![]()
![]()
![]()
Prev--Equability in the late Pleistocene || Next--North American central-highland fishes
Macrofossils of white and blue spruce (Picea glauca and P. pungens) and limber pine (Pinus flexilis), supplemented by rich landsnail and small-mammal faunas, indicate a taigalike successional mix of coniferous forest and aspen parkland on the loessal High Plains of western Kansas and Nebraska (now semiarid steppe) during the latest (Wisconsinan) glaciation of the Pleistocene. The fossil assemblages are most closely matched in the existing subalpine taiga of the Rocky Mountains.
Direct evidence on the paleovegetation and climate of the central and western Great Plains during the last glacial maximum (18,000±4,000 yrs ago) has been sparse relative to other regions of North America. The semiarid climate of the flat or undulating steppes is correlated with a general scarcity of lake sediments or bogs suitable for preservation of pollen or plant macrofossils. The region has largely frustrated palynologists, although substantial work has been published on Pleistocene records from the eastern periphery or from a few scattered sites within the Great Plains (Wendorf, 1961; Kapp, 1965, 1970; Wright, 1970; Wells, 1970a; Ritchie and DeVries, 1964; Moir, 1958; Watts and Bright, 1968; Watts and Wright, 1966; Ruhe, 1969; Gruger, 1973; King, 1973; Baker and Wain, 1985).
We now report dated plant and animal macrofossil evidence that indicates a taigalike, coniferous biome of Cordilleran (subalpine, Rocky Mountains) character on the central Great Plains of western Kansas and adjacent states during the latest (Wisconsinan, Weichselian) Pleistocene glaciation. Charcoalized wood and intact cones and leaves of conifers have been uncovered in close association with abundant shells of forest landsnails and bones of diverse small vertebrates in pleniglacial sediments; the fossil species are predominantly subalpine or boreal (even subarctic), though a small but significant contingent is now associated with temperate-deciduous, deep forest. Because the modern Plains region is largely treeless and, furthermore, has excessively hot summers with severe droughts, many of the species recorded in the late Pleistocene assemblages are now provincially extinct on the Great Plains. The central and northern Rocky Mountains, however, harbor extant populations of most of the boreal-subalpine species thus far recovered from Pleistocene sediments at our sites in the Central Plains (except, of course, the almost entirely extinct megafauna). Moreover, even within the northern Plains, there are numerous refugia for Pleistocene-relict species of trees, landsnails, and small mammals on forested ecological islands surrounded by steppe; the outstanding example is the Black Hills of South Dakota (fig. 1), but nonmountainous escarpments and canyons also serve as refugia for boreal or Cordilleran relicts (Wells, 1970b, c; McGregor and Barkley, 1977).
While reexamining the landsnail faunas that are ubiquitously preserved in the late Wisconsinan (Peorian) loess and associated alluvium in western Kansas and southwestern Nebraska (Leonard, 1952), we uncovered charcoalized wood fragments of spruce (Picea) at four widely spaced localities (Jewell, Graham, and Logan counties, Kansas, and Harlan County, Nebraska; fig. 1, table 1). At the Jewell County site (fig. 1, site 2; elevation 485 m [1,600 ft]), a thin streak of spruce charcoal extended horizontally for more than 100 m (330 ft) along a vertical cut in an upland deposit of Peorian loess (below the Brady paleosol). The Picea charcoal was 14C-dated to 14,450 yrs B.P., thus documenting a local forest fire in a late-Wisconsinan spruce flat. Scanning-electron microscopy of the charcoals from all four sites indicates that wood structures of Picea are almost perfectly preserved (fig. 2), but the species of spruce are difficult to distinguish on the basis of wood anatomy. Fortunately, however, several intact cones and numerous twigs and leaves, as well as abundant wood and charcoal of spruce were discovered together (in situ) at the Harlan County, Nebraska (North Cove), site (fig. 1, site 3; elevation 605 m [1,997 ft]), 110 km (66 mi) west-northwest of the Jewell County site. Picea wood from Harlan similarly dates to 14,770 yrs B.P. The cones and needles (fig. 3) enable the positive identification of the spruce as Picea glauca, the boreal white spruce of the Nearctic taiga that now grows from Alaska to Newfoundland and along the eastern flank of the Rocky Mountains to Montana, with outliers to the east on the Great Plains in the Cypress Hills of Saskatchewan and the Black Hills of South Dakota. The latter population is 600 km (360 mi) northwest of Harlan County and nearly 1,000 m (3,300 ft) higher in elevation. The fossil-rich North Cove site in Nebraska was uncovered by the authors in October 1983.
