Kansas Geological Survey, Open-file Report 96-35
Prev page--Start ||
Next page--Summary, References
Sites were selected to represent the loess-mantled upland, both proximal and distal to the Kansas River valley, loess and colluvium-mantled straths on the Kansas River valley wall, and late-Pleistocene terraces in the Kansas River valley. These landscape situations are diverse and represent different depositional and environmental regimes. All sites have been 14C dated to establish the chronostratigraphy (table 1), but two of the sites have been sampled for additional age control; the samples are currently pending.
Table 1--Radiocarbon (14C) Ages.
| Site Name (Core Number) | Depth (cm) | ISGS no.1 | δ13C (‰) | Corrected Age (yr Bp)2 |
|---|---|---|---|---|
| Sumner Hill (21)3 | 150-160/pit 6 | 3101 | -16.2 | 6,140±130 |
| 170-180/pit 6 | 3140 | -17.1 | 7,770±220 | |
| 200-210/pit 6 | 3142 | -16.8 | 11,230±310 | |
| 240-250/pit 6 | 3131 | -16.2 | 11,980±270 | |
| 260-270/pit 6 | 3143 | -15.9 | 13,240±310 | |
| 280-290/pit 6 | 3146 | -15.6 | 13,470±430 | |
| 215-225/pit 2 | 3164 | -15.5 | 19,170±650 | |
| 310-320/pit 5 | 3159 | -16.0 | 20,760±750 | |
| 255-265/pit 2 | 3160 | -15.4 | 21,580±740 | |
| 235-245/pit 2 | 3163 | -15.1 | 21,610±590 | |
| 330-340/pit 5 | 3158 | -17.0 | 22,560±990 | |
| 350-360/pit 5 | 3157 | -17.2 | 23,000±1,100 | |
| Bala Cemetery (19)4 | 150-220 | 2622 | -17.3 | 19,070±280 |
| Pump House Canyon5 | 130-140 | 3165 | -15.6 | 5,810±180 |
| 290-300 | 3167 | -21.6 | 18,830±450 | |
| 330-340 | 3166 | -18.4 | 23,010±850 | |
| Manhattan Airport (33) | 40-48 | 2996 | -14.4 | 2,080±70 |
| 163-168 | 2997 | -20.4 | 11,550±440 | |
| 525-528 | 3004 | -16.5 | 11,580±570 | |
| 228-233 | 3003 | -18.3 | 13,740±540 | |
| Manhattan Airport (20) | 168-240 | 2623 | -19.4 | 19,990±450 |
| No name (28) | 23-28 | 3005 | -15.5 | 1,000±100 |
| 28-33 | 3006 | -15.4 | 1,730±110 | |
| 147-152 | 3008 | -19.0 | 14,140±770 | |
| 202-207 | 3007 | -17.3 | 15,110±590 |
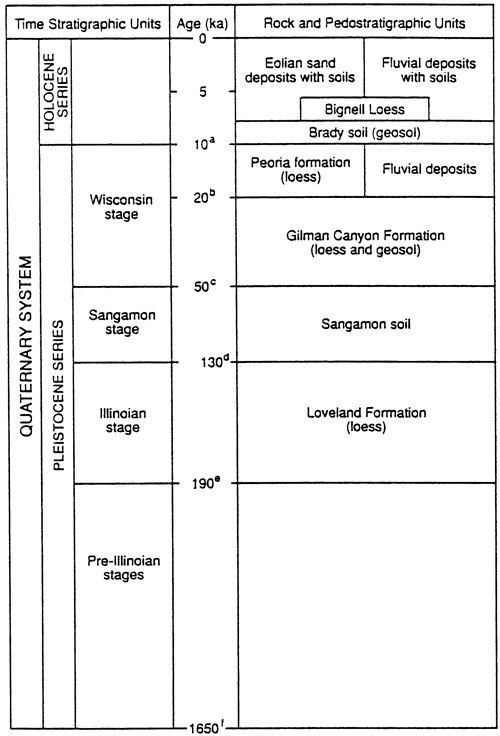
Figure 1--Late-Quaternary stratigraphic succession for the nonglaciated part of Kansas. Age designation rationale consists of the following: Pleistocene-Holocene boundary after Hopkins (1975); (b) the uppermost Gilman Canyon Formation 14C dates at about 20 ka (Johnson, 1993a, b); (c) the age of 50 ka is an estimate for the beginning of Gilman Canyon time, not the end of Sangamon soil development, i.e., the boundary is believed to be an unconformity; (d) 130 ka is consistent with the sea-level record and the age of marine isotope boundary 5e/6, the presumed end of the Illinoian glacial stage and with recent themoluminescence age determinations in south-central Nebraska and central Kansas (Maat and Johnson, 1996; Johnson and Muhs, 1996); (e) age of the marine isotope boundary 6/7 as determined by Martinson and others (1987); and (f) age of the Pliocene-Pleistocene boundary at the Virca, Italy section (Aguirre and Pasini, 1985).
Results from the extraction and analysis were unanticipated, based upon extensive experience in the extraction and analysis of phytoliths from loess deposits elsewhere in the region. Both concentration and preservation of phytoliths were poor, except for the upper part of the core. From 0 to 40 cm phytolith concentration and preservation were good, and from 40 to 60 cm concentrations and preservation were adequate for interpretation. From 60 to 350 cm, phytolith preservation and concentration were marginally adequate for analysis. Extraction procedures were repeatedly verified and samples rerun in order to check the concentrations. The effectiveness of the extraction procedure was attested to by good recovery of Lycopodium spike grains. Poor preservation typically associates with low concentrations. Further, phytoliths recovered frequently exhibited deep solutional pitting.
The degree of phytolith preservation correlates well with the pH stratigraphy of the core: Within the upper part of the core where preservation is good, pH values are about 5.9 to 7, but below that pH rises to a precarious level for the preservation of phytoliths (Pease, 1967). Other locations where phytolith analyses have been conducted on loess, pH, where measured, has been generally between 7 and 8. Although high pH levels may not be exclusively responsible for the poor preservation, they certainly seem to be a contributing factor given the close correlation between pH and preservation.
