Kansas Geological Survey, Bulletin 233, p. 345-358
by
Lee C. Gerhard
Kansas Geological Survey
Reefs are ideal for carbonate modeling because they have a modern geographic scale not dissimilar to their ancient counterparts. By assuming analogous accumulation rates and sea-level rate changes, it is possible to characterize reef growth mathematically. Generalizing reefs to develop comparisons has been difficult, but a process-based architectural classification solves much of this problem. The three classes of reefs--hydromechanical, biodetrital, and framework--are three members of a continuum of architectural styles. A detailed knowledge of reef fabrics permits interpretation of the influence of a variety of processes in response to environmental conditions. Sea-level change rates are one identifiable environmental condition that is now recognized as very important to reef architecture. The modern reefs of St. Croix are the basis for the classification scheme because of their detailed description and age dating. Current research on Pennsylvanian reefs is acquiring the data necessary to test ancient reefs; several reef complexes of other ages and geography have already been successfully tested. Equations for Holocene reef growth and sea-level rise rates can now be used to describe ancient reef architectural development.
An Acrobat PDF file containing the complete paper is available (1.1 MB).
Studies of modern coralgal reefs provide insight into the processes that control the development and architecture of ancient reefs. Although modern carbonate analogues generally suffer from scale differences with their ancient counterparts, for reefs the depositional scales are not significantly different. Modern reefs have characteristic architectures (framework, biodetrital, hydromechanical) that reflect their settings and the balance of processes that create them. Core drilling provides extensive information about the vertical sequence of fabrics in modern reefs.
Geology has not yet matured into the numerically based science necessary for modern computer analysis and prediction of physical characteristics. The advent of computer modeling of sedimentary systems has begun to provide predictability of facies on a gross scale, although most appear to replicate existing geologic systems rather than predict the behavior of specific time-space-process systems [e.g., Franseen and Watney (1989)]. Understanding reservoir compartmentalization and the ability to characterize petroleum reservoirs has become significant as the United States attempts to recover additional petroleum from existing pools. Numerical analytical techniques and computer modeling are methods that may assist in accessing that additional petroleum.
It is important that geology demonstrate reproducibility of experimental results and predictability for numerical values to be useful. Consequently, qualitative geologic interpretations based solely on logical deductions from observations are no longer sufficient. Integration of observations into computer models that can accurately predict system behavior has been a major step forward in the study of detrital sediment systems.
Many models of detrital systems appear to be valid and are useful for predicting facies; however, few have attempted to model carbonate systems [see, however, Koerschner and Read (1989) and Lawrence et al. (1990)]. Comments about carbonate modeling are reminiscent of Folk's (1959) statement that many students consider carbonates too difficult to study. Folk's carbonate classification, although not widely used today, provided an elegant view of carbonates that solved that difficulty. We face that same complexity in modeling carbonates today, but a process approach to understanding carbonates can lead to successful modeling. Gerhard and Burke (1986, 1990) and Burke and Gerhard (1987) proposed a reef classification based on processes that is designed to provide a view of the evolutionary architecture of reef systems; this classification is discussed and applied herein. The classification provides a basis for interpreting sea-level change rates and mechanical energy and slope settings for reefs. Like Folk's carbonate classification, the classification need not be generally adopted for the concept to lead to better understanding of the carbonate processes; subsequent classifications may be better.
One of the major problems in carbonate modeling is the inability of geologists to model geologic time directly and to measure carbonate depositional parameters directly in settings analogous to those commonly found in the rock record. Holocene sea levels are high, but the continental shelves and interiors are not flooded as they were in Pennsylvanian time, for instance, and certainly there are no good analogues for earlier broad epicontinental seas in carbonate-producing latitudes. These characteristics make carbonate modeling difficult. Reefs, on the other hand, are one part of the carbonate world that are similar in scale to their ancient counterparts, largely because reefs depend on areally restricted oceanographic settings.
Another problem is that few geologists appreciate the elegance and simplicity of carbonate sediment generation and accumulation processes. Carbonates are intrabasinal in origin and are of simple mineralogy (calcite, aragonite, magnesium calcite, and dolomite). Extensive accumulations are limited to warm latitudes. Carbonates contain environmentally diagnostic organism assemblages as part of most sequences and are sensitive to the depositional environment. That is, small changes in environmental setting can cause significant changes in sediment and organism characteristics. These characteristics make carbonates ideal model components.
Modern reefs, coral, and coralgal accumulations occupy a unique niche in the carbonate world. They are identical in scale with their ancient counterparts. Other types of reefs have analogues as well, from wave- and current-generated organic bar deposits to phylloid-like accumulations of Halimeda and Halimeda-dominated communities. Deriving ancient counterparts of modern reefs is made difficult by the highly generalized fabric descriptions of most carbonate rock bodies. Even descriptions written by carbonate experts are often too general to be useful for modeling. For instance, intervals of a few millimeters or centimeters thickness may be important for interpretation of the presence of a "condensed section" and consequent rapid sea-level rise, but such intervals may not be discernible at outcrops. Because of this limitation, much reef modeling will be based on existing oil-well cores, cores from scientific drilling programs, and new outcrop descriptions. These can be readily compared to drill records of modern reefs.
Several limiting factors control characteristics of specific fabrics within carbonate sediment bodies. Fabrics reflect mechanical energy levels by sorting and rounding, although these parameters may not be directly compared with sediments of siliciclastic origin (Gerhard and Stolzman, 1974). Turbidity is a controlling factor on organic productivity, relating to photosynthesis and sediment cleaning but mostly to availability of light [e.g., Gerhard (1978)]. Light level is more controlled by turbidity than by absolute water depth, although selective absorption of the red spectrum is water depth related.
The modeling progress discussed here depends heavily on my detailed core studies of modern reefs and on studies by Adey and Burke (1976) and Burke et al. (1989) in the Caribbean. The ancient reef discussion is based on considerable proprietary work, new scientific core drilling, and field observations from many localities.
There is no assurance that modern sea-level change rates and absolute sea-level values are similar to those of the past, although the assumption is that they are comparable. Organism plasticity is assumed to be high throughout geologic time and thus does not limit models. Finally, the proposed modeling assumes that reef-associated carbonate processes have not changed through time. The causes of sea-level changes are considered trivial concerns in this study.
All three-dimensionally discrete bodies of organically derived carbonate rocks are considered to be reefs. They result from the interplay of definable major sedimentary processes, and their depositional and diagenetic settings can be interpreted from observing their vertical sequence of fabrics [see also Longman (1981)]. Reef fabrics and architecture evolve in response to oceanographic setting, changing rates of sea-level rise, and processes of carbonate generation and degradation. Availability of organisms within a specific reef setting and interruptions in evolution may determine final architecture or compartmentalize architectural styles.