Figure 1--Map of United States and southern Canada with approximate extent of presettlement grasslands (gray); mountainous areas (black) and maximal limits of continental ice sheet during last glacial maximum (ca. 18,000 yrs ago) also are shown. Numbered square symbols are the sites of fossil records of cordilleran-boreal conifers, landsnail faunas, and small-mammal faunas located in or near the grassland province of the Great Plains and reported here: 1) Doniphan Co., KS, 12,420 yr BP; 2) Jewell Co., KS, 14,450 yr BP; 3) Harlan Co., NE, 14,770 yr BP; 4) Graham Co., KS, 17,930 yr BP; 5) Ellis Co., KS, 17,700 yr BP; 6) Harper Co., OK, 17,750 yr BP; 7) Logan Co., KS, 10,245 yr BP; 8) Fremont Co., CO, 22,720 yr BP (Neotoma middens); and 9) Bighorn Co., WY, 27,000-40,000 or more yr BP (Neotoma middens). Squares with dots (4, 8, 9) are macrofossil records of limber pine (Pinus flexilis); circles with dotted centers labeled P. flexilis are sites of living populations of limber pine, relict on scarps of Plains. Sites 2, 3, 4, and 7 have macrofossils records of spruce (Picea glauca at 3 and P. pungens at 7). Lettered dots north or east of above sites are previous macrofossil records of white spruce (P. glauca) from the Plains region: H = Hafichuk, Saskatchewan, Canada, 11,650 yr BP (Ritchie and DeVries, 1964); T = Tappen, Kidder Co., ND, 11,480 yr BP (Moir, 1958); PL = Pickerel Lake, Day Co., SD, 10,670 yr BP (Watts and Bright, 1968); R = Rosebud site, Todd Co., SD, 12,580 yr BP (Watts and Wright, 1966); IOWA group = ca. 20 spruce sites described by Ruhe (1969), 11,120-40,000 or more yr BP; M = Muscotah Marsh, Atchison Co., KS, 23,040 yr BP (Gruger, 1973); PT = Pomme de Terre group, Benton and Hickory cos., MO, 13,700, 16,580 yr BP, et al. (King, 1973). Isolated grasslands of Pacific Slope are PAL = Palouse Prairie; CAL = California Prairie.
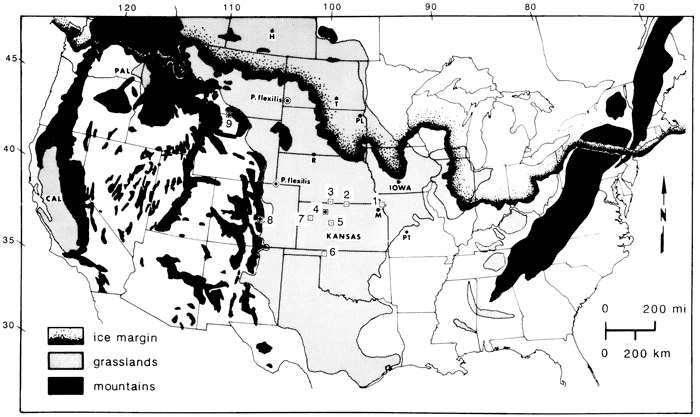
Table 1--Fossil assemblages of conifers and/or landsnails and small mammals from the central Great Plains; localities indexed to numbers on maps (figs. 1, 3). Quantitative landsnail data as % composition in samples of 103 to 3 X 104 fossil shells; Oklahoma site from Miller (1975). Lab numbers for 14C dates in order (left to right): Beta-12286, Beta-9320, GX-9355g, GX-9356, I-3460, GX-5812b, Beta-9321. Modern biogeographic headings in species list are generalized to indicate degree and direction of Holocene withdrawal from Plains.