All isolates were studied microscopically, but, because of concentration and preservation, every sample was quantitatively analyzed in the upper 350 cm and every other sample below that level. Results are presented in two formats: relative frequencies of all phytolith types encountered (fig. 2) and relative frequencies of grass short cells alone (fig. 3). The latter presentation was produced so that the presence of the other phytolith types would not obscure the interpretation of the phytoliths diagnostic of the various grass subfamilies.
The percentage diagrams (figs. 2, 3) have been subdivided into three zones: A, B, and C. Zone C represents the bulk of the core in which phytolith data are not interpretable, i.e., Gilman Canyon and earlier material. The break between Zones B and C coincides with the sedimentary, pedogenic, and isotopic break at about 358 cm. With such small short cell sums, the percentage bars are inflated and largely meaningless. Zone B, extending from the late Pleistocene to the late Holocene, exhibits a chloridoid peak centered on about 130 cm, or about 5 ka. At about 275-350 cm (14-13 ka), an increase in C4 panicoid types occurs, showing a change in the grass composition. Arboreal types appear within the zone, particularly about 11 ka and earlier; coniferous types (e.g., Picea and Pinus) are likely at least part of this arboreal vegetation based upon macro- and other microbotanical information from the late Pleistocene record of the region. Zone A represents the upper part of the core which is dominated by the surface soil development and late Holocene sedimentation. C4-type grasses dominate: panicoid and chloridoid subfamilies are high relative to the C3 pooids.
Figure 2--Frequency of Phytolith Types from the Sumner Hill Site.
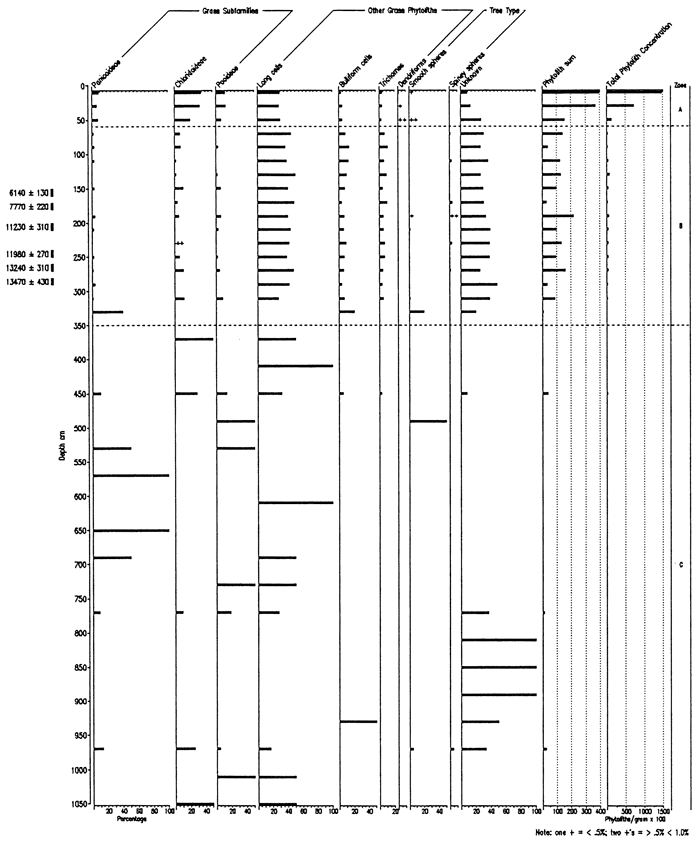
Figure 3--Frequency of Grass Short Cells and Arboreal Type Phytoliths from the Sumner Hill Site.

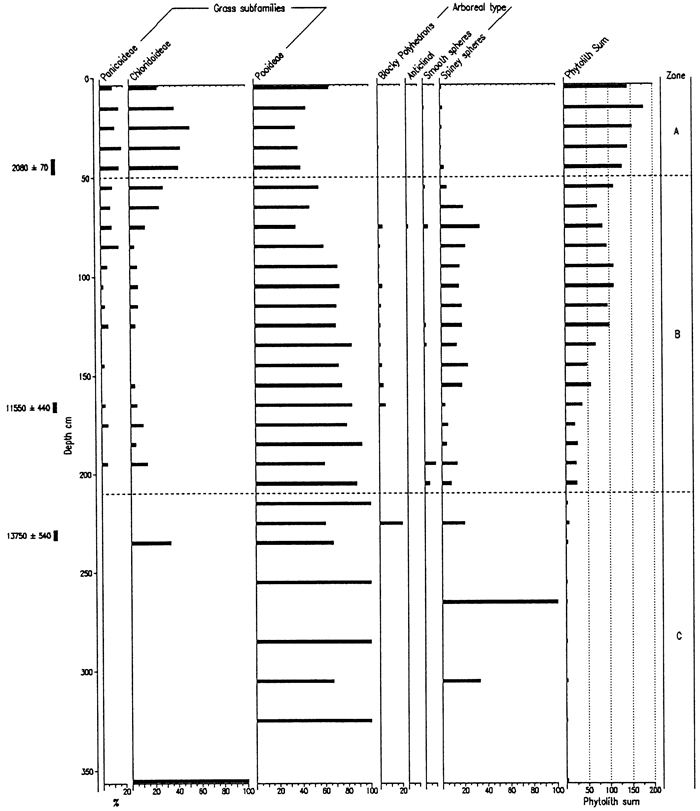
Biosilicate recovery from the Bala Cemetery site was very good to within the lowermost 40cm of the core, and the percentage of unknown phytoliths is relatively small (figs. 4, 5). Zone F (>240 cm depth) is uninterpretable and consists of the lower Gilman Canyon Formation and limestone residuum. Zone E, 240-170 cm, is distinguished by the relatively high frequencies of spiney spheres, the most common arboreal-type phytolith and indicative of a moist environment. Moreover, the predominance of pooid phytoliths suggests a cool, moist climate during this time. In Zone D, pooid types are at their highest percentage for the entire core, probably a reflection of the glacial maximum climate. There is, however, little evidence of trees during this time. The arboreal component appears to expand within Zone C, while the pooids decrease upward and chloridoids increase, i.e., a shift toward warmer, drier conditions. Zone B features a maxima in pooids, indicating decreased temperatures and a likely increase in available moisture. The uppermost zone (A), coinciding with the modern surface soil, exhibits a decrease in pooids and increase in the chloridoids and panicoids, i.e., warmer temperatures. The change from Zone B to A may be a function of either climate shift or historical cropping (e.g., sorgum). Age data for the site are yet pending and will provide the necessary time control for the interpretation and correlation with other sites.
