Kansas Geological Survey, Open-file Report 90-11
by
Lee G. Gerhard1 and Randolph Burke2
1Kansas Geological Survey
2North Dakota Geological Survey
KGS Open File Report 90-11
What the profession has historically and collectively called "reefs" have been variously described by physical and biological characteristics of their surficial cover (modern reefs), degree of cementation or rock geometry (ancient reefs) and geographic setting (both). Processes of carbonate generation, degradation and transportation determine the biotic composition, fabric and architecture of reef bodies, both modern and ancient, and thus can be the basis of reef definition and classification. In this scenario, balancing of the three processes leads to classification of reefs in an architectural continuum, portrayed as points or traces on ternary diagrams. Framework, biodetrital and hydromechanical reefs are the three end members of the classification.
Rates of sea-level-change and physiographic setting determine fabric response to sedimentary processes. Porosity evolution is related to primary architecture as well as to early diagenesis. Most petroleum produced from the broad spectrum of reefs is from the types classified here as biodetrital and hydromechanical reefs.
The writers are indebted to Robert H. Shaver, Indiana Geological Survey, and Timothy A. Cross, Colorado School of Mines, for their extended discussions of reefs, sedimentation, and sea-level changes. They each reviewed this manuscript in detail; the manuscript is much improved by their contributions and we appreciate their help.
Reef definition has been a singularly unproductive effort. Most studies consider organic taxonomy and community succession as the keys to knowledge and thus definition. Our purpose is to approach reef definition through architecture, genetic processes and sedimentary settings, which in turn permit us to understand and predict the evolution and ultimate character of fossil reefs.
Several recent papers rigorously define and limit the use of "reef" to those occurrences in which organic framework is architecturally predominate within a three-dimensional organic structure (Fagerstrom, 1987; James and Macintyre, 1985; Reid and Ginsburg, 1986). Although such definitions are valid, they do not lead to functional comprehension of the predictable relationships between depositional slope, biotic activity, mechanical energy, and rate of sea level changes and thence to generation, degradation and transportation processes. These relationships appear to forecast degrees and timing of cementation,vertical and lateral sequences of facies and sequence of sedimentary fabrics, other diagenetic history and porosity/permeability occurrence.
Heckel (1974) and Milliman (1974) have synthesized a great volume of literature on reefs. Heckel stressed the need for "wave-resistance" in his discussion and definition of reefs, thus emphasizing frameworks, encrusting and binding. Milliman also adopted the "rigid framework" concept in his work, and amplified that view in field discussions with the first author. Reid and Ginsburg (1986) applied the framework concept to their study of Triassic reefs, basing their determination of reefs versus nonreefs upon the presence of framework. James and Macintyre (1985) specifically required framework to define reefs in their discussion of reef organisms and growth. In early definitive studies, Lowenstam (1950) and Textoris and Carozzi (1964) specified framework as a basis of defining reef "core" pertaining to the Silurian reefs of the Illinois basin. They also suggested that there was an evolutionary sequence of development of the reefs, pointing to changes in architecture and internal structure. Textoris and Carozzi (1964) suggested multiple stages of reef development related to (mechanical) energy levels (see Shaver and Sunderman, 1989 for a critique of their evaluations) .
Sloss (1962) recognized that shapes of siliciclastic sedimentary bodies are controlled by the quantity, dispersal and nature of materials of which the deposits arc, formed and the receptor value (subsidence) of the basin in which they accumulated. His vision is applicable to the carbonate world and has been gradually incorporated into modern carbonate thought. For instance, Longman (1981) applied a process approach, but he suggested that modern reefs are only a few percent framework and that ancient reefs have more framework.
Other workers analyzing ancient reefs have recognized a role for sea-level changes. Neumann and Macintyre (1985) considered organism growth rates as a reflection of sea-level changes, but concentrated on the organisms rather than on processes controlling organism growth and morphology. Several papers, cited subsequently, have recognized the multiplicity of processes which control the geometry and fabric of some ancient reefs. Toomey (1970) wrote a perceptive analysis of upward changes in growth habit for an Ordovician reef recognizing the relationships between water depth and reef development.
In summary, existing literature supports a ruling hypothesis that "reefs" are framework-dominated organic structures. The ruling hypothesis is flawed by failing to recognize that individual reefs evolve, producing an architectural sequence of diverse fabrics. Framework development is a function of rates of growth, degradation and transportation, which change in response to depositional slope and rates of sea-level changes. Thus the degree of framework development has no inherent attribute for reef definition. Further, definitions based upon the ruling hypothesis have not gone beyond description, and have not been useful for effective prediction of facies and of diagenetic porosity development in subsurface settings.
It is our thesis that all three-dimensionally discrete bodies of organically derived carbonate rocks are "reefs," that they result from the interplay of definable major sedimentary processes, and that their depositional and diagenetic settings can be interpreted from observing their vertical sequence of fabrics. Reef fabrics and internal architecture evolve in response to changing rates of sea-level-rise. Availability of organisms within a specific reef setting and interruptions in this evolution may determine final architecture.
Part of our thesis is that the rates at which the processes of reef generation operate will determine the fabric of any reefs formed. As a corollary, fabrics of sediment (rocks) generated at a reef site can be interpreted with respect to the rates and balances of processes that operated.
We have deciphered a continuum of fabrics and architectural styles which are controlled more by sedimentary processes than by biotic evolution or community succession. Fabrics thus result from the interplay of organic growth, dynamic processes and depositional topography.
Our hypotheses are not complete; considerable field work and testing remains, but they have held up well to literature and preliminary field testing and are advanced here for discussion and comment.