| 14C date, yrs BP: | 14,700±100 | 14,450±140 | 179930±550 | 17,700±350 | 17,750±360 | 10,245±335 | 12,420±180 | |
|---|---|---|---|---|---|---|---|---|
| Locality: | Harlan Co., NE (3) |
Jewell Co., KS (2) |
Graham Co., KS (4) |
Ellis Co KS (5) |
Harper Co., OK (6) |
Logan Co., KS (7) |
Doniphan Co., KS (1) |
|
| Elevation, m: | 605 | 485 | 675 | 560 | 610 | 850 | 310 | |
| CONIFERS | ||||||||
| Cordilleran-boreal | ||||||||
| Picea spp. (charcoal) | + | + | + | + | ||||
| Picea glauca (leaves, twigs, cones) |
+ | |||||||
| Cordilleran | ||||||||
| Picea pungens (leaf) | + | |||||||
| Pinus flexilis (leaves) | + | |||||||
| LANDSNAILS | ||||||||
| Cordilleran | ||||||||
| Discus shimekii | 0.9 | 1.9 | 0.4 | + | ||||
| Pupilla blandii | 0.6 | 3.9 | + | + | ||||
| Cordilleran-boreal | ||||||||
| Columella alticola | 0.1 | 2.2 | 0.7 | |||||
| Pupilla muscorum | 0.6 | 0.1 | 5.3 | + | + | + | ||
| Vallonia gracilicosta | 56.2 | 15.4 | 63.4 | + | + | + | 26.3 | |
| Vertigo modesta | 5.1 | 35.1 | 13.0 | + | 1.5 | |||
| Cordilleran-boreal-eastern deciduous forest | ||||||||
| Cochlicopa lubrica | 0.6 | 0.4 | + | + | 0.3 | |||
| Discus cronkhitei | 5.6 | 20.0 | 1.2 | + | + | 7.6 | ||
| Euconulus fulvus | 0.4 | 4.7 | 0.6 | + | + | 0.3 | ||
| Nesovitrea electrina | 1.7 | 3.0 | + | + | 1.0 | |||
| Vertigo gouldii | 5.5 | 10.7 | 4.0 | |||||
| Cordilleran-eastern deciduous (disjunct) | ||||||||
| Helicodiscus singleyanus | + | |||||||
| Punctum minutissimum | 0.1 | 0.2 | 0.3 | + | 36.1 | |||
| Eastern deciduous forest | ||||||||
| Helicodiscus parallelus | 0.1 | + | + | + | ||||
| Hendersonia occulta | 0.1 | 0.5 | ||||||
| Stenotrema leai | + | |||||||
| Strobilops labyrinthica | 0.1 | + | ||||||
| Triodopsis albolabris | 0.9 | |||||||
| T. multilineata | 0.4 | |||||||
| Wide-ranging | ||||||||
| Gastrocopta armifera | 5.2 | 0.3 | + | + | ||||
| Hawaiia minuscula | 1.3 | 0.9 | + | + | 0.1 | |||
| Zonitoides arboreus | 2.7 | 0.5 | + | + | 1.4 | |||
| SMALL MAMMALS | ||||||||
| Cordilleran | ||||||||
| Eutamias minimus | + | |||||||
| Microtus montanus | + | + | ||||||
| Cordilleran-boreal | ||||||||
| Clethrionomys gapperi | + | + | + | |||||
| Lepus americanus | + | + | ||||||
| Microtus xanthognathus | + | + | ||||||
| Phenacomys intermedius | + | + | ||||||
| Sorex arcticus | + | + | + | |||||
| Synaptomys borealis | + | + | + | |||||
| Tamiasciurus hudsonicus | + | |||||||
| Cordilleran-Northern Plains | ||||||||
| Microtus pennsylvanicus (wide N., E.) |
+ | + | + | |||||
| Spermophilus kimballensis | + | + | ||||||
| S. tridecemlineatus (to Southern Plains) |
+ | + | ||||||
| Thomomys talpoides | + | + | + | |||||
| Zapus princeps | + | + | ||||||
Figure 2--Scanning electron microscopy of conifer charcoal from the grassland province of central Great Plains. [Note: images resized and magnification recalculated for web version.] a) Picea charcoal, radial section with vertical tracheids and ray tracheids (waffle pattern), the ray also sectioned tangentially (fore), x 178, Graham County, western Kansas (fig. 1, site 4), 17,930 yr B.P. b) uniseriate-bordered pits with tori, from (a) x 694. c) spiral thickenings and bordered pits of vertical tracheids radial section, x 180, Harlan County, Nebraska (site 3) 14,770 yrs B.P.