Figure 4--Frequency of Phytolith Types and Sponge Spicules from the Bola Cemetery Site.
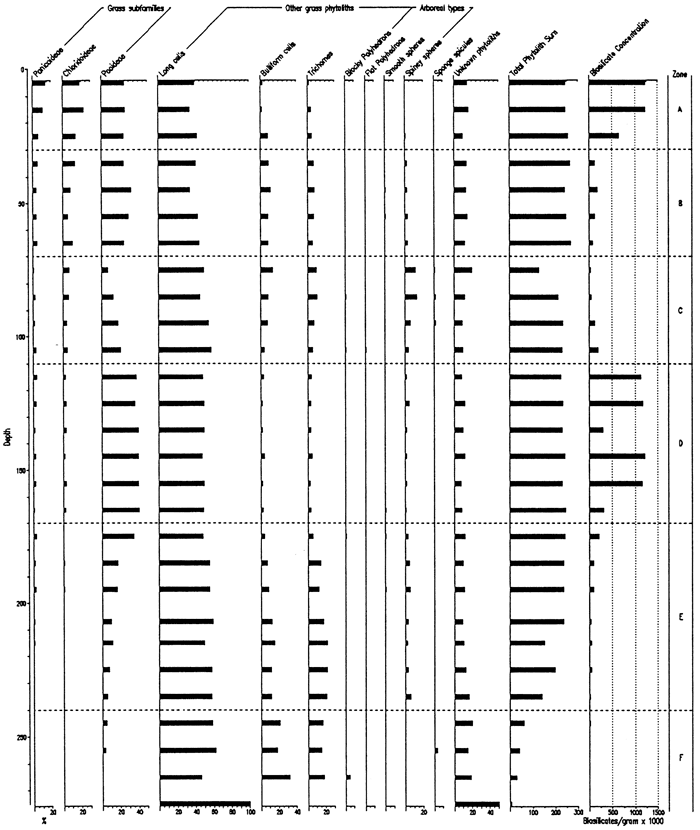
Figure 5--Frequency of Grass Short Cells and Arboreal Type Phytoliths from the Bala Cemetery Site.
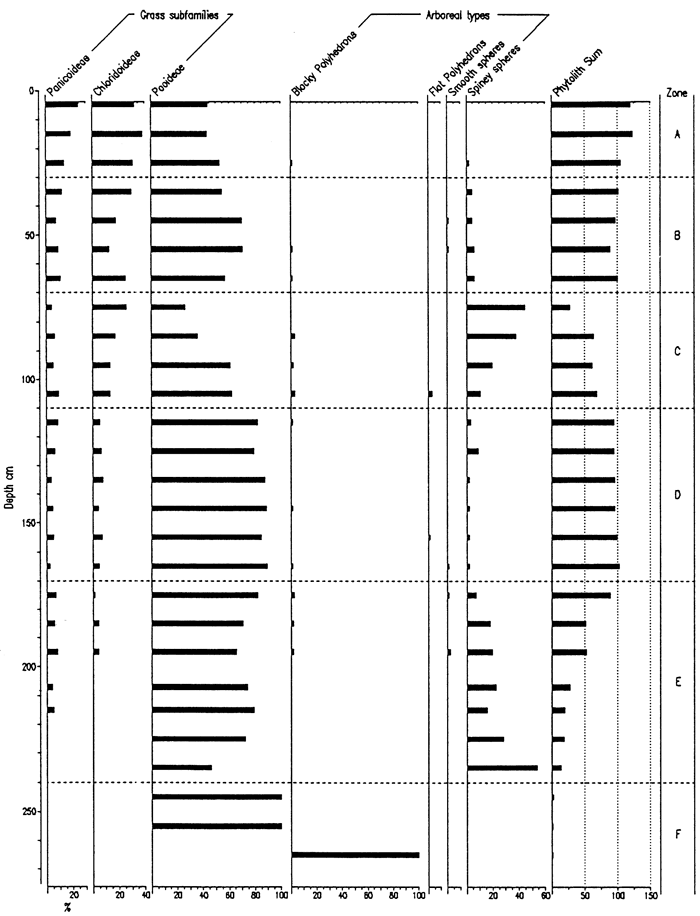
Phytolith preservation was extremely poor in samples collected at the Pump House Canyon site, except in the uppermost 40 cm (figs. 6, 7). Despite poor preservation, four zones are definable. Zone D (>90 cm) is uninterpretable due to the low concentrations of biosilicates, a likely result of corrosion due to elevated pH values. Zone C (70-90 cm) is defined by high frequencies of spiney spheres, an arboreal type. Similar ratios of spiney spheres and pooid short cells were recognized at the same depths (75 and 85 cm) in the Bala Cemetery core, indicating a similar type of vegetation at both sites. Zone B (40-70 cm) is characterized by a decrease in chloridoid types and increase in pooids, with a diversity of arboreal types. The change in frequencies of the three types of grass phytoliths parallels that for the Bala Cemetery site. Pooids dominate Zone A, although chloridoids are well represented; panicoid types play a minor role throughout.
Figure 6--Frequency of Phytolith Types and Sponge Spicules from the Pump House Creek Site.

Figure 7--Frequency of Grass Short Cells and Arboreal Type Phytoliths from the Pump House Creek Site.