It is our deliberate choice to use the term "reef" as a general term with appropriate architectural modifiers, rather than "bank" or "bioherm," in order to stress the inherent continuum of fabrics and processes we have interpreted from our studies. At one end of the continuum are "framework" reefs, so well illustrated by so many of our colleagues; at the other end are wave-initiated piles of skeletal carbonate sand. Biotic generation, degradation and mechanical transportation interweave to provide reef fabric. The rates at which these processes operate seem to be controlled by rates of sea-level change, especially rates of rise. The mix of primary mineralogies may determine ultimate porosity of a reef through early diagenesis.
Carbonate sediment is a product of growth of carbonate skeletons which are modified by biologic and mechanical degradation, providing sediment of varying size and shape. Sediment so generated is either incorporated into the reef or transported out of it. The balance between skeletal growth and degradation and transportation processes determines the reef body fabric and architecture, and ultimately, the type and degree of diagenesis. The degree of transportation of detritus out of a reef system is a function of wave energy and depositional slope. Transportation of detritus from a reef system creates the opportunity for framework to develop and for syngenetic cementation. Lack of sediment removal provides smaller pore space, and inhibits skeletal growth and syngenetic cementation. Production of carbonate sediment within an active reef system is so rapid that frameworks cannot develop unless the sediment is removed; frameworks can literally drown in their own detritus. In the opposite extreme, a hydromechanic pile of loose sediment will be the result of transportation or sweeping of carbonate sediment into a wave-induced barform.
Effects of the three major reef-construction processes, skeletal generation, degradation and transportation, can be graphically represented on a ternary diagram (fig. 1). A corresponding reef classification terminology, framework, biodetrital, and hydromechanical, is derived from the processes (fig. 2).
Figure 1--Ternary diagram showing the three major carbonate processes that determine reef architecture. Connecting plotted points representing individual samples will trace the evolution of process influence on the reef.
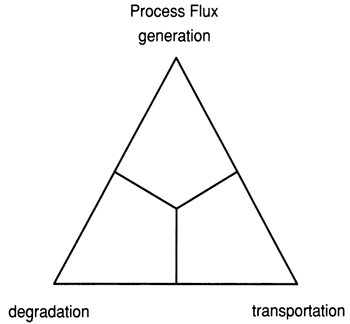
Figure 2--Ternary diagram of three major reef classes based on architecture developed by processes shown in figure 6. There is a correspondence between points of the two ternary diagrams, that is, framework reefs are dominated by generation processes, biodetrital reefs by degradation, and hydromechanical reefs by transportation.
Eustasy is the remaining dynamic process that must be considered as controlling reef fabric. Neumann and Macintyre (1985) have interpreted part of this relationship in their "catch-up, keep-up,or give-up" view of succession. However, while addressing organism response to sea-level changes, they did not consider the sequence of fabrics which is coincident with a normal cycle. Normal cycles, characterized as sine curves, have rapidly changing rates of rise, which, in conjunction with relatively stable organism growth rates, provide large variations in absolute water depth. We have not considered whether sea-level changes are true global eustasy or of local origin.
Studies of modern coralgal reefs have provided insight into the processes which control the development and architecture of ancient reefs. Although modern carbonate analogs generally suffer smaller scale than their ancient counterparts, for individual reefs scales are not significantly different. Modern reefs have characteristic fabrics and architectures (framework, biodetrital, hydromechanical) that reflect their settings and the balance of processes that create them. Core-drilling provides extensive information about the vertical sequence of fabrics in modern reefs.
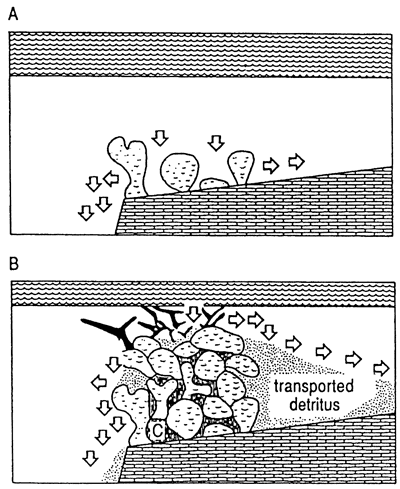
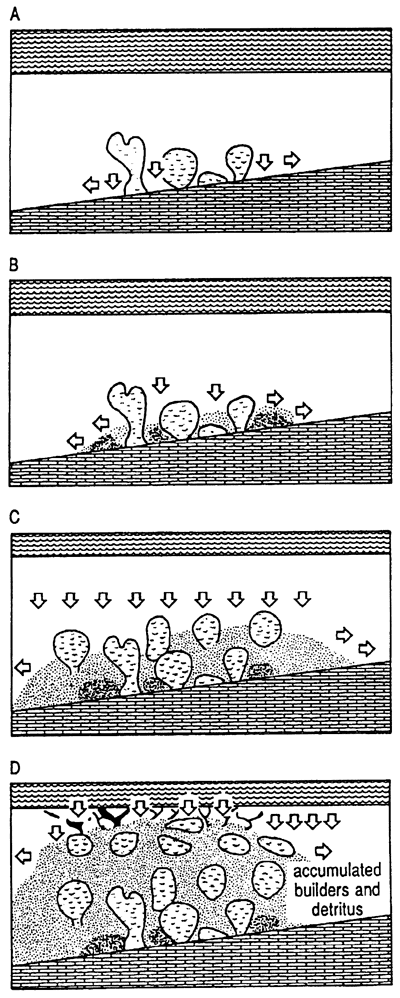
Macintyre and Glynn (1976) drilled on a transect across the fringing reefs at Galeta Point, Panama, where a Holocene reef complex is developed upon a Miocene substrate. These framework reefs are adjacent to a steep slope that descends from sea level to at least fourteen meters within one hundred meters away from the reef crest. The complex is under the direct influence of strong trade winds. In this exposed, high-mechanical energy setting, corals form the bulk of the reef and syngenetic cementation apparently is quite extensive, occurring most abundantly in the seaward portions of the reef. Characteristics of this reef setting and resultant reef architecture are steep depositional slope, high mechanical energy, framework and syngenetic cementation (fig. 3). Around the island of St. Croix, U.S.V.I., (location of most of the cited work), in a few exposed high-depositional slope settings, reef formation is similar to that described in Panama by Macintyre and Glynn (1976) in that framework development is predominate and syngenetic cements exist.