Part of the thesis is that the rates at which the processes of reef generation operate determine the fabric of any reef formed. Rates of organism growth, degradation, and detrital accumulation determine the relationship between water depth and sea-level rates of rise, for instance. As a corollary, fabrics of sediment (rocks) generated at a reef site can be interpreted with respect to the rates and balances of processes that operated, as discussed later [see also Gerhard and Burke (1990)]. The overall architecture (building) of a reef is thus derived from a sequence of fabrics (building blocks) that developed in response to various processes in specific oceanographic settings.
From these constraints and the theoretical premise, Gerhard and Burke (1990) deciphered a continuum of architectural styles that are controlled more by sedimentary processes than by biotic evolution or community succession. Fabrics result from the interplay of organic growth, dynamic processes, and depositional topography.
I deliberately use the term "reef' as a general term with appropriate architectural modifiers rather than "bank" or "bioherm" to stress the interpreted continuum of fabrics and processes. One member of the continuum is "framework" reefs (Lowenstam, 1950; Fagerstrom, 1987) , so well illustrated by many of my colleagues; another member is wave-initiated piles of skeletal carbonate sand (Marshall and Davies, 1988). The third member is reefs composed of mixed in situ benthic organisms and internally derived detritus. Biotic generation, degradation, and mechanical transportation interweave to provide reef fabrics, which make up the architectural style. The rates at which these processes operate seem to be controlled by rates of sea-level change, especially rates of rise and organic productivity (Adey and Burke, 1976). The mix of primary mineralogies coupled to the fabrics characteristic of each reef class influences ultimate porosity of a reef through early diagenesis (Milliman, 1974).
Carbonate sediment is a product of growth of carbonate skeletons, modified by biologic and mechanical degradation, resulting in sediment of varying size and shape. This sediment is either incorporated into the reef or transported out of it. The balance among skeletal growth, degradation, and transportation determines the reef body fabric and architecture and ultimately the type and degree of diagenesis. The degree of transportation of detritus out of a reef system is a function of wave energy and depositional slope. Transportation of detritus from a reef system creates the opportunity for a framework to develop and for syngenetic cementation. Lack of sediment removal provides smaller pore space and inhibits skeletal growth and syngenetic cementation. Production of carbonate sediment within an active reef system is so rapid that frameworks cannot develop unless the sediment is removed; frameworks can literally drown in their own detritus. At the opposite extreme, a hydromechanical pile of loose sediment will result from transportation or sweeping of carbonate sediment into a wave-induced bar form.
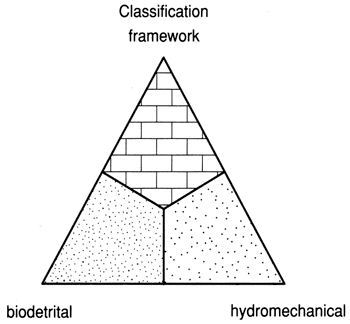
The effects of the three major reef-construction processes--skeletal generation, degradation, and transportation--can be graphically represented on a ternary diagram (fig. 1). A corresponding reef classification terminology--framework, biodetrital, and hydromechanical--is derived from the processes (fig. 2).
Figure 1--Ternary diagram showing the three major carbonate processes that determine reef architecture. Connecting plotted points, which represent individual samples, traces the evolution of process influence on the reef.
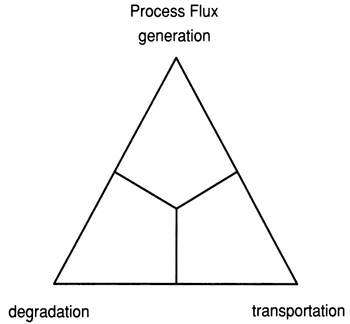
Figure 2--Ternary diagram of three major reef classes based on architecture developed by processes shown in fig. 1. Points on the two ternary diagrams correspond; that is, framework reefs are dominated by generation processes, biodetrital reefs by degradation, and hydromechanical reefs by transportation.
Eustasy is the remaining dynamic process that must be considered as controlling reef fabric. Neumann and Macintyre (1985) have considered this relationship in their "catch-up, keep-up, or give-up" view of succession. However, although they addressed organism response to sea-level changes, they did not consider the sequence of fabrics that marks a normal cycle. Normal cycles, characterized as sine curves, periodically have rapidly changing rates of rise, which, in conjunction with relatively stable organism growth rates, provide large variations in absolute water depth.
Framework reefs have organic frameworks that are further colonized by other organisms. They frequently contain a low amount of internal sediment and a high amount of syngenetic cement (James, 1983). They usually grow on a steep depositional slope flushed by strong impinging waves and currents. Degradation of carbonate skeletons of framework and other organisms supplies detritus to the reef system, but much is swept out of the reef and redeposited in aprons downslope and to the rear of the reef. Space that would otherwise be occupied by detrital skeletal sediment is left open but is later filled with cement. Porosity after early diagenesis may be low because of internal cementation.
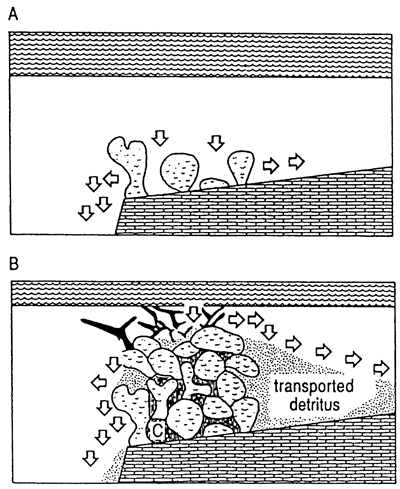
One modern example of a framework reef is Galeta Point, Panama. Macintyre and Glynn (1976) drilled on a transect across the fringing reefs at Galeta Point, where a Holocene reef complex has developed on a Miocene substrate adjacent to a steep slope that descends from sea level to at least 14 m (46 ft) within 100 m (300 ft) of the reef crest. The complex is under the direct influence of strong trade winds. In this exposed, high mechanical energy setting, corals form the bulk of the reef and syngenetic cementation is extensive, occurring most abundantly in the seaward portions of the reef. Characteristics of this reef setting and resultant reef architecture are steep depositional slope, high mechanical energy, framework, and syngenetic cementation (fig. 3). Around the island of St. Croix, US Virgin Islands, in a few exposed, high depositional slope settings, reef formation is similar to that described in Panama by Macintyre and Glynn (1976) in that framework development predominates and syngenetic cements exist.
Figure 3--Typical setting and fabric of framework reef. (A) Reef organisms grow on shelf-edge setting; sediment generated in reef is flushed. (B) Flushing permits cements to precipitate in lower-energy interstices; shelf-edge settings enhance flushing and propensity for framework generation. Black is most recent shallow, rapid-growth organism, such as Acropora palmata.