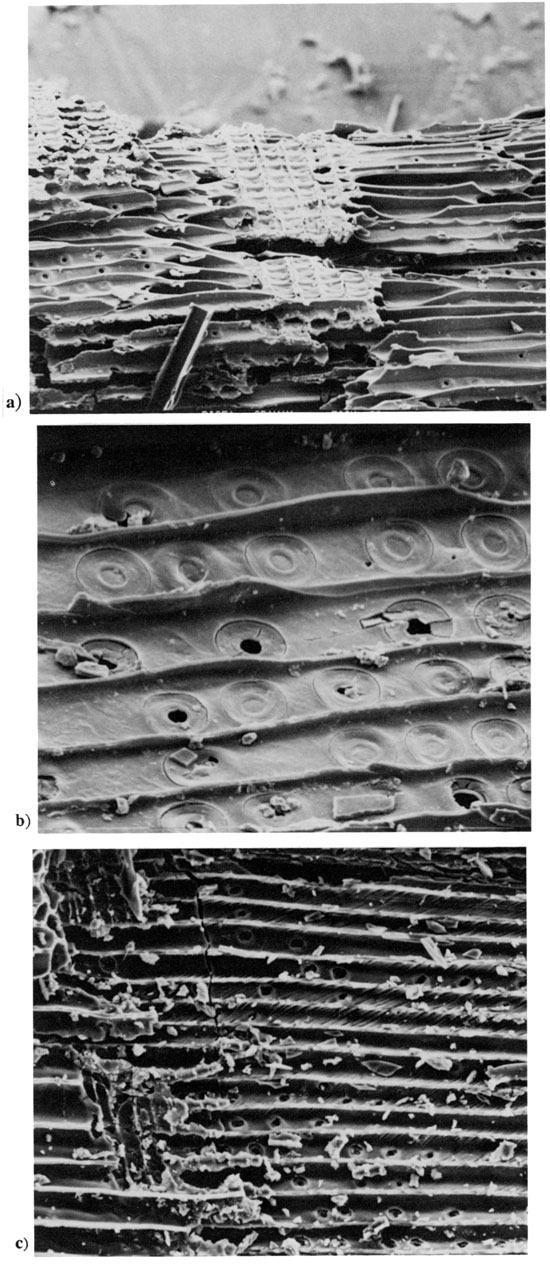
Figure 3--Macrofossils of Picea glauca from North Cove site, Harlan County, south-central Nebraska (fig. 1, site 3), 14,770 yrs B.P., the date on Picea wood. North Cove site was discovered October 1983, by Wells and Stewart, who later provided polleniferous sediments, other materials. and radiocarbon dates to other authors in this volume. [Note: images resized and magnification recalculated for web version.] a) branchlet with pegi!ke leaf cushions of genus, x 11. b) leaves, x 6. c) cones, opened and unopened (center), x 2.
On the other hand, the Logan County spruce-charcoal site (fig. 1, site 7; elevation 850 m [2,805 ft]) had a carbonized leaf of Picea pungens, a spruce now endemic to the Rocky Mountain region. The Picea pungens (blue spruce) was recovered from a paleoindian Bison-kill site in alluvial fill; a radiocarbon age on bone collagen of Bison was 10,245 yrs (Rogers and Martin, 1984). The Logan County site is 200 km (120 mi) southwest of the Harlan site and 90 km (54 mi) east of the Colorado/Kansas line. Ecologically, Picea pungens is the most likely cordilleran spruce to have extended eastward from the Rocky Mountains onto the High Plains during late-glacial time, as it extends downward along streams to lower elevations (to 2,000 m [6,600 ft]) than P. engelmannii in the Front Range of Colorado today; however, this is 300 km (180 mi) to the west and approximately 1,200 m (3,960 ft) higher than the Logan County spruce site. The alluvial records reported here are from small tributaries of streams with headwaters on the High Plains surface and not from stream systems arising in the Rocky Mountains (Platte, Arkansas); thus, the fossil materials must have been local in origin and not washed down from the mountains.
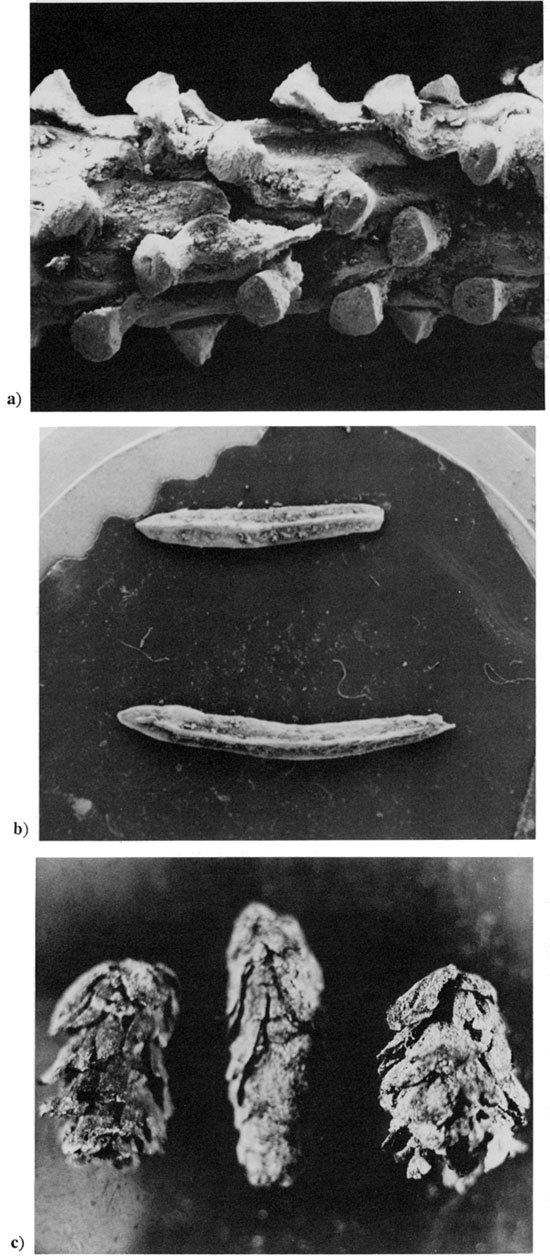
A definitively cordilleran character is imparted by the presence of fossil leaves of the Rocky Mountain limber pine (Pinus flexilis) at the Graham County Picea charcoal site (fig. 1, site 4; elevation 675 m [2,228 ft]), 120 km (72 mi) east-northeast of the Logan site or 90 km (54 mi) southwest of the Harlan site. Nine needle leaves of limber pine have thus far been retrieved from a silty loessal matrix containing abundant cordilleran-boreal animal fossils (landsnails and small vertebrates); collagen from bone of extinct Bison present in this fossil assemblage yielded a radiocarbon age of 17,930±550 yrs (Wells, 1983). The Pinus flexilis leaves were fragmented but are firmly identified (fig. 4). A major displacement of the subalpine limber-pine zone also is indicated by an independent source of macrofossil evidence: Neotoma (wood rat) middens (Wells, 1976) of full-glacial age from low elevations near the base of the Rocky Mountains. All the Pleistocene deposits thus far uncovered are dominated in composition by