Figure 3--Schematic diagram showing typical setting and fabric of framework reef. Reef organism detritus is cleaned by flushing, permitting cements to precipitate in lower energy interstices. Shelf-edge settings enhance flushing and propensity for framework generation.
In other settings with differing balances of processes, other characteristics develop. Extensive coring of reefs through the eastern Caribbean, such as by Adey (1975), Adey and Burke (1976, 1977), Adey et al. (1977), Macintyre et al. (1985), and Hubbard, Burke and Gill (1985), has provided a view of the architecture of modern coralgal reefs that permits further inferences as to the relationships among fabrics, processes, and settings.
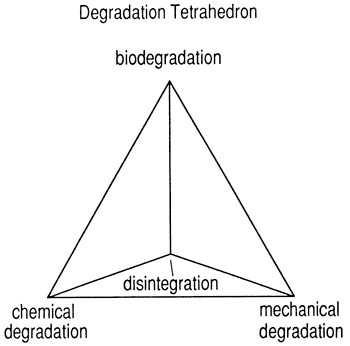
In most St. Croix reefs, where depositional slope becomes gentler with consequent lessened wave energy, a different architecture is developed. In this setting, reefs are composed predominantly of sediment; commonly, 60 to 70 percent of the mass is biodetrital sand or finer material. Only a thin veneer of living coral appears to form framework. Biodetrital sediment is produced within the reef mainly by bioerosion of living and dead skeletal material (Zankl and Schroeder, 1972) (fig. 4), but also by mechanical trituration. Wave and current energies are not high enough to sweep the biodetritus from the reef, nor is the depositional slope steep enough to cause extensive downslope transport of sediment. In effect, the reef framework organisms drown in their own detritus; growth and reproduction to form an interlocking framework is inhibited by the large amount, of detritus deposited within the reef. Individual organisms are widely spaced and thus prevented from interlocking.
Figure 4--Tetrahedral diagram showing four major degradation processes. A reef system detrital accumulation can be plotted as a point of balance of the four processes. As carbonate-secreting organisms grow, their skeletons are subject to constant degradation by boring organisms while alive, and biologic, chemical and mechanical degradation upon death.
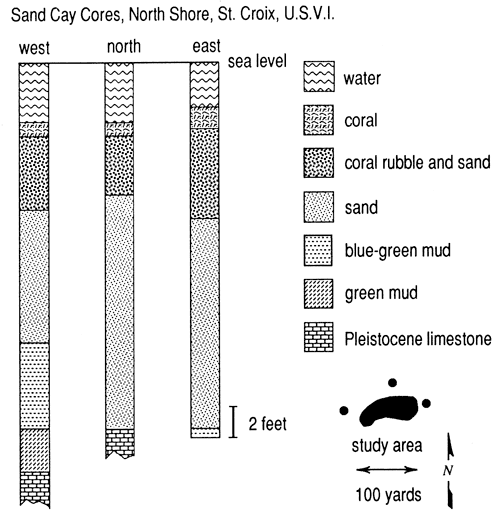
Cores taken at the western end of Teague Bay reef (Sand Cay) on St. Croix (Gerhard, 1981) further demonstrate this relationship, as do the cores of Adey and Burke from Martinique (Adey and Burke, 1976, figs. 12 and 13). The Sand Cay cores show biodetritus deposited on pre-Holocene limestone or Holocene fine-grained siliciclastic sediments; only a thin veneer of living reef coral or large rubble is present (fig. 5). No cementation has been observed in this setting, and pores are abundant but small, as compared to the framework-abundant reefs previously described. The polytaxonomic origin of the reef pile indicates that the sediment is mostly aragonite and magnesium calcite with fewer grains of calcite.
Figure 5--Logs of holes drilled at Sand Cay, St. Croix, U. S. Virgin Islands. Logs show that the reef was established as a sand accumulation, then rubble and sand, and finally, a living capstone of framework reef. This took place in a sea-level rise that has slowed in rate over the last 7000 years.
Characteristics of this reef setting and resultant reef architecture are moderate depositional slope; moderate mechanical energy; the resulting reef is a biodetrital sediment mass that has little or no framework and virtually no cementation (fig. 6).
Figure 6--Schematic diagram showing typical setting and fabric of biodetrital reefs. Sediment is generated in the reef system but is only partly transported out of the reef system. Consequently, coral skeletons appear to float in debris of sand and rubble of other size; reefs may drown in their own debris. Interstices are filled with sediment, substantially decreasing probability of syngenetic cementation and growth. Mixed mineralogies and grain sizes of these systems enhance secondary porosity development.
A third generalized reef setting and balance of processes exists, characterized by standing or oscillatory wave sets, low depositional slope, mechanical trituration of skeletons and, possibly, longshore currents. In this setting, accumulation of organically generated carbonate sediment predominates by hydromechanical "piling," perhaps later colonized by benthic skeletal animals and plants. Although many students of carbonate sedimentology would argue that such wave- and current-generated, three-dimensionally discrete bodies of organically generated but hydro mechanically reworked sediment are not reefs, we view these as simply one end member of a continuous architectural gradation.