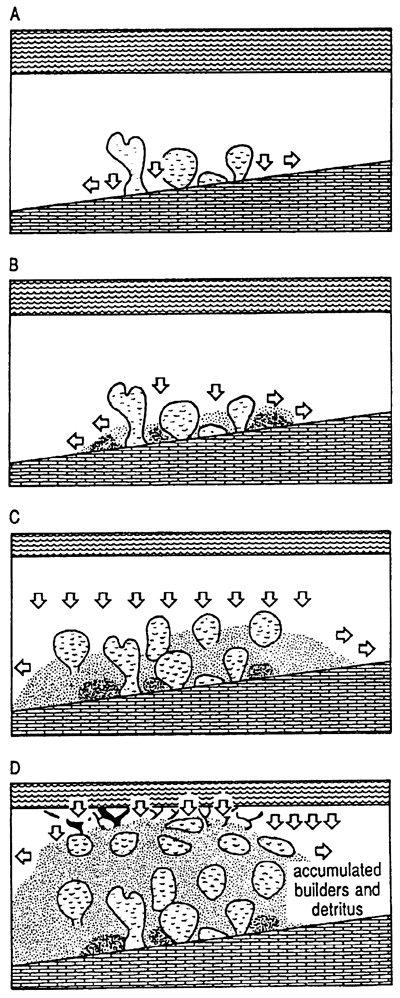
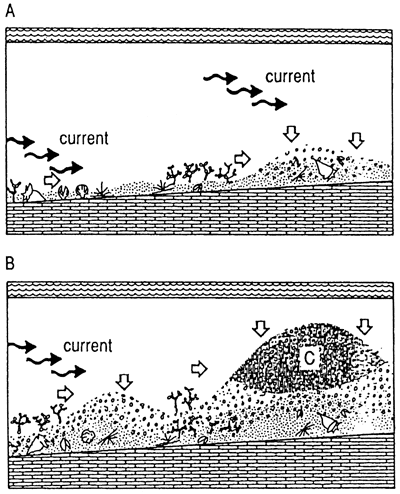
Biodetrital reefs In other settings with differing balances of processes, other characteristics develop. Biodetrital reefs grow on less steep depositional slopes with less strong wave trains and currents than framework reefs. Sediment produced by skeletal degradation largely remains unswept in the reef, smothering settlement of additional reef frame organisms. Biodetrital sediment is produced within the reef mainly by bioerosion of living and dead skeletal material (Zankl and Shroeder, 1972) (fig. 4) but also by mechanical trituration. The resultant architecture is over 50% sediment and less than 50% in situ skeletons. Individual fabrics may be completely sand or may consist of mixed sand and skeletons, depending on where the sample is taken. Sediment fills large pores so that internal cementation is minimal. Typically, in modern reefs the total sediment and skeleton package is of mixed mineralogy; that is, magnesium calcite, aragonite, and calcite grains are thoroughly mixed in the reef so that early diagenesis, particularly meteoric diagenesis, selectively dissolves aragonite and/or magnesium calcite, creating a sequence of porosity development and occlusion. Porosity in fossil reefs tends to be relatively high because of the original mixed mineralogy. Biodetrital reefs appear to be the most abundant class in the geologic record.
Figure 4--Typical setting and fabric of biodetrital reefs. Sediment is generated in the reef system but is only partly transported out of it. Consequently, coral skeletons appear to float in debris of sand and rubble of other size; reefs may drown in their own debris. Interstices are filled with sediment, substantially decreasing the probability of syngenetic cementation and growth. Mixed mineralogy and grain sizes of these systems enhance secondary porosity development. (A) Organisms grow in setting that is not effectively flushed. (B) Sediment begins to accumulate within reef. (C) Reef continues to grow, but significant sediment inhibits some growth and isolates individuals. (D) In the culmination phase of the formation of a biodetrital reef, the sediment has greater volume than the reef organisms. Black is most recent shallow, rapid-growth organism, such as Acroporapalmata, which may form small portion of framework at leading edge of reef.
Extensive coring of reefs through the eastern Caribbean, for example, by Adey (1975), Adey and Burke (1976, 1977), Adey et al. (1977), Macintyre et al. (1985), and Hubbard et al. (1985), provides a view of the architecture of modern coralgal reefs that permits further inferences to be made about the relationships among fabrics, processes, and settings, particularly for biodetrital reefs.
In most St. Croix reefs, where depositional slope becomes gentler with consequent lessened wave energy, a biodetrital architecture is developed. In this setting reefs are composed predominantly of sediment; commonly, 60-70% of the mass is biodetrital sand or finer material. Only a thin veneer of living coral appears to form the framework. Wave and current energies are not high enough to sweep the biodetritus from the reef, nor is the depositional slope steep enough to cause extensive downslope transport of sediment. In effect, the reef framework organisms drown in their own detritus; growth and reproduction to form an interlocking framework is inhibited by the large amount of detritus deposited within the reef. Individual organisms are widely spaced and thus prevented from interlocking.
Cores taken at the western end of Teague Bay reef (Sand Cay) on St. Croix (Gerhard, 1981) further demonstrate this relationship, as do the cores from Martinique taken by Adey and Burke (1976, figs. 12 and 13). The Sand Cay cores show biodetritus deposited on pre-Holocene limestone or Holocene fine-grained siliciclastic sediments; only a thin veneer of living reef coral or large rubble is present (fig. 5). No cementation has been observed in this setting, and pores are abundant but small compared to the framework reefs. The polytaxonomic origin of the reef pile indicates that the sediment is mostly aragonite and magnesium calcite with fewer grains of calcite.
Figure 5--Lithologic (sedimentologic) logs of holes drilled at Sand Cay, St. Croix, showing that the reef first accumulated sand, then rubble and sand, and, finally, a living capstone of framework reef. This took place during a sea-level rise that has slowed over the last 7,000 years.
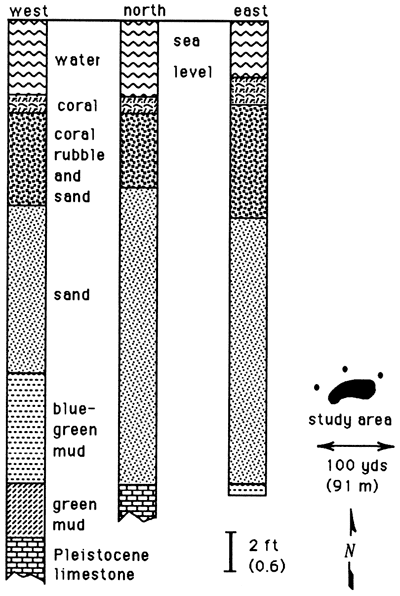
Burke et al. (1989) have shown that the length of Teague Bay reef is similar to the architecture seen at Sand Cay. Organic carbonate sands form the initial reef, followed upward by a mixed assemblage of head corals embedded in sand of reef origin and capped by a framework of branching corals and coralline algae.