Pinus flexilis at sites ranging in elevation from 1,300 to 1,920 m (4,290-6,336 ft; fig. 1, sites 8 and 9). A deposit from the Front Range of Colorado, south of Pike's Peak (fig. 1, site 8), with a radiocarbon age of 22,720±270 yrs (GX-5925), is composed of almost pure limber pine (many thousands of five-needled leaf fascicles), but this site is 1,250 m (4,125 ft) higher in elevation and 450 km (270 mi) farther west than the Graham County, Kansas, record of Pinus flexilis (dated at 17,930 yrs B.P.). Today, limber pine is usually found only above 2,400 m (7,920 ft) in the Front Range; a few anomalous populations occur on escarpments of the High Plains, however, with the pine persisting as low as 1,600 m (5,280 ft) about 60 km (36 mi) east of Cheyenne, Wyoming (Wells, 1965). The closest living limber pines to the Wisconsinan fossil site in western Kansas are nearly 1,000 m (3,300 ft) higher in elevation and approximately 400 km (240 mi) to the northwest at Pawnee Buttes on the plains of northeastern Colorado (fig. 1 shows location of living populations on scarps in grasslands of North Dakota and Colorado, labeled P. flexilis). The remarkable High Plains populations of Pinus flexilis are widely disjunct and extremely local and are undoubtedly relicts of plains-wide Pleistocene populations.
Figure 4--Macrofossils or limber pine (Pinus flexilis) from Graham County. Kansas (Fig. 1, Site 4), 17,930 yrs B.P. [Note: images resized and magnification recalculated for web version.] a) fragment of needle-leaf, showing stomatal grooves on outer, convex face, x 8: b) transverse section of (a), showing characteristic pie-wedge shape of a pine with a five-needled fascicle, x 33.
South of the Laurentide Ice Sheet, large areas of the Missouri-Mississippi River drainage system were blanketed with sheets of loess (eolian silt) of varying thicknesses. The late Wisconsinan (Peorian) loess is thickest on bluffs along streams with silt-rich valley trains (a putative source), but also mantles the vast upland interfluves of the plains, thinning with distance from sources (Thorp and Smith, 1952). Loess is unfavorable for preservation of fossil materials prone to decay because of excellent aeration imposed by the interstitial pore space; pollen and plant macrofossils (except charcoal) are rare and poorly preserved in the loess, unless capped by an overburden of clayey till that impedes the diffusion of oxygen. For example, the Des Moines lobe of the Laurentide Ice in a late surge sealed with till the underlying, full-glacial Peorian loess, thus preserving the logs and stumps of a dozed-down forest of spruce and other conifers that grew on the loess of central Iowa (Ruhe, 1969). Plant macrofossils are excellently preserved in other unoxidized sediments, as in clay-rich pond and stream fills, occasionally exposed by fresh cuts in the loess.