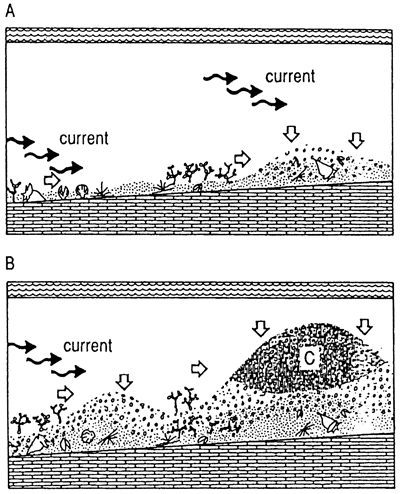
Davies and Marshall (1985), among others (Hine et al, 1988; Marshall and Davies, 1988; Orme and Salama, 1988; Phipps and Roberts, 1988; Roberts, et al, 1988), documented hydromechanically formed piles of Halimeda (bioherms, in their terminology) behind the main coral reef fronts of the Great Barrier Reef, Australia, and in other localities in the Pacific and Caribbean. They have shown that the algal plate mounds within these reef complexes are significantly older than the framework coral reefs themselves. They attributed initiation of algal accumulation to upwelling onto the pre-existing Pleistocene limestone platform, which dips gently westward from the front of the coral reefs. However their seismic profile of geometry and amplitude of the banks strongly suggests hydromechanical accumulation in addition to in situ growth. Little seismic evidence exists for current-bedded internal structure, but bioturbation tends to be high in shallow water Halimeda accumulations in St. Croix (Gerhard, 1978). High productivity of Halimeda can be attributed to upwelling, but such settings also provide the necessary mechanical energy to accumulate low-density algal plates. Algal and other skeletal grains of the present surface appear to be mostly autochthonous to the accumulations. Additional growth is taking place preferentially on tops of the wave-generated mounds, likely in response to higher ambient light conditions (less turbidity) which enhance photosynthesis (see also discussion in Gerhard, 1978). No doubt the reefs have been colonized by Halimeda throughout their growth, so that in situ algal plates are combined with mechanically added sediment grains to form the present geometry. At present wave trains are cut off by the coral reef front, and only low wave energy impinges on these reefs. These algal bodies are similar to Carboniferous algal reefs, particularly of the Midcontinent, Permian basin and Paradox basin, United States.
Characteristics of this reef setting and resultant reef architecture are very low depositional slope, variable but frequently low mechanical energy, hydromechanical accumulation of reef grains and little or no syngenetic cementation (fig. 7). When algae or other sediment-producers are prolific, grains will be trapped in situ. Hydromechanical reefs do not generally provide a suitable substrate for the settlement of framework organisms, as the grains are subject to both bioturbation and wave and current reworking. Not all hydromechanical reefs are of algal origin.
Figure 7--Schematic diagram of hydromechanical reef showing the accumulation of organic debris in a wave-dominated regime. Establishment of a productive community either on or nearby the reef accumulation with sufficient constancy of wave and current geometry and intensity can provide necessary skeletal materials. Although an extreme example of this reef type could be a carbonate-sand bar, most are propagated by vegetative communities.
As reefs of any framework percentage grow, a certain amount of detritus is generated by biological and hydromechanical degradation and accumulates within the reef. If this detritus is not removed from the reef, it will to some degree inhibit organic skeletal. growth. If detrital generation is high and transportation of detritus out of the reef is low, the accumulation of detritus (bioclasts) will provide a significant or major part of the reef mass in comparison with amount of inter-locking framework, establishing a biodetrital reef. In the extreme, the reef will develop as a three-dimensionally discrete body of organic detritus, a hydromechanical reef.
In contrast, if the reef setting is such that waves and currents winnow the degradation products (biodetritus) from the reef, then the reef will be characterized by organic skeletons and open space, encouraging the establishment of well-developed framework. From this one can infer that fast growing reef organisms in a sediment-swept setting will most likely produce framework(framework reefs).
Rates at which these processes operate are controlled by various chemical, thermal and mechanical attributes of the sedimentary environment, which are not discussed here. Instead, we focus on two additional attributes which are not as commonly discussed are those of substrate geometry and rates of sea-level change.
Substrate geometry can be a major factor determining reef fabric, because the depositional slope and topography modulate the wave and current regime of the ocean. Shelf-edge reef locations with steep declivities promote growth of framework-generating organisms and concurrent removal of detritus from the reefs. Uninhibited wave trains impinge upon shelf-edge settings, transferring potential energy directly to the reef mass rather than gradually transferring energy to the bottom and reefs on the lesser slopes of platforms.
From our interpretations we conclude that reef fabrics are gradational and that reef classification should recognize the continuum of architecture. Three major reef types can serve as end members of a ternary classification system (fig. 2). These end members are directly comparable to the process ternary end members that form the basis of their diagnosis (fig. 1). This permits an interpretation of the physical and biologic setting of a reef preserved through its fabrics. The three reef class names, framework, biodetrital and hydromechanical, are descriptive, evolve from sedimentary and oceanographic processes, and are continuous variables. For these reasons, the nomenclature ternary is divided into fields which permit the grouping of similar, but not identical, reefs. Others may wish to substitute reef for framework,bioherm for biodetrital, and bank for hydromechanical, but we regard our terms as more descriptive, better related to the objects described, and more useful in the diagnosis of ancient reef evolution.
Most classifications and terminology sets fail to recognize the growth sequence modern reefs and their ancient counterparts have undergone, which frequently carries them from the hydromechanical class through biodetrital and sometimes to framework. Current reef nomenclature assumes stable environmental conditions during reef growth, thus homogeneous reef fabrics, i.e., reef, bank, bioherm. Not all reefs go though the complete metamorphosis of fabrics, but sufficient numbers do so as to permit development of an idealized reef architecture/nomenclature sequence. Lowenstam (1950, fig. 8, 10) illustrated his concept of reef development by illustrating the core of Silurian reefs as vertically continuous and homogeneous, although a variety of different fabrics and organisms occupied the flanks. Textoris and Carozzi (1964) suggested that the same Silurian reefs underwent a six-stage development based upon inferred water depth, taxons of skeletal particles, and mechanical energy levels. Neither recognized the more universal relationships of architectural change, but corings of modern reefs were not available to them nor was there a well-developed body of literature about sediment/organism interrelationships.