Characteristics of this reef setting and resultant reef architecture are moderate depositional slope and moderate mechanical energy. The resulting main reef is a biodetrital sediment mass that has little or no framework and virtually no cementation (fig. 6). The reef lies on a foundation of hydromechanically generated sandbar sediment and is capped with a thin veneer of framework.
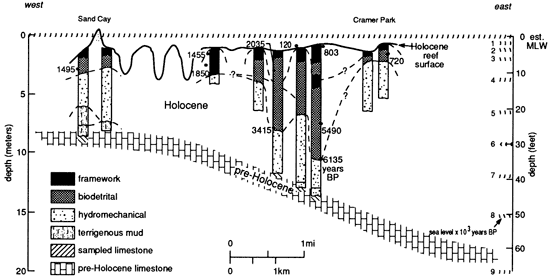
Figure 6--Cross section along reef strike, Teague Bay reef, St. Croix, showing relative abundance of framework organisms and biodetrital sediment. Modified from Burke et al. (1989).
Hydromechanical reefs are characterized by standing or oscillatory wave sets, low depositional slope, mechanical trituration of skeletons, and, possibly, longshore currents. In these settings accumulation of organically generated carbonate sediment predominates by hydromechanical piling, perhaps later colonized by benthic skeletal animals and plants. Although many students of carbonate sedimentology would argue that such wave- and current-generated three-dimensionally discrete bodies of organically generated but hydromechanically reworked sediment are not reefs, I view these as simply one member of a three-member continuous architectural series.
Davies and Marshall (1985), among others (Hine et al., 1988; Marshall and Davies, 1988; Orme and Salama, 1988; Phipps and Roberts, 1988; Roberts et al., 1988), documented hydromechanically formed piles of Halimeda (bioherms, in their terminology) behind the main coral reef fronts of the Great Barrier Reef, Australia, and in other localities in the Pacific and Caribbean. They showed that the algal plate mounds within these reef complexes are significantly older than the framework coral reefs themselves. They attribute initiation of algal accumulation to upwelling onto the preexisting Pleistocene limestone platform, which dips gently westward from the front of the coral reefs. However, their seismic profile geometry and the amplitude of the banks strongly suggest hydromechanical accumulation in addition to in situ growth; the reefs appear to have some internal bedding and external bar-type geometry. Little seismic evidence exists for current-bedded internal structure, but bioturbation tends to be high in shallow-water Halimeda accumulations in St. Croix (Gerhard, 1978). High productivity of Halimeda can be attributed to upwelling, but such settings also provide the necessary mechanical energy to accumulate low-density algal plates. Algal and other skeletal grains of the present surface appear to be mostly autochthonous to the accumulations. Additional growth occurs preferentially on the tops of the wave-generated mounds in response to higher ambient light conditions (less turbidity), which enhance photosynthesis [see also the discussion by Gerhard (1978)]. No doubt the reefs have been colon zed by Halimeda throughout their growth, so that in situ algal plates are combined with mechanically added sediment grains to form the present geometry. At present, wave trains are cut off by the coral reef front, and only low wave energy impinges on these reefs.
Original descriptions suggest that these Pacific Ocean algal sediment bodies were entirely modern and formed in the deep water that now covers them; the seismic profiles indicate otherwise. Also, photosynthesis is severely inhibited at the modern depths; in the Caribbean modern flooding surfaces are inhabited by growing mollusk-Halimeda banks [see Gerhard (1978)]. Prolific growth of the resident algae would seem to be precluded, but full coring of the banks should answer the questions raised about the banks' origin. The algal bodies are similar in overall taxonomy and morphology to Carboniferous phylloid-algal reefs, particularly of the midcontinent, Permian basin, and Paradox basin, United States.
Characteristics of the hydromechanical reef setting and resultant reef architecture are low depositional slope, variable but frequently low mechanical energy, and hydromechanical accumulation of reef grains. In polytaxon hydromechanical reefs (multiple-organism carbonate sand banks in the modern foraminiferal- skeletal reefs in the Pennsylvanian, for instance), there appears to be little or no syngenetic cementation (fig. 7). However, in phylloid-algal accumulations, isopachous cements predate dissolution of the aragonitic algal plates and enclose internal silt and pellets. When algae or other sediment producers are prolific, reef grains will be trapped in situ.
Figure 7--Hydromechanical reef showing the accumulation of organic debris in a wave-dominated regime. Establishment of a productive community either on or nearby the reef with sufficient constancy of wave and current geometry and intensity can provide necessary skeletal materials. Although an extreme example of this reef type could be a carbonate-sand bar, most are propagated by vegetative communities.
Hydromechanical reefs do not usually provide a suitable substrate for the settlement of framework organisms because the grains are subject to both bioturbation and wave and current reworking. Deepening of water may enhance the chances for settlement of frame builders. Ooid banks are not included in this class of reefs because they are not of demonstrable organic origin.
As reefs of any framework percentage grow, detritus is generated by biologic and hydromechanical degradation and accumulates within the reef. If this detritus is not removed from the reef, it will inhibit organic skeletal growth. If detrital generation is high and transportation of detritus out of the reef is low, the accumulation of detritus (bioclasts) will provide a significant or major part of the reef mass compared to the amount of interlocking framework, establishing a biodetrital reef. In the extreme, the reef will develop as a three-dimensionally discrete body of organic detritus, a hydromechanical reef.
In contrast, if the reef setting is such that waves and currents winnow the degradation products (biodetritus) from the reef, then the reef will be characterized by organic skeletons and open space, encouraging the establishment of a well-developed framework. Thus fast-growing reef organisms in a sediment-swept setting will most likely produce framework reefs.
The rates at which these processes operate are controlled by various chemical, thermal, and mechanical attributes of the sedimentary environment and are not discussed here. Instead, I focus on two additional attributes that are not commonly discussed: substrate geometry and rate of sea-level change.
Substrate geometry can be a major factor determining reef fabric because the depositional slope and topography modulate the wave and current regime of the ocean. Shelf-edge reef locations with steep declivities promote growth of framework-generating organisms and concurrent removal of detritus from the reefs. Uninhibited wave trains impinge on shelf-edge settings, transferring potential energy directly to the reef mass rather than gradually transferring energy to the bottom and to reefs on the lesser slopes of platforms.
I conclude that reef fabrics are gradational. The major architectural styles and the reef classification should recognize a continuum of dimensions. Three major reef types can serve as members of this classification system (fig. 2). These members are directly comparable to the process ternary end-members that are the basis of their diagnosis (fig. 1), and together they permit an interpretation of the physical and biologic setting of a reef preserved through its fabrics and architectural style. The three reef class names (framework, biodetrital, and hydromechanical) are descriptive, evolve from sedimentary and oceanographic processes, and are continuous variables. For these reasons the nomenclature ternary diagram is divided into fields that permit the grouping of similar but not identical reefs. Others may wish to substitute "reef" for "framework bioherm" for "biodetrital," and "bank" for "hydromechanical," but the terms used here are more descriptive, better related to the objects described, and more useful in the diagnosis of ancient reef evolution.
