Kansas Geological Survey, Subsurface Geology 12, p. 61-63
by
Roger L. Kaesler
The University of Kansas
Since the mid-1960's, when high-speed, digital computers enabled sedimentary geologists to begin thinking about modeling, a concurrent but more or less unrelated revitalization of paleontology has produced significant advances in our understanding of fossils and the ancient communities of which they were a part. Among other effects of the revitalization has been an increased interest in paleobiology, the biology of ancient organisms. Now fewer paleontologists than in the past study biostratigraphy, and paleontologists new to the profession are likely to concern themselves with paleoecology, taphonomy, evolution, or extinction.
Paleontology has advanced on a broad front in the past two and a half decades. Such topics as taphonomy, time averaging, temporal resolution, species richness and diversity, constructional morphology, punctuated equilibrium, community evolution, competition theory, and extinction, now a part of every paleontologist's tool kit, had scarcely been thought of when the era of modeling began (fig. 1A). Because of this emphasis on paleobiology, however, advances in paleontology that are directly applicable to sedimentary modeling are sometimes not readily available to sedimentologists, especially those interested primarily in modeling physical processes. My goal, emphasizing ancient communities, is to summarize some of the aspects of paleontology that can be incorporated into sedimentary models. I set this goal in the belief that sedimentary models and discussions of sedimentology or stratigraphy are incomplete unless they incorporate information from paleontology.
Figure 1--A) Paleontological subjects that impinge on modeling of sedimentology and stratigraphy; B) competition and other interaction in biological communities; C) short-term and long-term changes in paleocommunities; D) factors that complicate our understanding of changes in paleocommunities-taphonomy, time averaging, and temporal resolution; E) aspects of community evolution; and F) questions about extinction.
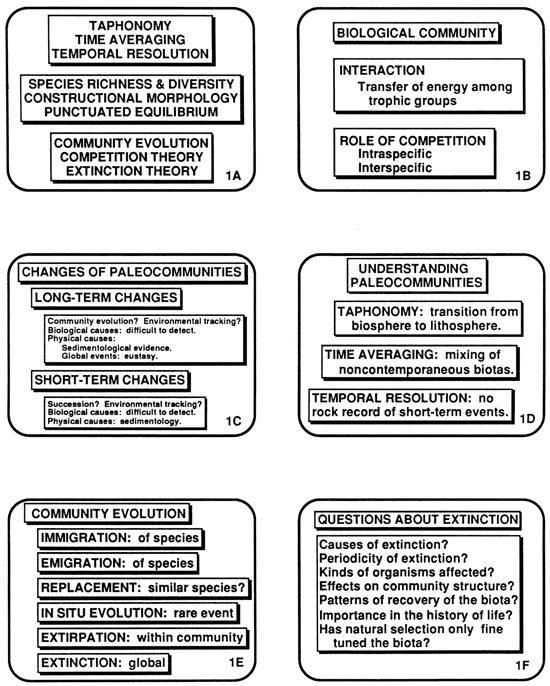
From the plethora of definitions of biological community (fig. 1B) one can distill the essence: communities comprise groups of organisms that live together and interact in various ways and to varying degrees (Kauffman and Scott, 1976; Strong et al., 1984). The interaction may include, but is not limited to, transfer of energy among trophic groups. Until recently, many ecologists judged that communities were structured by competition between species (McIntosh, 1987), and clear-cut instances of such competition have been observed (Smythe, 1986). It is now evident, however, that most of the competition in communities is between conspecific organisms. Moreover, other kinds of interactions are at least as important as competition (Price, 1984), and in many communities one searches in vain for conclusive evidence of the impact of interspecific competition (e.g., Price et al., 1984).
Changes that have taken place in paleocommunities, biological communities from the geological past (Scott and West, 1976), may be considered at various time scales (fig. 1C). Short-term changes took place over a few days or weeks, perhaps in response to short-term sedimentological events. Long-term changes may have taken as long as a geological period (see, e. g., Fagerstrom, 1987). Neglecting information about the relevant paleocommunities at any time scale will have a cost to a sedimentary model that is expressed in precision, realism, or generality (Levins, 1966; Schopf, 1972).
Understanding both qualitative and quantitative changes that have taken place in paleocommunities is complicated by three factors: taphonomy, time averaging, and temporal resolution in the stratigraphic record (fig. 1D). Taphonomy, while it tells us a lot about sedimentary processes, results in loss of paleobiological information because many organisms are selectively removed from the subfossil or fossil record by taphonomic processes (Lawrence, 1968, 1971; Kidwell and Behrensmeyer, 1988). Time averaging, caused by bioturbation, slow rates of sedimentation, and coarse sampling, obscures our understanding of communities by mixing organisms that did not live together (Walker and Bambach, 1971; Fursich, 1978). Finally, lack of temporal resolution, often a product of time averaging, means that whole classes of interesting and important, short-term, biological properties of ancient communities cannot be resolved in much of the sedimentary record (Schindel, 1980; Sadler, 1981).
Short-term changes in species richness and diversity and the whole process of succession (Connell and Slatyer, 1977) are typically impossible to resolve in much of the fossil record (Walker and Bambach, 1971). The causative environmental changes, however, are sometimes readily apparent from the sedimentological record. Pitfalls include expecting all important environmental changes that were important to organisms to have been recorded in the sedimentary rock and, conversely, expecting all sedimentological changes to have been important to organisms.