The modern St. Croix reef complex includes three fabrics: first is initially pure sand (hydromechanical), followed upward by a thick sequence of corals (or other framebuilders) imbedded in sand (biodetrital) and then covered with a veneer of branching corals (framework) that are still living. Adey and Burke (1976) have provided the most detailed data set of modern reefs which characterizes this idealized sequence. From their reef coring program in the Caribbean they were able to determine ages for the sampled corals as well as their position within the reef sequence. These data were then compared to sea level history for the region to provide a time-depth data series (fig. 8). Cores drilled on Sand Cay, St.Croix, U. S. Virgin Islands (Gerhard, 1981) add detail of reef architecture to the data of Adey and Burke to illustrate the sequence of fabrics typical of these coralgal reefs (fig. 5). Reef fabrics progressed through the sequence of nomenclature in response to changing environmental conditions, mainly water depth. Water depth changes result from changes in the rate of sea-level, rise relative to reef accretion rates.
Figure 8--Time-depth sea-level curve for St. Croix, U.S. Virgin Islands (modified from Adey and Burke, 1976).
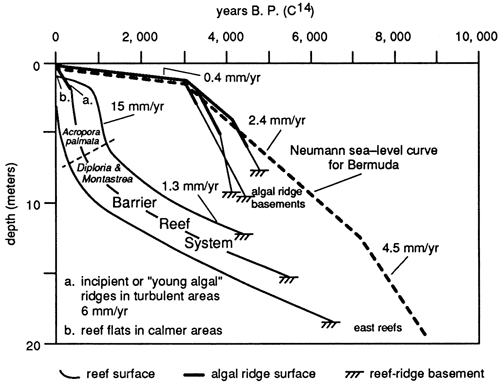
Holocene sea-level rise postulated by Adey and Burke (1976) is asymmetric; rapid sea level rise is followed by ever slowing rates of rise. Absolute water depth is not congruent with the rate of sea level-rise rate of; rather, maximum water depth occurs near the inflection point from the greatest rate of rise to a moderate rate of rise, reflecting rates of sedimentation and reef-organism growth (fig. 9).
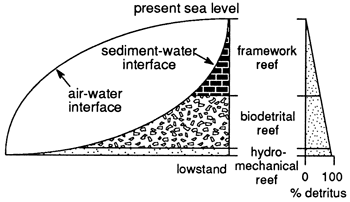
Figure 9--Schematic diagram showing the relationships of sea-level change to absolute water depth, detritus generation, sedimentation and reef architecture. Left portion shows reef classes plotted against sea level (air-water interface) and the sediment-water interface. At lowstand a hydromechanical reef initiates reef development in very shallow water. Absolute water depth increases as rapid sea rise takes place, where biodetrital reef growth is initiated. When the biodetrital reef growth rate exceeds rate of sea-level rise, absolute water depth decreases. Finally, when the reef grows sufficiently shallow to permit rapid-growing reef framework organisms to thrive, a framework reef architecture is developed and reaches surface. On left, schematic shows the abundance of detritus in the reef decreases upward with change in reef architectural class.
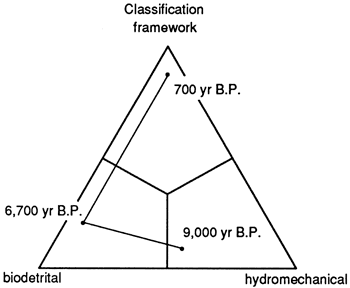
The Teague Bay reef system, studied by Adey and Burke (1976) and Gerhard (1981), is located on the landward margin of the St. Croix platform. No shelf declivity is present,, although there is a deeper reef along the shelf edge margin that appears to be relatively free of detritus. Characterization of Teague Bay reef requires specification of point in time, water depth and sediment level in vertical section. One easy method of analyzing the evolution of the reef (and its resultant nomenclature) is to plot reef fabric through time on a ternary diagram (fig. 10). It appears that framework reefs are a special result of lowered rates of sea-level-rise (apparently at the conclusion of a rise); subaerial erosion coincident with relative lowering of sea level may well destroy many of the framework veneers, preserving only their earlier biodetrital or hydromechanical fabrics.
Figure 10--Ternary diagram of reef classification based upon reef architecture, with plot of samples and consequent interpretation of evolution. Based on data in Adey and Burke (1976) from St. Croix, U. S. Virgin Islands. Reef was initiated as a hydromechanical reef with stabilizing head corals about 9000 ybp., then gradually evolved to a full biodetrital reef with minor framework corals about 6700 ybp., and finally established a mechanical framework about 700 ybp.
Initial Teague Bay reef growth was as a sand-dominated bank with relatively small numbers of corals, coincident with the initial sea level rise over the exposed Pleistocene substrate. After initial colonization, water depths rapidly increased during the highest rate of sea-level rise (4.5 mm/yr.), from approximately 8000 ybp to 7000 ybp. Initial hydromechanical accumulations were supplanted by coral-head-dominated biodetrital reefs, but water depths continued to increase because sea level rise from 7000 to 3200 ybp. was greater than sediment accumulation rates (1.3 mm/yr). Slopes were too gentle and water depth too great for waves of normal amplitude to winnow sediment from the reef. At about 3200 ybp the rate of change of sea-level rise dropped to about 0.4 mm/yr, whereas the sediment-accumulation rate was maintained at 1.3 mm/yr. Significant accommodation potential existed for continued growth. Head corals and detritus characterized reef accretion as sedimentation and organic reef growth exceeded the lowered rate of sea level rise. Thus, absolute water depths decreased.