Most classifications and terminology sets fail to recognize the growth sequence of modern reefs and their ancient counterparts, frequently carrying them from the hydromechanical class through the biodetrital class and sometimes to the framework class. Current reef nomenclature assumes stable environmental conditions during reef growth and thus homogeneous reef fabrics (i.e., reef, bank, bioherm). Not all reefs go though the complete metamorphosis of fabrics, but a sufficient number do, developing an idealized reef architecture and nomenclature sequence.
The Teague Bay reef system, studied by Adey and Burke (1976), Burke et al. (1989), and Gerhard (1981), is located on the landward margin of the St. Croix platform. No shelf declivity is present, although there is a deeper reef along the shelf-edge margin that appears to be relatively free of detritus. This modern reef complex contains all three architectural styles. The initial part of the reef is pure sand (hydromechanical), followed upward by a thick sequence of corals (or other frame builders) embedded in sand (biodetrital) and then covered with a veneer of branching corals (framework), much of which is still living.
Adey and Burke (1976) and Burke et al. (1989) have provided the most detailed data sets for modern reefs that characterizes this idealized sequence. From their reef coring program in the Caribbean, they were able to determine ages for the sampled corals and the corals' position within the reef sequence. These data were then compared to sea-level history for the region to provide a time-depth data series (fig. 8). Cores drilled on Sand Cay, St. Croix (Gerhard, 1981), add details of reef architecture to the data of Adey and Burke and illustrate the sequence of fabrics typical of these coralgal reefs (fig. 5). Reef fabrics progressed through the sequence of nomenclature in response to changing environmental conditions, mainly water depth and turbidity, which decrease photosynthetic activity. Water depth changes result from changes in the rate of sea-level rise relative to reef accretion rates.
Figure 8--Time-depth sea-level curve for St. Croix [based on Adey and Burke (1976)].
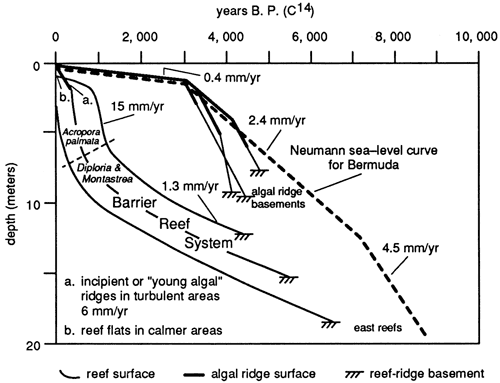
The Holocene sea-level rise postulated by Adey and Burke (1976) is asymmetric; rapid sea-level rise is followed by ever-slowing rates of rise. Absolute water depth is not congruent with the rate of sea-level rise; rather, maximum water depth occurs near the inflection point from the greatest rate of rise to a moderate rate of rise, reflecting rates of sedimentation and reef organism growth (fig. 9).
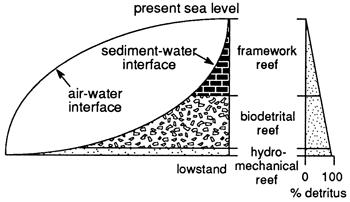
Figure 9--Relationships of sea-level change to absolute water depth, detritus generation, sedimentation, and reef architecture. Left-hand portion shows reef classes plotted against sea level (air-water interface) and the sediment-water interface. At lowstand a hydromechanical reef initiates reef development in shallow water. Absolute water depth increases as rapid sea-level rise takes place, and biodetrital reef growth is initiated. When the biodetrital reef growth rate exceeds the rate of sea-level rise, the absolute water depth decreases. Finally, when the reef grows sufficiently shallow to permit rapid-growing reef framework organisms to thrive, a framework reef architecture is developed and reaches the surface. The abundance of detritus in the reef decreases upward with change in reef architectural class (right-hand side).
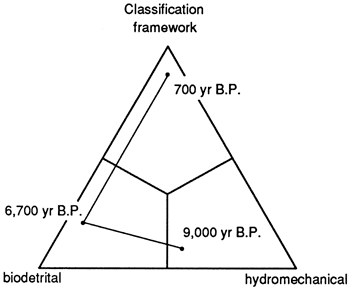
Characterization of the Teague Bay reef requires specification of time, water depth, and sediment level in vertical section. One easy method of analyzing the evolution of the reef (and its resultant nomenclature) is to plot reef fabric through time on a ternary diagram (fig. 10). It appears that framework reefs are a special result of lowered rates of sea-level rise (apparently at the conclusion of a rise); subaerial erosion coincident with relative lowering of sea level may destroy many of the framework veneers, preserving only their earlier biodetrital or hydromechanical fabrics.
Figure 10--Ternary diagram of reef classification based on reef architecture with plot of samples and consequent interpretation of evolution. Based on data by Adey and Burke (1976) from St. Croix. Reef was initiated as a hydromechanical reef with stabilizing head corals at 9,000 years B.P., then gradually evolved into a full biodetrital reef with minor framework corals at 6,700 years B.P., and finally established a mechanical framework at 700 years B.P.
Initial Teague Bay reef growth was as a sand-dominated bank with a relatively small number of corals, coincident with the initial sea-level rise over the exposed Pleistocene substrate. After initial colonization, water depth rapidly increased during the highest rate of sea-level rise (4.5 mm/yr), from approximately 8,000 years B.P. to 7,000 years B.P. Initial hydromechanical accumulations were supplanted by coral-head-dominated biodetrital reefs, but water depth continued to increase because the sea-level rise from 7,000 to 3,200 years B.P. was greater than the sediment accumulation rate (1.3 mm/yr). Slopes were too gentle and water depth too great for waves of normal amplitude to winnow sediment from the reef. At 3,200 years B.P. the rate of change of sea-level rise dropped to 0.4 mm/yr, whereas the sediment accumulation rate was maintained at 1.3 mm/yr. Significant accommodation potential existed for continued growth. Head corals and detritus characterized reef accretion when sedimentation and organic reef growth exceeded the lowered rate of sea-level rise. Thus absolute water depth decreased.
At 1,200 years B.P. water depths were shallow enough that Acropora palmata could establish on the reef. This coral is productive only in shallow water and has a growth rate in excess of a centimeter per year today on the same reef. Prolific A. palmata growth created a veneer of framework on the upper surface of the Teague Bay reef, which breaks surface today, although the lower slopes of the reef remain a biodetrital reef. Much of the detritus generated within the A. palmata zone is washed landward into the backreef or down the foreslope, permitting a thin upper surface framework to grow.