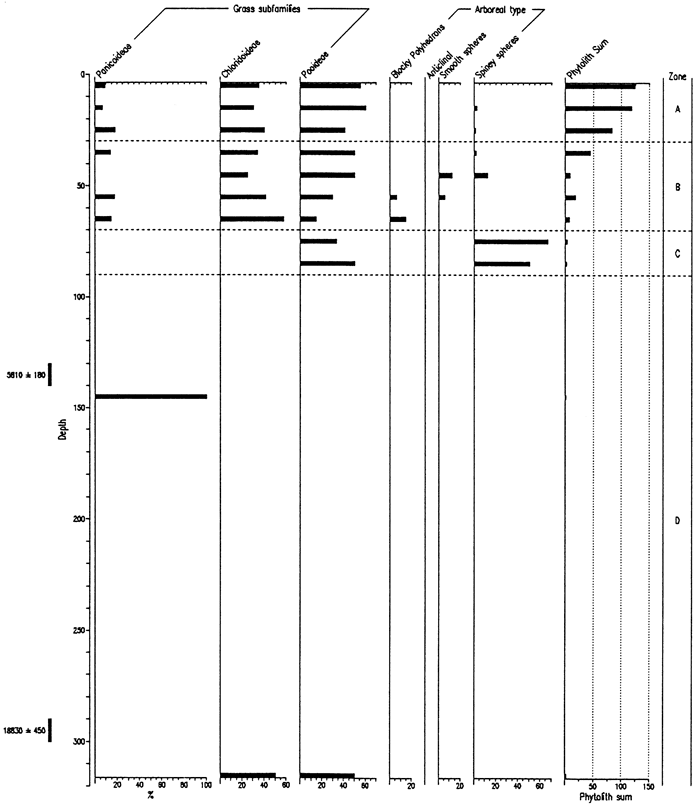
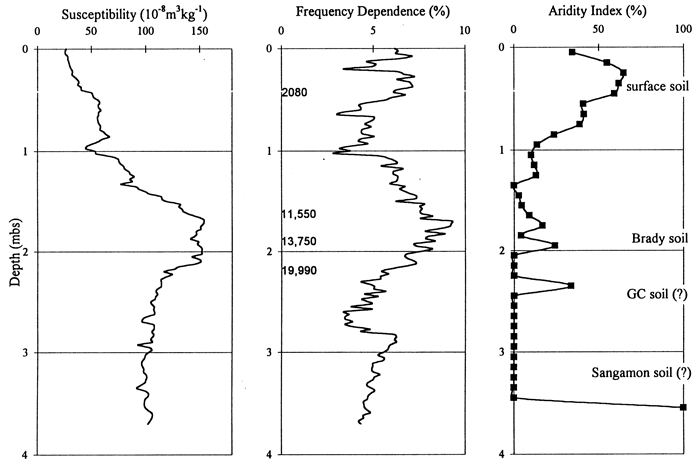
On the basis of samples analyzed from the Manhattan Airport backhoe trench, the diagrams (figs. 8, 9) have been subdivided into three zones. The lowermost zone, C, has been differentiated on the basis of poor preservation and low concentration of phytoliths. Consequently, variations in phytolith values before about 13 ka are meaningless and uninterpretable. Zone B, or the latest Pleistocene and Holocene, exhibits good phytolith preservation. The pooids are dominant, but decline in the upper part of the zone, i.e., as the Holocene progressed; chloridoids increased, but panicoids showed little change. Significant numbers of arboreal-type phytoliths, particularly spiney spheres, are present throughout this zone. Zone A, coincident with the surface soil and encompassing about the last 2,000 years, indicates a maximum in chloridoid grasses and a decrease in the arboreal component from Zone B.
Figure 8--Frequency of Phytolith Types and Sponge Spicules from the Manhattan Airport Site.

Figure 9--Frequencies of Grass Short Cells and Arboreal Type Phytoliths from the Manhattan Airport Site.
A vision of how the upland landscape of Fort Riley evolved during the last 120,000 years or so is now beginning to emerge. Such a perspective provide a useful context for viewing environmental reconstructions. Evolution of the uplands is, of course, not a process isolated from the valley environment; the alluvial setting is extremely sensitive to environmental change and intimately linked to the upland change. Reconstructions of the alluvial history is presently underway and will likely provide data of greater temporal resolution than that of the uplands, at least for the Holocene.
During pre-Illinioan time, the ancestral Kansas River was cutting a strath in the limestones of the present valley walls, tens of meters above its present elevation. This bedrock surface and it covering of alluvial sands and gravels were subsequently mantled with Loveland loess, at a time when the river had entrenched to near its present level. Loess fall was relatively limited, which indicates that the source was either the alluvial plains to the northwest or the Missouri River valley to the east; either source is sufficiently removed spatially or meteorologically such that little loess fall would occur in this distal location. At the end of Illinoian time, the Sangamon soil developed in this thin mantle of loess. The alluvial phase of the Sangamon soil observed at the Manhattan airport site indicates the river was certainly no higher than this high terrace during the last interglacial.
From the ubiquity of the Sangamon soil in the study area as well as throughout the region, it is apparent that erosion was limited following Sangamon time, i.e., during the early Wisconsinan glacial stage. The Sangamon soil appeared to have survived and continued to dominate the landscape until the middle Wisconsin. At this time another loess unit was being deposited, i.e., the Gilman Canyon loess. The source of this loess is unknown, but it appears to different than that of the Loveland loess because it maintains a uniform thickness throughout the central Great Plains, rather than exhibiting a distance decay function as with the Loveland and Peoria loesses. Gilman Canyon loess fall was, however, exceedingly slow: the meter or so of loess accumulated over approximately 20,000 years, i.e., from before 40 ka to about 20 ka. Other than the relatively thin nature of the loess unit, the occurrence of relatively low depositional rates (surface stability) is indicated by the presence of two or more periods of soil development. The pedogenesis occurred primarily during the latter part of Gilman Canyon time (Farmdalian substage?) in that soil development typically dominates the upper half of the formation.
The contact between the Gilman Canyon Formation and the overlying Peoria loess is transitional. It appears that Gilman Canyon pedogenesis waned as the late-Wisconsinan loess began to fall such that accumulation rates eventually exceeded the rate of pedogenesis. Peoria loess deposition was, like the Loveland loess, relatively low in the study area. The loess unit thickens to the west and northwest in proximity to the sources, e.g., the Sand Hills of Nebraska, Platte River valley, and to a lesser extent adjacent to the other large river valleys crossing the central Great Plains, e.g., the Republican and Kansas. Accordingly, Peoria loess is thickest in the northwestern part of the reservation; over 2 m appear to be present at the Bala Cemetery site.
Although no direct evidence has yet been found, some of the Peoria loess, as with other loess units, was probably lost, at least locally, during erosional maturation of the late-Wisconsinan landscape. Clearly, sites to the northwest such as Bala Cemetery have realized erosion of the Peoria loess and overlying Brady soil, assuming it developed in the area. At some localities, such as Sumner Hill, the total thickness of Peoria loess present at the end of the Pleistocene was completely involved in Brady soil development.