At about 1200 ybp water depths were shallow enough that Acropora palmata could establish on the reef. This coral is productive only in shallow water and has growth rates in excess of a centimeter a year today on the same reef. Prolific A. palmata growth created a veneer of framework on the upper surface of the Teague Bay reef that breaks surface today, although the lower slopes of the reef remain, nevertheless, a biodetrital reef. Much of the detritus generated within the A. palmata zone is washed landwards into the backreef or down the foreslope, permitting a thin framework to exist.
Ancient reef fabrics have followed the same development patterns as modern reefs. Framework, biodetrital and hydromechanical classes are all present, although most three-dimensional organically derived rock bodies most frequently display hydromechanical and biodetrital characteristics. Full framework development is uncommon even at the tops of ancient reefs. Literature descriptions of ancient reefs rarely are sufficiently detailed to ensure full diagnosis of the fabrics and architecture, but the general descriptions available permit categorization into major classification "fields." Our current research concentrates on development of detailed descriptions from a variety of types, ages and geometries of ancient reefs and testing of our concepts of fabric evolution and architecture. Examples from the literature and limited new field descriptions are the basis for the following discussion of ancient reef classification and paragenesis. Our discussion is regarded as preliminary and incomplete because of the limited data now available.
The Capitan Reef core, as exposed at the mouth of Walnut Canyon, White City, New Mexico, is an example of a fossil framework reef. Described in detail by many authors, this reef's organic constituents are mostly sponges with associated calcareous algae and other organisms of uncertain affinities (Tubiphytes). Little detritus exists between organism skeletons (fig. 11), instead, the space between skeletons is filled with fans of cement, assumed to be originally submarine aragonite(Pray, 1987). In situ sponges, lack of major organic debris, and presence of major amounts of interstitial cements leads to interpretation of very early cementation, steep depositional topography and original framework.
Figure 11--Photomicrograph of submarine cements in framework reef, Capitan Formation, Walnut Canyon, White City, New Mexico. Note radiating crystals, assumed to have been originally aragonite. Field of view is approximately 2.5 cm. wide.

Early cementation of the Capitan reef obliterated any primary porosity and probably initiated replacement of the primary skeletal structures. Retention of the primary cement fabrics through later paragenesis indicates that no effective permeability ever existed, and the reef remains impermeable now. Several writers (such as Moore and Wilde, 1986), have illustrated development of extensive erosional surfaces between episodes of submarine deposition, reef growth and cementation. We suggest that early cementation of framework fabric is typical and may diminish the potential for porosity and permeability development iii later paragenetic stages. Detailed description of a full Capitan lithologic sequence has not been available, so that evolution of fabric is uncertain.
Pliocene reef sets on the south-central margin of St. Croix, U. S. Virgin Islands, are typical biodetrital reefs, apparently very similar in overall fabric to the contemporary reefs of Teague Bay described previously. Corals, mollusks, and smaller organisms form a mass of organic skeletons interspersed in detrital carbonate sediment, and forming a three-dimensional bay-barrier reef configuration (fig. 12).
Figure 12--Pliocene biodetrital reef on St. Croix, U. S. Virgin Islands, showing the massive nature of the reef. Cavities represent the former presence of large coral heads.

Initial Pliocene sedimentation included reworking of exposed Miocene foraminiferal wackestones and packstones during early sea-level rise, followed by development of red algal hardgrounds. The hardgrounds appear to represent the maximum water depths. Shelf and reef-associated rocks were subsequently deposited. At least two reefs existed at the same locality, both parallel to the modern strandline. Subaerial erosion and diagenesis is evident in exposure between an upper and lower reef.
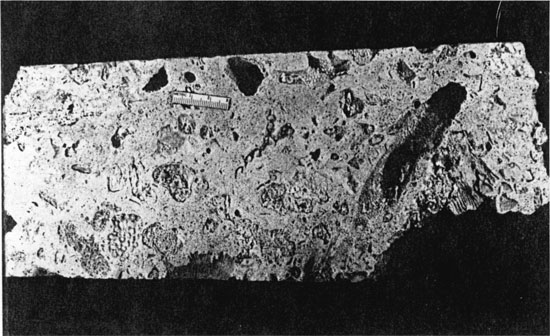
One of the most interesting features of this biodetrital reef is its paragenesis. This biodetrital reef also indicates that multistage diagenesis provides for the development of secondary porosity. Much of the carbonate in a modern coralgal reef, whatever type, is aragonite. Since aragonite is metastable in fresh water, any exposure of a permeable reef to meteoric waters can enhance dissolution of coral, mollusk, and codiacean algal grains. Our example of this early diagenesis and calcitization occurred in the Pliocene of St. Croix, where a biodetrital reef core is elevated some 40 feet above present sea level. Detritus matrix was cemented relatively early by calcite, aragonite was later removed by vadose dissolution. Resulting voids remain open. Mollusk steinkerns are abundant, but most spectacular are the large single coral cavities remaining, each containing a threaded network of filled burrows and borings from the original coral. This negative print provides a detailed view of the biodegradation process (fig. 13). We believe that this paragenesis is common in biodetrital reefs, that such reefs are more amenable to complex paragenesis and are more likely to have effective permeability for petroleum production.
Figure 13--Detail of specimen showing bioerosion traces and diagenetically evacuated coral and mollusc molds. Scale in centimeters.
Development of piles of modern organic carbonate debris to form three-dimensional bodies of sediment has been demonstrated previously. Fossil examples are phylloid algal reefs, Komia reefs, and skeletal carbonate sand bars. For example, Cisco Formation reefs of Lea County, New Mexico, drilled in the 1960's, appear to be clean piles of carbonate sand. We have studied no cores of these particular reefs, therefore our discussion will focus on phylloid and Komia reefs.