Long-term changes in species richness and diversity, if they were not taphonomically induced, are likely to have resulted from community evolution and environmental tracking by organisms or communities of organisms (Brondos and Kaesler, 1976). In principle, community evolution, the long-term, irreversible, temporal changes in the structure and composition of a community, can be tracked through thick sections of sedimentary rock such as occur in the late Paleozoic rocks of the midcontinent (Moore, 1964). The replacement of one species by another that results in community evolution (fig. 1E) does not imply that the two species are necessarily closely related nor that niches cannot be split and recombined. Either the precursor, its replacement, or both species may be missing from the fossil record.
Perhaps the most important changes in ancient communities have been mass extinctions (Jablonski, 1986). In recent years the search for the cause of mass extinctions has been intensive. Evidence now suggests that at least some mass extinctions in the geological past have resulted from bolides (McLaren, 1983), but other causes have been suggested as well. Understanding mass extinctions and incorporating them into sedimentary models depend on isolating their causes, and although the periodicity of mass extinctions (Sepkoski and Raup, 1986) may allow them to be fit into sedimentary models, we are a long way from understanding their effects. What kinds of organisms were most likely to have been affected by mass extinctions? %at were the concomitant effects on paleocommunities? Has extinction been of such overriding importance that natural selection has been relegated to the role of mere fine tuning? Seeking answers to these questions will occupy paleontologists for the rest of this century (fig. 1F) Their results will impact strongly the work of sedimentary modelers.
Brondos, M. D., and Kaesler, R. L., 1976, Diversity of assemblages of late Paleozoic Ostracoda; in, Structure and Classification of Paleocommunities, R. W. Scott and R. R. West (eds.): Dowden, Hutchinson & Ross, Inc., p. 213-234
Connell, J. H., and Slatyer, R. O., 1977, Mechanisms of succession in natural communities and their role in community stability and organization: American Naturalist, v. 111, p. 1,119-1,144
Fagerstrom, J. A., 1987, The evolution of reef communities: Wiley Interscience, 600 p.
Fursich, F. T., 1978, The influence of faunal condensation and mixing on the preservation of fossil benthic communities: Lethaia, v. 11, p. 243-250
Jablonski, D. J., 1986, Causes and consequences of mass extinctions-a comparative approach; in, Dynamics of Extinction, D. K. Elliott (ed.): John Wiley & Sons, p. 183-229
Kauffman, E. G., and Scott, R. W., 1976, Basic concepts of community ecology and paleoecology; in, Structure and Classification of Paleocommunities, R. W. Scott and R. R. West (eds.): Dowden, Hutchinson & Ross, Inc., p. 1-28
Kidwell, S. M., and Behrensmeyer, A. K., 1988, Overview--ecological and evolutionary implications of taphonomic processes; in, Ecological and Evolutionary Implications of Taphonomic Processes, A. K. Behrensmeyer and S. M. Kidwell (eds.): Palaeogeography, Palaeoclimatology, Palaeoecology, v. 63, p. 1-13
Lawrence, D. R., 1968, Taphonomy and information losses in fossil communities: Geological Society of America, Bulletin, v. 79, p. 1,315-1,330
Lawrence, D. R., 1971, The nature and structure of paleoecology: Journal of Paleontology, v. 45, p. 593-W7
Levins, R., 1966, The strategy of model building in population biology: American Scientist, v. 54, p. 421-431
McIntosh, R. P., 1987, Pluralism in ecology: Annual Review of Systematics and Ecology, v. 18, p. 321-341
McLaren, D. J., 1983, Bolides and biostratigraphy: Geological Society of America, Bulletin, v. 94, p. 313-324
Moore, R. C., 1964, Paleoecological aspects of Kansas Pennsylvanian and Permian cyclothems: Kansas Geological Survey, Bulletin 169, p. 287-380 [available online]
Price, P. W., 1984, Alternative paradigms in community ecology; in, A New Ecology--Novel Approaches to Interactive Systems, P. W. Price, C. N. Slobodchikoff, and W. S. Gaud (eds.): Wiley Interscience, p. 353-383
Price, P. W., Slobodchidoff, C. N., and Gaud, W. S., 1984, Introduction--is there a new ecology?; in, A New Ecology--Novel Approaches to Interactive Systems, P. W. Price, C. N. Slobodchikoff, and W. S. Gaud (eds.): Wiley Interscience, p. 1-11
Sadler, P. M., 1981, Sediment-accumulation rates and the completeness of the stratigraphic record: Journal of Geology, v. 89, p. 569-584
Schindel, D. E., 1980, Microstratigraphic sampling and the limits of paleontological resolution: Paleobiology, v. 6, p. 408-426
Schopf, T. J. M., 1972, Varieties of paleobiologic experience; in, Models in Paleobiology, T. J. M. Schopf (ed.): Freeman, Cooper & Company, p. 8-25
Scott, R. W., and West, R. R. (eds.), 1976, Structure and classification of paleocommunities: Dowden, Hutchinson & Ross, Inc., 291 p.
Sepkoski, J. J., and Raup, D. M., 1986, Periodicity in marine extinction events; in, Dynamics of Extinction, D. K. Elliott (ed.): John Wiley & Sons, p. 3-36
Smythe, N., 1986, Competition and resource partitioning in the guild of neotropical terrestrial frugivorous mammals: Annual Review of Ecology and Systematics, v. 17, p. 169-188
Strong, D. R., Jr., Simberloff, D. Abele, L. G., and Thistle, A. B. (eds.), 1984, Ecological communities--conceptual issues and the evidence: Princeton University Press, 613 p.
Walker, K. R., and Bambach, R. K., 1971, The significance of fossil assemblages from fine-grained sediments--time-averaged communities: Geological Society of America, Abstracts with Programs, v. 3, p. 783-784
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version May 12, 2010. Original publication date 1989.
URL=http://www.kgs.ku.edu/Publications/Bulletins/Sub12/Kaesler/index.html