The next major pedogenic period (after the Gilman Canyon) occurred during the Pleistocene/ Holocene transition. The Brady soil, developed in the upper Peoria loess as deposition ceased, is, on the basis of excavations and coring, well represented in the southern part of the reservation. Consequently, the upland surface was largely stabilized during this time of environmental shift from the Wisconsinan glacial stage of the Pleistocene into the Holocene. Deposition of the Bignell loess, beginning during the early Holocene, buried the Brady soil. The southerly wind flow off the Kansas River valley apparently transported silts and fine sands from the alluvial bottom up onto the uplands. The rapid distance decay factor is suggested by the lack of Bignell loess at the Bala Cemetery site to the northwest of the Sumner Hill and Pump House Canyon sites. The contrast may be due to differential erosion, but this is deemed unlikely given that the two sites to the south adjacent to the Kansas River valley are in equally or more erosion-prone settings. Since the Brady soil is presumed to have developed to the northwest but is now apparently gone (e.g., Bala Cemetery), a thin layer of the Bignell loess was probably present but quickly eroded; now the Peoria loess is now exposed with its associated surface soil.
Throughout the reservation, surface soil development appears relatively recent, probably within the last 2,000 to 1,000 years and even less locally. This relative surface youth indicates that either erosion has occurred recently or that Bignell loess has been episodically or slowly accumulating on the landscape until relatively recently. The latter scenario appears correct, at least for areas along the southern margin of the reservation adjacent to the Kansas River valley, in that the Holocene upland surface has been periodically stable (slow or zero loess accumulation) such that soil formation could occur. These short periods of stability are manifest as weak or incipient soils developed in the Bignell loess, such as those indicated at the Pump House Canyon site. The preservation of cultural sequences, ranging from Clovis to Woodland and Protohistoric, beginning in the Brady soil and extending up through the Bignell loess into the surface soil at archaeological sites in these setting within the region (e.g., Johnson, 1996a) also indicates that there has been little erosion of the Bignell loess, at least locally.
Given the preservation and distribution of late-Quaternary deposits on the uplands, Fort Riley is an ideal study area for modeling landscape evolution in the context of cultural history. Further, the loess and alluvial deposits of the reservation permit ready extraction and interpretation of proxy data for the reconstruction of past environments.
Opal phytolith analysis of prehistoric sediments was first done in the Great Plains about ten years ago, but only a handful of studies have been reported; only 17 studies have been done on a total of 15 sites in Nebraska, Kansas, Oklahoma and Texas. Of these studies, nine were seeking archaeological information (e.g., cultigen use), and seven for environmental reconstruction.
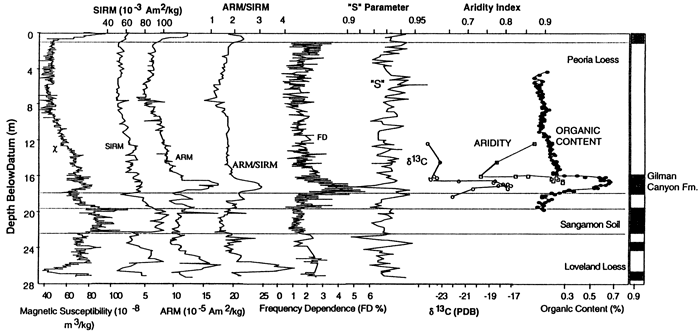
Of the five sites investigated in Nebraska, the Eustis ash pit is the most notable. Mining of volcanic ash up into the 1950s resulted in a 35 m-high vertical face exposing loess and intercalated soils. Analysis of 32 samples collected from throughout the section demonstrated that phytoliths are relatively concentrated within the loess at that locality and that they are abundant in both the buried soils and in the intervening loess units (Fredlund et al., 1985). Results indicated that the soils formed during periods that were relatively warmer and drier than the periods of major loess deposition, including the modern surface soil. Two subsequent studies on samples from the site further articulated the utility of phytolith analysis in environmental reconstruction. An analysis by Bozarth of samples collected at close interval from the Gilman Canyon Formation and adjacent lower Peoria loess indicated a cool, moist grassland during early Gilman Canyon time which shifted to a warmer, drier grassland as soil development occurred and back to cool, more moist conditions again when the Peoria loess began to accumulate (Johnson et al., 1993a). A very good correlation exits between the δ13C data and the aridity index computed from the phytolith data for the Gilman Canyon Formation; six magnetic parameters reflect the unique climatic episode as well through the climatic controls on weathering (fig. 10). In a novel approach, Fredlund (1993) extracted O, H, and C isotopic data from individual phytoliths to test the feasibility doing such to reconstruct past environments; in this pilot study, each of the samples analyzed produced results approximating the climatic conditions of the respective period.
Figure 10--Stratigraphic plots of magnetic parameters (χ, SIRM, ARM, ARM/SIRM, FD, and S) compared with stratigraphic plots of δ13C values derived from bulk organic carbon, an aridity index derived from opal phytolith data, and percent organic matter, all from the Eustis ash pit in southwestern Nebraska (Johnson, 1996b).
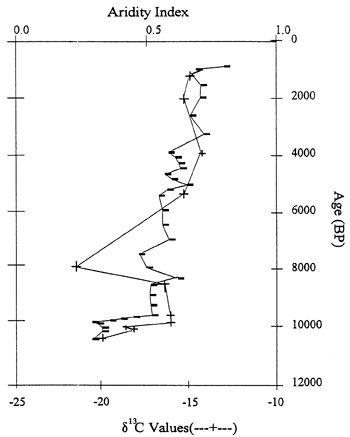
The Sargent site, also in southwestern Nebraska, consists of a 6m-high vertical exposure within a gully cut into the loess-mantled upland along the south wall of the Platte River valley. Exposed is late Pleistocene Peoria loess overlain by Holocene Bignell loess containing at least five buried soils. Fossil biosilicates (phytoliths, sponge spicules, statospheres) were well preserved in all 34 samples analyzed (fig. 11; Bozarth, 1992b). The phytolith record indicates that a deciduous forest or open woodland was growing at the site about 10,640 years B.P. By about 10,080 years B.P., the site had been transformed into a pooid-dominated grassland, reflecting a relatively cool and moist growing season climate. After 10,000 years B.P., there was a relatively rapid increase in chloridoid grasses, reflecting increasing temperatures and reduced available moisture, i.e., appearance of the Holocene climatic regime. Conditions continued to become drier and warmer until about 3,800 years B.P., when a slight increase in available moisture occurred as indicated by the rise in panicoid phytoliths. The present dominance of chloridoid-type phytoliths reflects the situation of the site in the ecotone separating grama grass-buffalo grass from needle grass-wheat grass. A plot of the aridity index (after Diekmeyer, 1994) against δ13C data from the site reveals a close correspondence between the two data sets and defines the increasing aridity throughout the Holocene (fig. 12).
