. V., 1983, Late Quaternary vegetation of the Great Plains: Transactions of the Nebraska Academy of Sciences, v. XI (special issue), p. 83-89.
Wilson, L. R., 1966, Palynology of the Domebo site; in, Domebo. A Paleoindian Mammoth Kill in the Prairie Plains: Lawton, Oklahoma, Contribution of the Museum of the Great Plains, no. 1, p. 44-50.
Wright, H. E., Jr., 1970, Vegetational history of the central Plains; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: Lawrence. Kansas, University of Kansas Press, p. 157-172.
Wright, H. E., Jr., 1981, Vegetation east of the Rocky Mountains 18,000 years ago: Quaternary Research, v. 15, p. 113-125.
Wright, H. E., Jr., Almendinger, J. C., and Grueger, J., 1985, Pollen diagram from the Nebraska Sandhills and the age of the dunes: Quaternary Research, v. 24, p. 115-120.
Prev--Wisconsinan Mammalian Faunas || Next--Grass-opal phytoliths as climatic indicators
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version updated March 31, 2010. Original publication date 1987.
URL=http://www.kgs.ku.edu/Publications/Bulletins/GB5/Fredlund/index.html