The general scarcity of plant macrofossils, pollen, and vertebrate fossils in the loess itself is offset, on the other hand, by the wide occurrence of exquisitely preserved shells of landsnails in astonishing numbers (on the order of 10-100 shells per dm3 or liter of matrix). During the last glaciation, approximately 30 species of landsnails were extensively distributed on the immense, interfluvial uplands of the Great Plains and its eastward salient, the "Prairie Peninsula" of the Midwest (fig. 1), where the blanket of calcareous Peorian loess ensured preservation (Shimek, 1888, 1930; Frye and Leonard, 1952; Taylor, 1965). Many of these upland species of gastropods (the more characteristic loess snails are illustrated in fig. 5) are preserved also in clay-rich alluvial and lacustrine sediments, along with a rich fauna of aquatic mollusks that is entirely lacking in the upland loess. The remarkable ubiquity of pleniglacial landsnail faunas in loess on the High Plains of western Kansas (fig. 6) and their cordilleran-boreal character has been well documented by Leonard (1952).
Figure 5--Loess snails observed in present study (all x 5), illlustrating most of the more extralocal species (listed in table 1) that are now extinct on the Central Plains. [Note: images resized and magnification recalculated for web version.] The megasnails (Triodopsis sp.) were recorded only in northeastern Kansas, the shells of T. albolabris directly dated at 12,420 yrs B.P. The other species ranged westward on the Great Plains during the last glacial, leaving myriad fossil shells in loess and alluvium dated 14,000-18,000 yrs B.P.

Figure 6--Location of 93 landsnail faunas dating back to the most recent (Wisconsinan) glaciation of the Pleistocene in Kansas and adjacent states; most were analyzed by Leonard (1952). Numbered sites (open circles) are analyzed in this paper (table 1); site 6 from Miller (1975). Most of the 93 sites are in full-glacial loess of western Kansas.
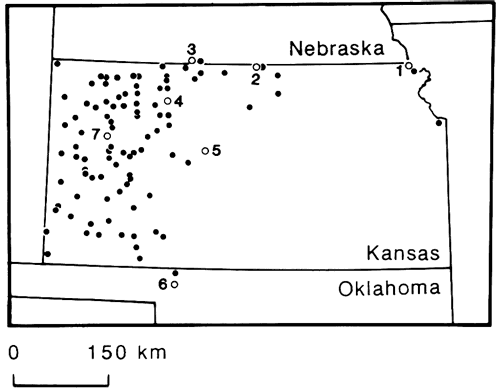
Perhaps the outstanding event of North American molluscan history in the late Quaternary was the virtual disappearance of this rich landsnail fauna from upland habitats of the Great Plains. At some time during the transition from late-glacial to Holocene climate (beginning at least 12,500 yrs ago), the pleniglacial landsnails of the Central Plains began to shift their ranges radically until they eventually became more or less restricted to the existing position of cordilleran-boreal forests in the Rocky Mountains, Canada, and the Northeast.
A small contingent of the loess snails of the Plains is now confined to the eastern deciduous forest. A notable example is Hendersonia occulta, formerly widespread on the loess sheets of the Midwest as far west as central Kansas, but now regionally extinct in the Plains and distinctly local even in the forests east of the Mississippi River (fig. 7). The few other species of this type extend westward in the Great Plains today only in riparian woodlands (e.g., Helicodiscus parallelus).
Figure 7--Distribution of a typical cordilleran-boreal landsnail, Vertigo modesta, and an eastern-deciduous-forest landsnail, Hendersonia occulta, both common in pleniglacial and late-glacial (at least as late as 12,420 yrs b.p.) loess of Kansas. The Vertigo was confined to the northern half (cf. Discus shimekii, fig. 8) and the Hendersonia to the eastern half of Kansas (solid black), where both species became extinct and are now widely extralocal (map from Leonard, 1952).
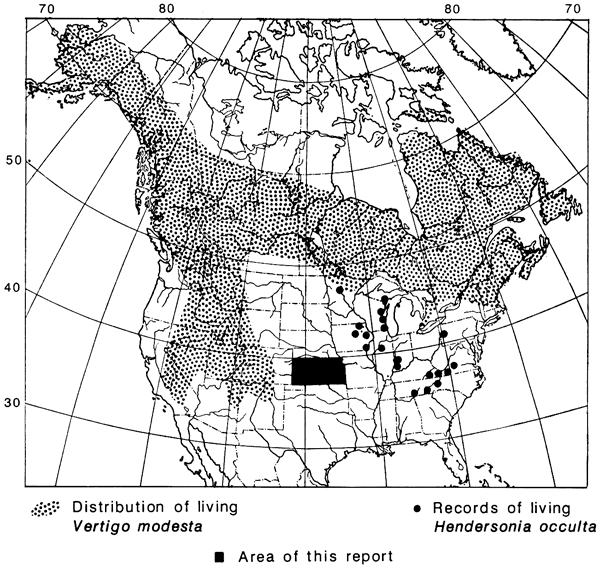
Aside from the deciduous-forest species, all of the landsnails that suffered extinction on the Plains have survived in the Rocky Mountains and some are now endemic to the Rockies (table 1). On the other hand, none of the erstwhile Plains species is endemic to the boreal taiga belt, although most range widely from Alaska to Newfoundland, and some are circumboreal through Eurasia (fig. 7).