Figure 11--Frequency of Occurrence (%) of Phytoliths at the Sargent Site, Southwestern Nebraska (Bozarth 1992b).
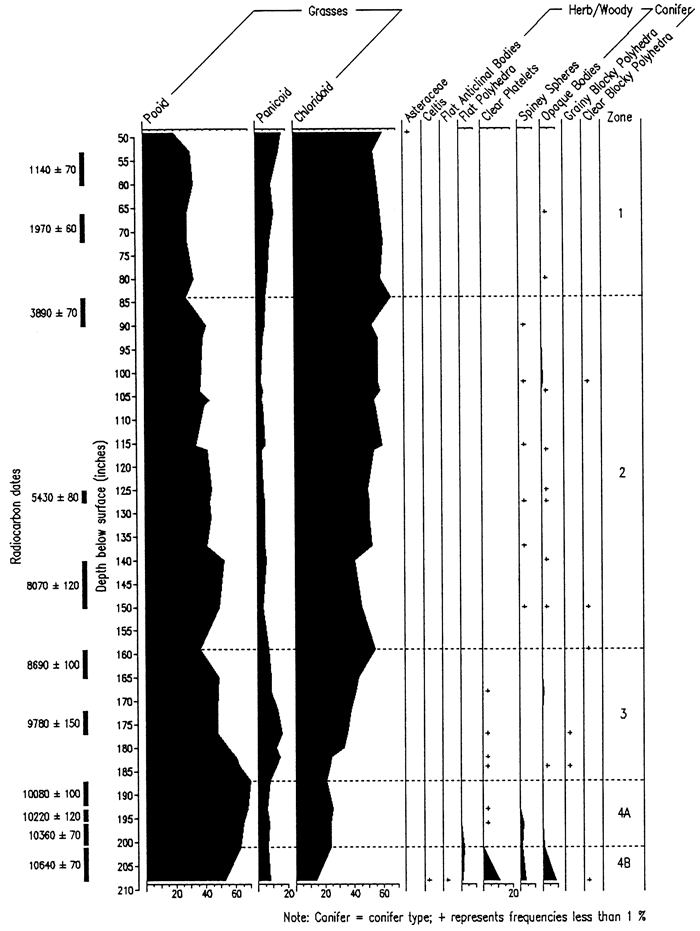
Figure 12--Aridity index (after Diekmeyer, 1994) and δ13C data curves for the Sargent site, southwestern Nebraska (Bozarth, 1992b; Dort, 1996).
The La Sena site is a Paleoindian archaeological site buried in a loess-mantled terrace in southwestern Nebraska and was exposed by wave action on Harry Truman Reservoir (May, 1991; May and Holen, 1993). The cultural deposit, consisting of a bed of fractured mammoth bones, appears to have been a butchering site at about 18,000 years B.P. Well-preserved phytoliths were isolated from four sediment samples. The dominance of pooid phytoliths in the mammoth bone bed, compared to a modern analog, indicated that the climate was much cooler and more moist than at present. Moreover, the presence of non-grass phytoliths suggested that deciduous trees were growing on or near the site at that time (Bozarth, 1991b).
Two disparate areas in Nebraska were investigated for the presence of cultigen phytoliths. Numerous well-preserved phytoliths were isolated from samples collected at three archaeological sites on Harlan County Lake in south-central Nebraska. No phytoliths derived from cultigens were found, but those grass phytoliths detected indicated an environment similar to that of today (Bozarth, 1987d). At archaeological site 23DX3, a Central Plains Tradition village discovered in northeastern Nebraska, Bozarth (1984b) identified silicified sunflower-like, multi-celled hair bases in phytolith isolates from sediment samples collected in cultural features.
Eight sites have been studied in Kansas, four environmental reconstructions and four archaeological investigations. Kurmann (1981, 1985) extracted phytoliths, as well as pollen and fungal spores from both a modern soil and a buried soil in alluvial fill of the Flint Hills region of east-central Kansas with the intent of obtaining a correlation with local vegetation. Pollen could distinguish woodland from prairie but was unable to differentiate short-grass from tall-grass prairie, but the phytolith study did so. In another environmental study, Bozarth (1986) collected nine samples between 20 and 213 cm depth from within a cutbank of the Saline River on the upper reaches of Wilson Lake, central Kansas in order to characterize the environment at the time of alluvial deposition. Well-preserved phytoliths were isolated from all samples, including a modern analog. A comparison of the fossil phytoliths with the modern assemblage indicated that a period of alluviation after 5090 years B.P. and before 1940 years B.P. was drier than today, whereas the period of soil formation about 1940 years B.P. was more moist than at present. A comparison of the relative frequencies of chloridoid, panicoid, and pooid types indicates that the vegetation at the Wilson Lake area has been a mixed-grass prairie since that time.
In an attempt to reconstruct the diet of prehistoric bison of the central Great Plains, phytoliths were extracted from bison tooth calculus and impacta collected from a specimen found in Finney County, southwestern Kansas (Bozarth, 1993b). The phytolith assemblages derived from the calculus and impacta sample isolates indicated that the animal ate mostly short grasses (chloridoids) such as buffalo grass and blue grama (Bouteloua gracilis), but the presence of panicoid and pooid types provided evidence that the animal also grazed to some extent on the C4 tall grasses and the C3 cool-season grasses. The Jetmore mammoth site (14H01), located in western Kansas, consisted of a buried mammoth bone deposit that dated to 9,235 years B.P. (Bozarth, 1984b). Analysis of phytoliths isolated from four subsurface samples and one modern (surface) analog sample indicated that most of the grasses growing concurrently with the mammoth(s) were panicoids, suggesting that the area was more humid than at present. Phytolith data also indicated that deciduous trees were growing at the locality during the early Holocene. Fossil pollen analysis conducted on the same samples suggested an open woodland environment.