Some well-exposed phylloid reefs occur in the Ismay Zone of the Hermosa Formation, along the San Juan River between Bluff and Mexican Hat, Utah (fig. 14). Although very detailed measured sections are not yet available, reconnaissance study of fabric evolution and reef spacing provide the following observations. Reef spacing appears to be relatively even along the river exposures, suggesting a wave period spacing origin (Brinton, 1986, Plate 1). Reef crests are at the same elevation, suggesting growth to sea level. That may be partly a result of subaerial erosion at the conclusion of reef growth, but in the one exposure where the top of a reef could be examined, the topography appear to be a primary attribute of the reef. Inter-reef deposits lap onto the reefs, are fine grained, dark and shaly, and are truncated by overlying beds. Initial reef deposits are skeletal wackestones containing some phylloid algae, brachiopods and echinoderm (crinoid) fragments. Increasing amounts of skeletal material occur upward into a skeletal, intraclastic packstone. Some of this packstone is phylloid-algal dominated, but a surprising abundance of micritic intraclasts are present (fig. 15). These intraclasts appear to be fragments of algal mats or mud layers disrupted either by burrowing or dessication and incorporated into the matrix by bioturbation.
Figure 14--Phylloid reefs in Ismay Zone, Hermosa Formation, Eight-mile Rapids, San Juan River Canyon, Utah (Paradox Group). Note relief of reef core and drape of flank beds, showing that reef had positive topography during growth. White line traces approximate depositional topography.

Figure 15--Photograph of intraclasts from the reef shown in figure 14. These intraclasts indicate subaerial exposure during reef growth or at least major disruption of algal mats or mud layers in shallow water. Two-pound hammer is scale.

At the top of the reef subaerial exposure is indicated by truncation, thin autoclastic breccia and, more rarely by dissolution features. A thin skeletal wackestone overlies the unconformity, succeeded upwards by a very thin black, shaly, Chaetetes-bearing limestone. This in turn grades upward into gray skeletal wackestone of the next cycle. Amounts of detritus and porosity are inversely related, porosity being greatest in upper units (enhanced by subaerial diagenesis) and fine-grained detritus being most abundant in the basal units.
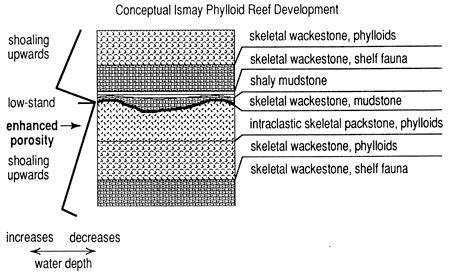
Relationship of sea level-rise rate changes to sedimentation in this reef development is reasonably clear. Initial transgressive deposits are the thin skeletal wackestone, covered by a condensed section of black shaly carbonate rarely containing Chaetetes. Overlying gray skeletal wackestone represents sedimentation during slowing rate of rise. Later colonization with phylloid algae represents vertical growth into the photic zone. The 'Overall reef deposit shapes suggest wave and current control of initial phylloid accumulations(fig. 16). Stillstand or relative lowering of sea level accounts for the subaerial exposure surface at the top of the reef.
Figure 16--Schematic diagram of Ismay reef evolution based on unpublished measured sections, San Juan River Canyon, Utah. Growth of the reef and its architecture is related to the rate at which sea-level rise occurred.
Porosity development appears to be related to subaerial exposure in this reef type as well as in the biodetrital reefs, and it may reflect both the lack of early cementation, bimodal algal plate and mud grain-size distribution, and the presence of mixed aragonite and calcite skeletal mineralogies.
Carbonate generation, transportation and degradation processes control the formation of reefs, coupled to physical settings and changes in the rate of sea-level rise. Resulting fabrics record these controlling parameters and permit the tracing of reef fabric development. Frequently porosity development is related to subaerial exposure, but early cementation is common in framework reefs and has tended to prevent secondary or later porosity enhancement.
Diagenesis of reefs and other shallow water carbonate deposits can be initiated by submarine cementation, as in the example of ancient framework reefs where adequate open space and water turnover occurred. For most other reefs, cessation of sea level rise or elevation of the reef and exposure to meteoric waters provide a significant early diagenetic paragenesis that can influence future petroleum trapping.
Although these concepts appear to be well-founded in observations, continued study is necessary to test the hypotheses and determine the validity of proposed models.
Adey, W. H., Macintyre, I. G., and Stuckenrath, R., 1977, Relict barrier reef system off St. Croix: Its implications with respect to late Cenozoic coral reef development in the western Atlantic: Proceedings of the Third International Coral Reef Symposium, Miami, p. 15-21.
Adey, W. H., 1975, The algal ridges and coral reefs of St. Croix: Their structure and Holocene development: Atoll Research Bulletin, n. 187, p. 1-67.
Adey, W. H., and Burke, R. B., 1976, Holocene bioherms (algal ridges and bank-barrier reefs) of the eastern Caribbean: Geological Society of America, Bulletin, v. 87, p. 95-109.
Adey, W. H., and Burke, R. B., 1977, Holocene bioherms of Lesser Antilles--Geologic control of development,: American Association of Petroleum Geologists, Studies in Geology, no. 4, p. 67-81.
Brinton, Lise, 1986, Deposition and diagenesis of Middle Pennsylvanian phylloid algal banks, Paradox Formation, Ismay Zone, Ismay field and San Juan Canyon, Paradox basin, Utah and Colorado: Masters thesis T-3101, Colorado School of Mines, Golden, Colorado, 315 p.