Remarkably, the species that contracted to endemism in the Cordillera had pleniglacial ranges that entirely spanned the Great Plains, extending then across Iowa even into Illinois, east of the Mississippi River (e.g., Oreohelix strigosa, Discus shimekii; cf. fig. 8). The cordilleran-boreal Columella alticola reached Ohio (LaRocque, 1966; Miller and Wittine, 1972). However, the more subalpine or subarctic species have been recorded as fossils only within a limited zone to the south of the Laurentide Ice, and they did not occur in the southern Great Plains (fig. 8). For example, the presently montane Oreohelix is unrecorded even in Kansas, but their shells have been found in Peorian loess closer to the ice in Nebraska, Iowa, and Illinois (Prest and Rhodes, 1981). Perhaps along the front of the mountainous Ice Sheet, a marginal orographic zone of cool, cloudy climate stretched continuously from the Rockies in Montana southeastward through the Plains to the interior lowland of the Midwest, thus providing a migrational corridor for cordilleran landsnails (fig. 8).
Figure 8--Distribution of the cordilleran-subalpine landsnail, Discus shimekii, at present and during the last glacial maximum (of which it is considered to be an index fossil by Leonard, 1952); documented Wisconsinan occurrences are indicated by dots. The eastward limit in Illinois seems to correlate with the Laurentide ice barrier and farther south with a shift in forest composition. Note relatively northern southern limits along the east-west gradient from the Mississippi River west to western Kansas.
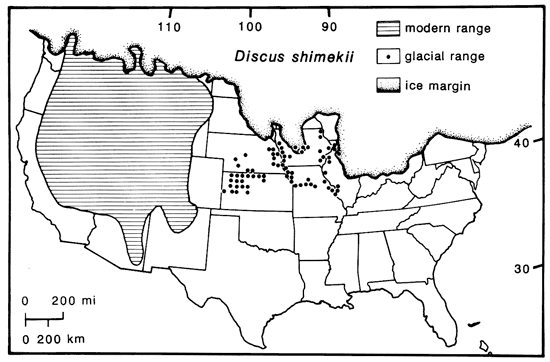
Other perhaps less-exacting species of cordilleran landsnails, such as Pupilla blandii, extended far from any mountains or ice on the southern Plains, e.g., on the Llano Estacado and Edwards Plateau of Texas; P. blandii was widely associated there with boreal-cordilleran Pupilla muscorum and Discus cronkhitei during the last glacial (Frye and Leonard, 1957), indicating much cooler and probably moister summers then in the southern Great Plains as well.
The timing of the great demise of cordilleran-boreal landsnails on the Great Plains is as yet poorly known. We have uncovered a rich landsnail fauna from stratigraphically young loess in northeastern Kansas (fig. 1, site 1; elevation only 310 m [1,023 ft]) that documents a partial persistence of full-glacial ("Upper Peorian") species mingling with perhaps more recently arrived "early Holocene" ("Bignell") species (e.g., the large polygrids Triodopsis albolabris and T. multilineata, indicative of temperate deciduous forest); this assemblage was dated to 12,420 yrs B.P. on the Triodopsis shells, which are supposed to be lacking (Leonard, 1952) in older loess here (table 1).
The main biogeographic implications of the late Pleistocene landsnail faunas preserved in pleniglacial loess and alluvium of the Great Plains are indicated in the organization of table 1. Clearly, the full-glacial environment of the Plains and entire Midwest was radically different from that of the modern grasslands, where very few of the ubiquitously abundant loess snails have persisted into the Holocene (Leonard, 1959). All of the loess snails of pleniglacial age (18,000±4,000 yrs ago) are more or less abundantly extant today, however, in forested regions adjacent to the Great Plains. Aside from the small contributions from snail fauna of the eastern deciduous forest, the loess assemblages are almost exactly matched by the main landsnail fauna from the subalpine taiga of the Rocky Mountains (Leonard, 1952; Pilsbry, 1939; Bequaert and Miller, 1973).