Phytoliths were isolated from four sediment samples collected in two archaeological sites at Perry Lake, northeastern Kansas with the goal of documenting the aboriginal use of cultigens (Bozarth, 1990a). No economic species were identified from the phytolith record, but pooid, chloridoid, and panicoid grasses were common at both sites. Phytoliths from dicots, likely deciduous trees, were also present in small concentrations. Site 14MN328, representing a Great Bend Aspect village in central Kansas, was occupied by the late prehistoric Wichita, a group known for extensive cultivation some 500 years ago. Bozarth (1989, 1990c, 1993a) isolated phytoliths from samples collected in seven cultural features, with the intent of searching for the presence of cultigens. Three bean (Phaseolus) phytoliths were recovered from a sample representing a trash-filled storage pit, and a maize cob phytolith was identified from a hearth sample. The Hatcher site (14D019) consists of the remains of a prehistoric habitation structure (Plains Village period) located adjacent to Clinton Lake in northeastern Kansas. Analysis of phytoliths isolated from archaeological and modern analog samples indicated that the site was covered with tall-grass prairie at the time of occupation. An associated examination of phytoliths contained within a daub concentration demonstrated that panicoid and pooid grasses were the most common grasses used in construction at the site (Bozarth, 1987b).
Another archaeological site studied, the Stigenwalt site (14LT351) in southeastern Kansas, revealed what has become known as the Stigenwalt complex, an early Archaic manifestation that occurred over a fourteen-hundred year period beginning about 8,810 years B.P. Analysis of well-preserved phytoliths from the site suggested that the primary vegetation community of the area at the time of occupation was one of mixed-grass prairie similar to that found to the west in central Kansas today (Bozarth, 1990b). This is a relatively dry type of grassland, significantly different from the more mesic mix of tall-grass prairie and cross-timbers forest common to the region at the present.
A single archaeological site in Oklahoma has undergone phytolith analysis: the Coody Creek site (34MS31) is a late Caddo village located in the eastern part of the state. Phytoliths were isolated from nine samples collected within four features as part of a paleoenthnobotanical study (Bozarth, 1984b). All samples yielded numerous well-preserved phytoliths, including squash-like groups of silicified epidermal cells
In a study of late-Quaternary alluvial fills in the Texas Panhandle, samples from eight sites were processed for their biosilicate contents, including phytoliths, siliceous bodies, and sponge spicules (Bozarth, 1995). Preservation was relatively good in samples from five of the sites: relatively high frequencies of chloridoid type phytoliths indicated that short-grass prairie was the dominant vegetation at Mustang Springs from about 10,000 years B.P. to the present, at the Edmonson site about 8,000 years B.P., at the Lubbock, Texas landfill about 3,100 years B.P., at the Sundown site about 2,100 year B.P., and at the Flagg site about 770 years B.P. Collectively, these findings suggest that short-grass prairie covered most of the Southern High Plains throughout the Holocene.
To date, phytoliths from very few sites have been analyzed, and most of those that have provide only a limited vignette of the past. Only the Sargent site in southwestern Nebraska provides a relatively detailed view of the Holocene; it is, however, much farther north than Fort Riley and therefore did not likely experience the extremes in moisture stress during the late Pleistocene and Holocene found to the south into Kansas. Fort Riley offers an opal phytolith record that complements the SIRA data and magnetic records (Johnson et al., 1994; Johnson, 1996b).
Whereas the resulting diagrams depicting the phytolith data were characterized above ("Results"), the following provides an interpretation in the context of the preceding discussion of past environments for the central Great Plains and the Fort Riley area.
The Sumner Hill site provides a viable phytolith record of the latest Pleistocene and Holocene, although variance is relatively large, due in part to poor preservation and low concentrations (fig. 2, 3). During the latest Pleistocene, about 14 to 11 ka, all three grass subfamilies seem to have been represented in a mixed grassland environment on this bluff-top landscape position. At least two samples from this interval indicate a strong panicoid grass representation, i.e., warm, but moist grassland climate. After about 11 ka, the panicoid grasses stayed about the same, but the warm, dry-adapted C4 grasses increased in percentage and the C3 grasses slowly declined, an expression of the progressively increasing temperatures as the Holocene grassland climate was being defined. The aridity index (fig. 13) exhibits a fluctuating curve due to the variation in the percentages of the phytolith types. Not all of this is likely due to low phytolith counts; some is certainly a function of climatic variability during the Holocene. The overall pattern is, however, a decrease in the cool-season C3 grasses and an increase in the warm-season C4 grasses, particularly the chloridoid species, from the late Pleistocene to the present. Magnetic data display little correlation with the aridity index, an indication that the detail of the phytolith record is largely due to low concentration and preservation (fig. 13).
Figure 13--Magnetic susceptibility and frequency dependence and aridity index data curves for the Sumner Hill site.