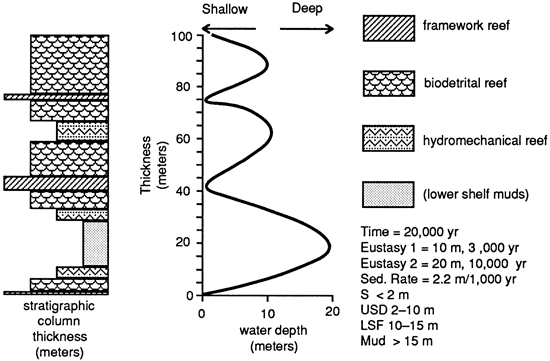
Initial models of time versus depth versus architectural style were made using the one-dimensional model developed by T. A. Cross and M. Lessenger (Colorado School of Mines; unpublished Macintosh program). The model was designed for detrital systems analysis (fig. 11) and its parameters include definition of eustatic cycle duration (in this case, two different duration and amplitude cycles) and a sedimentation rate averaged from the St. Croix data. Water depths for the various reef architectural classes were set on nominal values, again based on ranges observed on St. Croix. Figure 11 illustrates a run of 100,000 years. This run resulted in a shoaling-upward reef in which the framework grew at low water levels and increasing amounts of biodetrital reef accumulated through time.
Figure 11--Reef architectural sequence generated by the Cross-Lessenger model. Modeling parameters include definition of two eustatic cycles. The sedimentation rate is averaged from the St. Croix data. Model was run for 100,000 years.
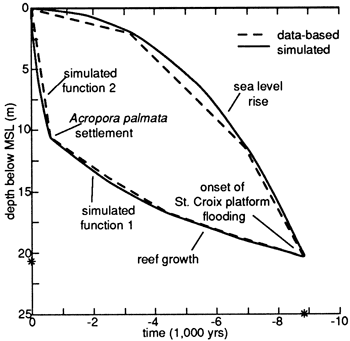
Work with this particular model was valuable for predicting a generalized sequence of architectural styles. However, that version of the program has not been able to model the St. Croix data fully. To incorporate both the varying accumulation rates of the reef surface and the varying rates of sea-level rise, I derived polynomial equations for both. For sea-level rise an unlimited equation of relatively simple construction appears to be adequate:
Ysea = A0 + A1X + A2X2 + A3X3 + . . . , (3)
where A0 = 2.77 x 10-16, which is approximately equal to 0, A1 = 0.0156337, A2 = 0.142036, A3 = 0.0 114601, Ysea is the elevation of sea level, and X is the time of that elevation (elevation is calculated from present). For the reef accumulation rate, two limited polynomial equations provide that
Ysed = A0 + A1X + A2X2 + A3X3 + . . . , (3)
where A0 = 8.8547, A1 = 2.48163, A2 = -0.219042, and A3 = 0.00897463 for values of 10.5 ≤ Y ≤ 20; X ≤ 0.7; Ysed is the elevation of the sediment-water interface, and X is the time of that elevation (elevation is calculated from present). However, when Y ≤ 10.5 and X ≥ 0.7, A0 = 0.487708, A1 = 23.4657, A2 = -13.0544, and A3 is negligible.
The calculated curves closely fit the measured St. Croix curves (fig. 12). These equations are being incorporated into a new version of the Cross-Lessenger model and into a two-dimensional model of carbonate and clastic sedimentation developed for study of the stratigraphic architecture and facies distribution within Pennsylvanian cyclic sequences of the midcontinent region (John French, personal communication, 1990). There remains the problem of characterizing longer-period sea-level curves, testing these simulations against published interpretive curves for pre-Holocene and pre-Quaternary time, and comparing results to detailed measured stratigraphic sections.
Figure 12--Plot of calculated curves versus measured St. Croix curves [derived from Adey and Burke (1976)].
The existing architectural classification and consequent models do not yet address the heterogeneity of reefs in the plane of depositional dip. This is not a trivial question, for reservoir heterogeneity studies must address three-dimensional space. French's developing model (personal communication, 1990) portrays dip facies changes, but in a stratigraphic sense, not at the individual reservoir scale.
As previously discussed, few existing measured sections or core descriptions can fully analyze overall reef architecture or decipher sea-level history. A core was drilled in the Ismay zone of the Hermosa Formation, San Juan River canyon, Utah, in June 1989 by the Kansas Geological Survey as part of its research on Pennsylvanian reefs and sequence stratigraphic concepts. The core recovered a nearly complete sea-level cycle of deposition (fig. 13), providing a good comparison of lithology and sea-level change rates. The Ismay reef is a hydromechanical reef, composed mostly of phylloid-algae organic particles.
Figure 13--Stratigraphic section cut by the KGS core 89/1 in the San Juan River canyon. Ismay zone of Hermosa Formation, Pennsylvanian.
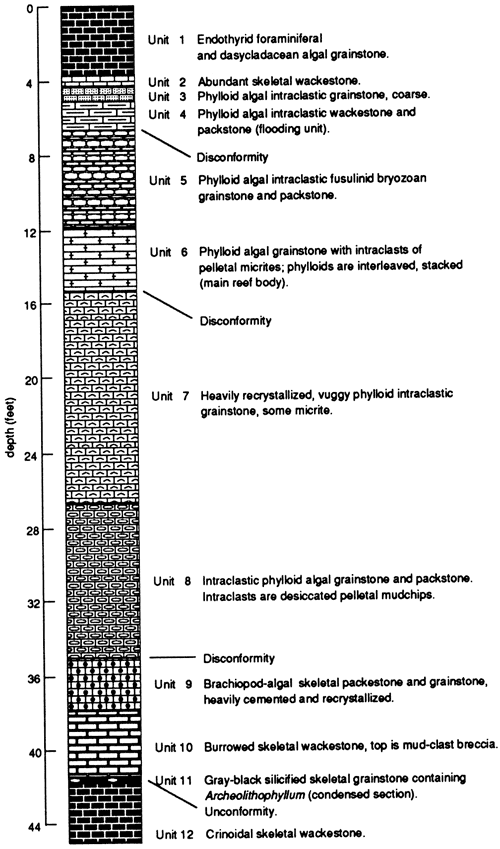
Early descriptions from field exposures suggest that the reefs are exclusively algal. However, there is a well-defined vertical zonation of codiacean algal plates, coupled with mud clasts and other skeletal grains (figs. 13 and 14). Reservoir properties were formed by multiple diagenetic events, in- cluding post-cementation but early dissolution of aragonite algal fronds.