Davies, P.J. and Marshall, J.F., 1985, Halimeda bioherms--low energy reefs, northern Great Barrier Reef: Proc. Fifth International Coral Reef Congress, Tahiti, V. 5, p. 1-7.
Fagerstrom, J.A., 1987, The evolution of reef communities: John Wiley and Sons, New York, 600 p.
Gerhard, L.C., 1978, Reconnaissance study of sediment facies of Salt River estuary, St. Croix. U.S.V.I.: Mixed carbonates and clastics: The Mountain Geologist, v. 15, no. 3, p. 109-123.
Gerhard, L.C., 1981, Origin and evolution of Sand Cay (Candlelight Reef), U.S. Virgin Islands: Atoll Research Bulletin, No. 242, p. 1 -11, 11 pl.
Heckel, P. H., 1974, Carbonate buildup in the geologic record: a review; in, Reefs in time and space, L. F. Laporte, ed.: Soc. Economic Geologists and Paleontologists, Special Publication No. 18, p. 90-154.
Hine, A. C., Hallock, P., Harris, M. W., Mullins, H. T., Belknap, D. F., and Japp, W. C., 1988, Halimeda bioherms along an open seaway--Miskito Channel, Nicaraguan Rise, SW Caribbean Sea: Coral Reefs, v. 6, n. 3/4, p. 173-178.
Hubbard, D. K., Burke, R. P., and Gill, I. P., 1985, Accretion in deep, shelf-edge reefs of St. Croix, US VI; in, Deep-Water Carbonates, P.D. Crevello and P. M. Harris, eds.: Society of Economic Paleontologists and Mineralogists, Core Workshop 6, p. 491-527
James, N. P., and Macintyre, I.G., 1985, Reefs: zonation, depositional facies, diagenesis: Quarterly of the Colorado School of Mines, v. 80, no. 3, 70 p.
Longman, M. W., 1981, A process approach to recognizing facies of reef complexes; in, European Fossil Reef Models, D. M. Toomey, ed.: Society of Economic Mineralogists and Paleontologists, Special Publication 30, p. 9-40
Lowenstam, H. A., 1950, Niagaran reefs of the Great Lakes area: Journal of Geology, v. 58, p. 430-487
Macintyre, I. G., and Glynn, P. W., 1976, Evolution of modern Caribbean fringing reef--Galeta Point, Panama: American Association of Petroleum Geologists, Bulletin, v. 60, p. 1054-1072
Macintyre, I. G., Multer, H. G., Zankl, H. L., Hubbard, D. K., Weiss, M. P., and Stuckenrath, R., 1985, Growth and depositional facies of a windward reef complex (Nonesuch Bay, Antigua, W. I.): Proceedings of the Fifth International Coral Reef Congress, v. 6, p. 605-610
Marshall, J. F., and Davies, P. J., 1988, Halimeda bioherms of the northern Great Barrier Reef. Coral Reefs, v. 6, nos. 3/4, p. 139-148
Milliman, J. D., 1974, Marine carbonates: Springer-Verlag, Berlin, 375 p.
Moore, G. E., and Wilde, G. L., eds., 1986, Lower and Middle Guadalupian facies, stratigraphy, and reservoir geometries, San Andres/Grayburg Formations, Guadalupe Mountains, New Mexico and Texas: Permian basin Section, Society of Economic Paleontologists and Mineralogists Publication 86-25, 144 p.
Neumann, A. C., and Macintyre, I., 1985, Reef response to sea level rise--keep-up, catch-up or give-up: Proceedings of the Fifth International Coral Reef Congress, v. 3, p. 105-109
Orme, G. R., and Salama, M. S., 1988, Form and seismic stratigraphy of Halimeda banks in part of the northern Great Barrier Reef Province: Coral Reefs, v. 6, nos. 3/4, p. 131-138
Phipps, C. V. G., and Roberts, H. H., 1988, Seismic characteristics and accretion history of Halimeda bioherms on Kalukalukuang Bank, eastern Java Sea (Indonesia): Coral Reefs, v. 6, nos. 3/4, p. 149-160
Pray, L. C., 1987, Capitan reef complex (Permian), Guadalupe Mountains, southwestern United States--a classic sedimentologic model in flux (abs.): American Association of Petroleum Geologists, Bulletin, v. 71, p. 1,441-1,442
Reid, R.P., and Ginsburg, R. N., 1986, The role of framework in upper Triassic patch reefs in the Yukon (Canada): Palaios, v. 1, no. 6, p. 590-600.
Roberts, H. H., and Aharon, P., and Phipps, C. V., 1988, Morphology and sedimentology of Halimeda bioherms from the eastern Java Sea (Indonesia): Coral Reefs, v. 6, nos. 3/4, p. 161-172
Sloss, L. L., 1962, Stratigraphic models in exploration: American Association of Petroleum Geologists, Bulletin, v. 46, p. 1050-1057.
Shaver, R. H., and Sunderman, J. A., 1989, Silurian seascapes: Water depth, clinothems, reef geometry. and other motifs--A critical review of the Silurian reef model: Geological Society of America, Bulletin, v. 101, p. 939-951.
Textoris, D.A., and Carozzi, A. V., 1964, Petrography and evolution of Niagaran (Silurian) reefs, Indiana: American Association of Petroleum Geologists, Bulletin, v. 48, p. 397-426.
Toomey, D. F., 1970, An unhurried look at a Lower Ordovician mound horizon, Southern Franklin Mountains, West Texas: Journal of Sedimentary Petrology, v. 40, p. 1318-1334.
Zankl, H., and Shroeder, J. H., 1972, Interaction of genetic processes in Holocene reefs off North Eluthera Island, Bahamas: Geologische Rundschau, v. 61, p. 520-541
Kansas Geological Survey, Energy Research
Placed online April 21, 2010; original report from 1990
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/PRS/publication/1990/OFR90_11/index.html