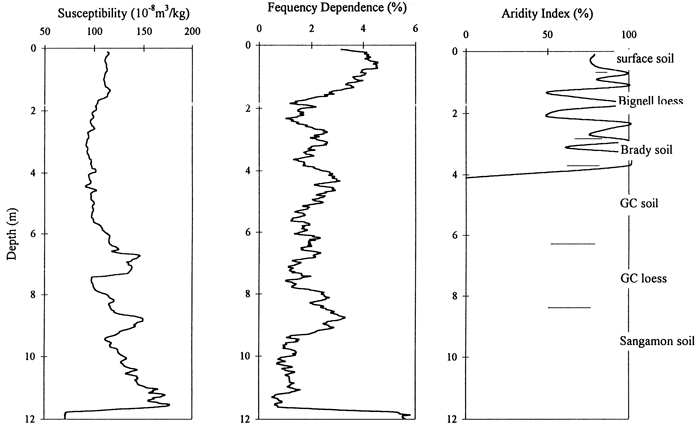
The Bala Cemetery site provides a quality phytolith reconstruction of the grassland environment on the broad uplands in the northeastern part of the reservation during the late Pleistocene (fig. 4, 5). C3 grasses are well represented throughout the sedimentary record, reflecting cool growing season temperatures and relatively low moisture stress during loess deposition. Temperatures were also too low for the C4 panicoid grass to successfully compete for a prominent role. Climate of the Last Glacial Maximum (LGM) is apparent in the pooid subfamily maximum (Zone D), which probably dates from about 18 to 16 ka. Spiney sphere concentrations indicate that deciduous trees were represented prior to and subsequent to the LGM; during the LGM coniferous trees may have replaced the deciduous varieties, but are not represented or recognized in the phytolith record. The drop in C3 grasses occurring at the top of Zone 3 is marked by a slight increase in the C4 chloridoid grasses; this may be an indication of a short-term perturbation in climate. The upper 50 cm of the profile indicates a slight warming as both types of C4 grasses increase. The rise in the panicoid C4 species indicates that moisture levels are staying relatively high, i.e., the main change is an increase in temperature. The decline in available moisture is pronounced in the Holocene, which is unrepresented here. The aridity index (fig. 14) depicts the gradual increase in moisture stress as the Pleistocene ends and the Holocene approaches. The brief drying at the top of Zone C is represented by the jump in the curve at about 70cm depth. The frequency dependence curve approximates the aridity curve, with the magnetic increase at about 1.5 m possibly relating to soil development during the LGM (fig. 14).
Figure 14--Magnetic susceptibility and frequency dependence and aridity index data curves for the Bala Cemetery site.
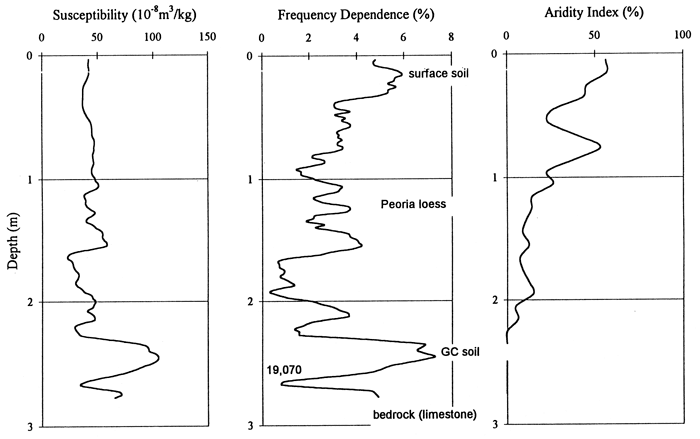
The phytolith record at the Pump House Canyon site is abbreviated, encompassing only the last 2-3,000 years, based on the single Holocene age of 5,810 years B.P. (fig. 6, 7). For a brief period, possibly several hundred years, the strath was occupied by C3 grasses and deciduous trees. A probable interpretation of the relatively high pooid grass population and trees is that the strath environment has a microclimate slightly less severe than the exposed upland sites above. Shortly thereafter the C3 grasses declined and the xeric C4 grasses dominated. This was a short-lived period: xeric C4 grasses decreased slightly and then varied little to the present; and C3 grasses expanded their role, with little subsequent change. The curve of the aridity index (fig. 15), restricted to the upper 90 cm, illustrates two dry peaks, which may correlate with soil formation identified in the frequency dependence curve.
Figure 15--Magnetic susceptibility and frequency dependence and aridity index data curves for the Pump House Canyon site.
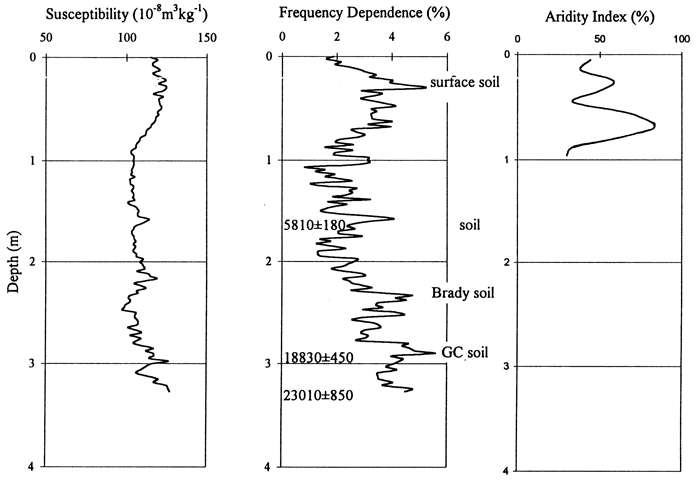
Phytolith stratigraphy from the Manhattan Airport site provides insight to conditions on the flood plain and terraces of the late Pleistocene and Holocene. The viable phytolith record begins about 13 ka. Cool-season grasses dominated the alluvial surface at this site from the Pleistocene-Holocene transition until the middle Holocene; these C3 grasses may, however, be edaphic, i.e., adapted to wet flood plain conditions, and not represent the broad scale climatic patterns. Deciduous arboreal species were also a major component during this time. Sometime prior to 2,080 years B.P. the C3 grasses declined and both xeric and mesic C4 grasses increased; the arboreal component also declined dramatically. Without higher-resolution age control, the timing of this change can only be estimated, but ages of 5,000 or 3,500 years B.P. are reasonable guesses given the peak of alluvial activity evident in the stratigraphic record at this time (Johnson and Martin, 1987; Johnson and Logan, 1990; Mandel, 1995; Johnson et al., 1996). This latter change may again be at least in part a response to changing local conditions, e.g., lowering water table with channel entrenchment, rather than to only regional climate shift. Nonetheless, the change is nicely documented and would impact aboriginal activity on this particular alluvial surface. The aridity curve (fig. 16), valid for only the upper 2 m, illustrates the amelioration in climate following the Pleistocene-Holocene transition and corresponds well with the magnetic data curves.
Figure 16--Magnetic susceptibility and frequency dependence and aridity index data curves for the Manhattan Airport site.
Prev page--Start || Next page--Summary, References
Kansas Geological Survey, Geology
Placed online Aug. 10, 2012
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/OFR/1996/OFR96_35/page2
.html