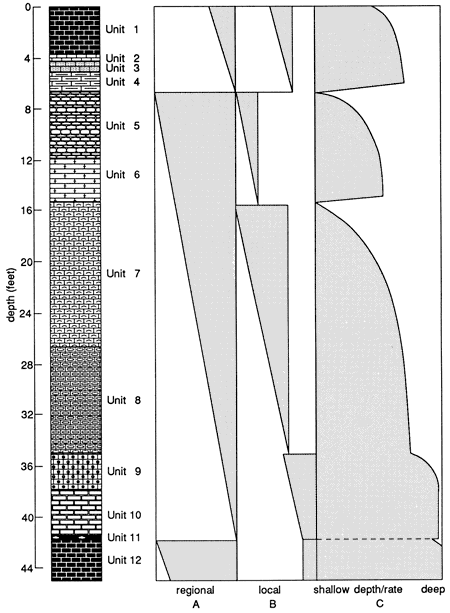
Figure 14--Stacking geometries and sea-level curve for the Ismay core from the San Juan River canyon. Column A represents the interpreted major stacking geometry for the region. Column B is the interpreted local stacking geometry, which could also include changes in reef geometry and sea-level changes. Column C is the interpreted sea-level changes that resulted in the stacking geometry. Steepness of curve represents rate of change.
Reef growth was initiated with a sea-level rise. The lowest beds cored were organic crinoidal muds, typical basinal deposits for the Pennsylvanian. A thin [0.15 ft (3 cm)] condensed section consists of silicified black skeletal micrite. Archeolithophyllum is abundant in this unit, the only unit in which it has been observed, and no other alga is present. This condensed section represents a relatively long span of time for its thickness; the rapidly rising sea shut off sediment generation even in the carbonate environment, and the absolute water depth increased because of the high rate of sea-level rise and the low accumulation rate. The preponderance of Archeolithophyllum, a fossil precursor rhodophyte, in that unit probably indicates a deeper and more turbid depositional system.
Shoaling-upward is indicated by the advent of bioturbated skeletal wackestones and then grainstones above the condensed unit. Mud clasts at the top of unit 9 (fig. 13) suggest that a minor cessation in rise occurred. Preliminary diagenetic interpretation suggests that an unconformity separates units 8 and 9.
The overlying rocks (fig. 13, units 7 and 8) are mixed phylloid-algal plates that are heavily recrystallized, except where they occur in micrite matrix, and interspersed with abundant micritic intraclasts, suggesting that high mechanical energy and exposure conditions alternated during deposition. Tubular foraminifers and other skeletal grains are present. Shallow-water deposition is indicated. Full phylloid reef development occurs in unit 6, where phylloid-algal plates and mud clasts predominate and other constituents are restricted to thin interbeds.
A more muddy fabric occurs in unit 5 (fig. 13) with the advent of a mixed skeletal assemblage typical of modern grass beds. Thin-walled gastropods, foraminifers (including fusulinids), and bryozoans are present within a predominantly phylloid algae and intraclast fabric. This unit appears to be a small cycle in itself. These fabrics culminate in unit 3, which has a flat pebble conglomerate bed, indicating final exposure of the reef body. Mechanical energy was high, based on the sorting of the phylloids and their interleaving.
The remainder of the core is deeper, quieter water carbonate with dasycladacean algae, endothyrid foraminifers, fusulinids, and tubular foraminifers. This represents the start of the next cycle, with low rates of sea-level change.
Porosity within this system is developed mainly in the heavily wave- and current-worked phylloid and intraclast-dominated fabrics. Much of the porosity is related to early submarine cementation and then apparent meteoric dissolution of the aragonitic algae and mud, but full paragenesis is complex. The tops of the mounds are rounded and aligned as if by prevailing currents, although it is unclear whether the reefs are spaced by oscillatory waves.
In the Ismay algal reef hydromechanical accumulation of intraclasts and phylloid plates accumulated to form reefs. Early cementation in the subaqueous setting provided some rigidity to the mass; increased shallowness of the reef tops increased the growth rates of the algae because of increased light for photosynthesis, further differentiating the reef topography. In this case no settlement of potential frame-building organisms occurred; thus the reef never changed from the initial hydromechanical accumulation.
In contrast, field examination of reefs in the Barker Creek zone of the Hermosa Formation, below the Ismay, encountered a nearly 6-ft-thick (1.8-m-thick) basal wave-washed and current-bedded initial deposit overlain by a biodetrital accumulation of Chaetetes and skeletal sand, which in turn is overlain by a massive "mud" reef core. Flank deposits indicate that a mixed fauna was present, and the mud core suggests that a fleshy algal meadow (grass bed analogue) was the major reef environment, requiring shoal water just as the phylloid algae do. No evidence of hydromechanical accumulation exists in the main core, and no framework was developed. Capping deposits are skeletal sand. This reef was clearly subaerially exposed at its top; massive dissolution effects are present.
Framework reefs are relatively sparse in the Paleozoic geologic record, although the Capitan reef (Permian) exposed at Walnut Canyon, White's City, New Mexico, is an example. Unfortunately, the exposure contains only the upper portion of the reef. It is mainly a sponge reef but was tightly cemented during growth to provide a framework. Little interstitial sediment exists, leading to the interpretation of sediment winnowing by slope and mechanical wave and current impingement (Pray, 1987).
Reefs can be modeled on a small scale using a reef architectural sequence developed from modern reef processes. Application of this architectural sequence to ancient reefs makes it possible to identify similar architectural sequences and to model sea-level rates of rise and sediment accumulation. Development of three architectural classes discriminates significance of carbonate aggradation (framework class), degradation (biodetrital class), and transportation of carbonate grains (hydromechanical class). Absolute rates of modern sea-level change and reef growth may not be analogous to ancient reefs, but there is no evidence to the contrary. Modern reefs are comparable in geographic scale to ancient reefs, unlike most carbonates.
Comparison of ancient reef fabrics to the ideal architectural sequence shows that Pennsylvanian reef sea-level relationships, among others, can be interpreted from rock sequences. Computer modeling is less advanced. Initial computer models were operated on a one-dimensional model with general success. However, current efforts are underway to simulate Holocene sea-level rise and reef growth rates from St. Croix. The object of the modeling is to predict porosity occurrence through a better understanding of the environmental controls on fabric evolution.
Adey, W. H., 1975, The algal ridges and coral reefs of St. Croix--their structure and Holocene development: Atoll Research Bulletin, v. 187, p. 1-67
Adey, W. H., and Burke, R. B., 1976, Holocene bioherms (algal ridges and bank-barrier reefs) of the eastern Caribbean: Geological Society of America Bulletin, v. 87, p. 95-109
Adey, W. H., and Burke, R. B., 1977, Holocene bioherms of Lesser Antilles-geologic control of development: American Association of Petroleum Geologists, Studies in Geology 4, p. 67-81
Adey, W. H., Macintyre, I. G., and Stuckenrath, R., 1977, Relict barrier reef system off St. Croix--its implications with respect to late Cenozoic coral reef development in the western Atlantic: Proceedings of the Third International Coral Reef Symposium, p. 15-21
Burke, R. B., and Gerhard, L. C., 1987, Reefs, bioherms and banks--a semantic and genetic continuum (abs.): American Association of Petroleum Geologists, Bulletin, v. 71, p. 535
Burke, R. B., Adey, W. H., and Macintyre, I. G., 1989, Overview of the Holocene history, architecture, and structural components of Tague reef and lagoon; in, Terrestrial and Marine Geology of St. Croix, U.S. Virgin Islands, D. K. Hubbard, ed.: West Indies Laboratory, Teague Bay, St. Croix, U.S. Virgin Islands, Special Publication 8, p. 105-109
Davies, P. J., and Marshall, J. F., 1985, Halimeda bioherms--low energy reefs, northern Great Barrier Reef: Proceedings of the Fifth International Coral Reef Congress, v. 5, p. 1-7
Fagerstrom, J. A., 1987, The evolution of reef communities: John Wiley and Sons, New York, 600 p.
Folk, R. L., 1959, Practical petrographic classification of limestones: American Association of Petroleum Geologists, Bulletin, v. 43, p. 1-38
Franseen, E. K., and Watney, W. L., eds., 1989, Sedimentary modeling--computer simulation of depositional sequences: Kansas Geological Survey, Subsurface Geology Series 12, p. 31-34
Gerhard, L. C., 1978, Reconnaissance study of sediment facies of Salt River estuary, St. Croix, US VI--mixed carbonates and clastics: Mountain Geologist, v. 15, no. 3, p. 109-123
Gerhard, L. C., 1981, Origin and evolution of Sand Cay (Candlelight Reef), US Virgin Islands: Atoll Research Bulletin, no. 242, p. 1-11
Gerhard, L. C., and Burke, R. B., 1986, Reefs, bioherms and banks--a semantic and genetic continuum: Abstracts of the 12th International Sedimentological Congress, p. 118
Gerhard, L. C., and Burke, R. B., 1990, Reefs, bioherms and banks--a semantic and genetic continuum: Kansas Geological Survey, Open-file Report 90-11, 23 p. [available online]
Gerhard, L. C., and Stolzman, R., 1974, Sedimentology of Boiler Bay, St. Croix; in, Guidebook to the Geology and Ecology of Some Marine and Terrestrial Environments, St. Croix, US VI: West Indies Laboratory, Teague Bay, US Virgin Islands, Special Publication 5
Gray, R. S., 1967, Cache Field--a Pennsylvanian algal reservoir in southwestern Colorado: American Association of Petroleum Geologists, Bulletin, v. 51, p. 1,959-1,978
Hine, A. C., Hallock, P., Harris, M. W., Mullins, H. T., Belknap, D. F., and Japp, W. C., 1988, Halimeda bioherms along an open seaway--Miskito Channel, Nicaraguan Rise, SW Caribbean Sea: Coral Reefs, v. 6, nos. 3/4, p. 173-178
Hubbard, D. K., Burke, R. P., and Gill, I. P., 1985, Accretion in deep, shelf-edge reefs of St. Croix, US VI; in, Deep-Water Carbonates, P.D. Crevello and P. M. Harris, eds.: Society of Economic Paleontologists and Mineralogists, Core Workshop 6, p. 491-527
James, N. P., 1983, Reef environment; in, Carbonate Depositional Environments, P. A. Scholle, D. G. Bebout, and C. H. Moore, eds.: American Association of Petroleum Geologists, Memoir 33, p. 345-440
Koerschner, W. F., III, and Read, J. F., 1989, Field and modeling studies of Cambrian carbonate cycles, Virginia Appalachians: Journal of Sedimentary Petrology, v. 59, p. 107-110
Lawrence, D. T., Doyle, M., and Aigner, T., 1990, Stratigraphic simulation of sedimentary basins--concepts and calibration: American Association of Petroleum Geologists, Bulletin, v. 74, p.273-295
Longman, M. W., 1981, A process approach to recognizing facies of reef complexes; in, European Fossil Reef Models, D. M. Toomey, ed.: Society of Economic Mineralogists and Paleontologists, Special Publication 30, p. 9-40
Lowenstam, H. A., 1950, Niagaran reefs of the Great Lakes area: Journal of Geology, v. 58, p. 430-487
Macintyre, I. G., and Glynn, P. W., 1976, Evolution of modern Caribbean fringing reef--Galeta Point, Panama: American Association of Petroleum Geologists, Bulletin, v. 60, p. 1,054-1,072
Macintyre, I. G., Multer, H. G., Zankl, H. L., Hubbard, D. K., Weiss, M. P., and Stuckenrath, R., 1985, Growth and depositional facies of a windward reef complex (Nonesuch Bay, Antigua, WI): Proceedings of the Fifth International Coral Reef Congress, v. 6, p. 605-610
Marshall, J. F., and Davies, P. J., 1988, Halimeda bioherms of the northern Great Barrier Reef. Coral Reefs, v. 6, nos. 3/4, p. 139-148
Milliman, J. D., 1974, Marine carbonates: Springer-Verlag, Berlin, 375 p.
Neumann, A. C., and Macintyre, I., 1985, Reef response to sea level rise--keep-up, catch-up or give-up: Proceedings of the Fifth International Coral Reef Congress, v. 3, p. 105-109
Orme, G. R., and Salama, M. S., 1988, Form and seismic stratigraphy of Halimeda banks in part of the northern Great Barrier Reef Province: Coral Reefs, v. 6, nos. 3/4, p. 131-138
Phipps, C. V. G., and Roberts, H. H., 1988, Seismic characteristics and accretion history of Halimeda bioherms on Kalukalukuang Bank, eastern Java Sea (Indonesia): Coral Reefs, v. 6, nos. 3/4, p. 149-160
Pray, L. C., 1987, Capitan reef complex (Permian), Guadalupe Mountains, southwestern United States--a classic sedimentologic model in flux (abs.): American Association of Petroleum Geologists, Bulletin, v. 71, p. 1,441-1,442
Pray, L. C., and Wray, J. L., 1963, Porous algal facies (Pennsylvanian), Honaker Trail, San Juan Canyon, Utah; in, Shelf Carbonates of the Paradox Basin, R. O. Bass and S. L. Sharps, eds.: Four Comers Geological Society, p. 204-234
Roberts, H. H., and Aharon, P., and Phipps, C. V., 1988, Morphology and sedimentology of Halimeda bioherms from the eastern Java Sea (Indonesia): Coral Reefs, v. 6, nos. 3/4, p. 161-172
Zankl, H., and Shroeder, J. H., 1972, Interaction of genetic processes in Holocene reefs off North Eluthera Island, Bahamas: Geologische Rundschau, v. 61, p. 520-541
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version April 21, 2010. Original publication date 1991.
URL=http://www.kgs.ku.edu/Publications/Bulletins/233/Gerhard/index.html