Ecological data on the modern landsnails of taiga indicate that they feed on decomposing plant litter, especially of leaves, twigs, or wood of forest trees or on associated fungi involved in the decomposition (LaRocque, 1966; Pilsbry, 1939). Grasslands are inhabited today by only a few wide-ranging generalists among the pleniglacial loess snails (e.g., Hawaiia minuscula, Zonitoides arboreus), which are also present in forest vegetation (where they are much more abundant). Like most of the invertebrate fauna of forest soils (Lutz and Chandler, 1951), landsnails are most abundant and diverse in litter provided by nutrient-pumping, broadleaf-deciduous trees (mull-type soils) and much less so in needle-leaf duff from conifers (mull-type soils). Thus, in the modern cordilleran-boreal taiga, assemblages almost identical to the Plains-wide Pleistocene loess-snail faunas are mainly restricted to successional phases of the forest dominated by deciduous trees (Karlin, 1961; Kralkaj, 1986), notably the ubiquitous aspen (Populus tremuloides). The snails are especially abundant and diverse in species in older stands of aspen with an understory of regenerating spruce, probably because there has been a longer buildup of rich, broadleaf litter and a cumulative recruitment of landsnails since the last forest fire. Catastrophic destruction by fire is the almost inevitable fate of the coniferous phase of subarctic or subalpine taiga (Slaughter et al., 1971; Ahlgren and Ahlgren, 1960; Lutz, 1956), even near the limits of forest growth (Payette and Gagnon, 1985), and spruce charcoal is of wide occurrence in the taiga (Payette and Gagnon, 1985). Quick-growing deciduous woodlands of aspen and birch are an almost taiga-wide feature of the fire-spawned succession (Halliday and Brown, 1943).
Because of the featureless continuity of the flat or gently undulating Plains. fires raging through the postulated Pleistocene spruce flats would have burned over immense areas, checked only at long intervals by abrupt topographic breaks (scarps, rivers), serving as fire breaks (Wells, 1965, 1970a, b, c). Hence. transient successional phases of cool-season grasses and other herbs using the Calvin or C3 photosynthetic pathway (e.g., fireweed, Epilobium angustifolium; fringed sage, Anemisia frigida) and more persistent deciduous trees (particularly aspen, which proliferates laterally by suckering from the widely spreading roots) are predictable components of a fire-mosaic in the full-glacial landscape of the Central Plains and Midwest (Wells, 1983). Direct evidence of fire in the spruce component , dating back to the last glacial maximum, is provided by our findings of spruce charcoal at four of the sites (table 1); the 100-m streak of Picea charcoal in the loess at one site affords a glimpse of a 14,500-yr-old burn in an upland stand of white spruce. Indirect evidence for the widespread upland occurrence of cool-temperate deciduous trees, such as aspen or birch (Betula papyrifera), is more widely contributed by the ubiquitous Pleistocene landsnail faunas (of cordilleran-boreal affinities) in the endless sheets of eolian silt that mantle the uplands of the Great Plains and Midwest.
A further indication of sera! patterning in a taigalike biome on the Great Plains is seen in the fossil bones of small vertebrates (chiefly rodents), co-occurring with the full- to late-glacial remains of the conifers and snails at several sites. Small, herbivorous mammals with limited feeding territories are more sensitive and faithful indicators of their plant communities than the widely mobile megafauna. Again, an outstanding predominance of cordilleran-boreal taxa is found among the retrieved fossils from the Central Plains (table 1). Most suggestive of the coniferous phase of taiga are the red squirrel (Tamiasciurus hudsonicus), redbacked vole (Clethrionomys gapperi), another boreal vole (Microtus xanthognathus), and the northern bog-lemming (Synaptomys borealis); all are present in the Rocky Mountains today, but the hyperboreal bog-lemming and Arctic shrew (Sorex arcticus) barely enter the United States in Montana and Minnesota. The phase of aspen with grassy openings is perhaps suggested by the western jumping mouse (Zapus princeps), the heather vole (Phenacomys intermedius), and the varying hare (Lepus americanus). all surviving today in the Cordillera. Stronger indicators of a more open, grassland phase include fossil remains of ground squirrels (Spermophilus tridecemlineatus and the extinct S. kimballensis, the latter represented today in the Rockies by S. elegans), a pocket gopher (Thomomys talpoides), and meadow voles (Microtus montanus and M. pennsylvanicus); again, these mammals are abundantly extant in the Rocky Mountain taiga (in the grassy-parkland phase).
The coexistence of all these ecologically diverse, taiga-inhabiting mammals with tell-tale charcoal of spruce is in agreement with the well-known fire cycle of the modern subalpine or subarctic taiga, entailing a relatively rapid succession from aspen parkland to spruce forest within a century or two (Slaughter et al., 1971; Ahlgren and Ahlgren, 1960; Lutz, 1956). Of course, the more open phases of the fire-induced succession could have accommodated the rich Pleistocene megafauna (e.g., Bison, Camelops, Equus, Mammuthus) as well, affording enhanced carrying capacity with both browsing and grazing niches. An increase in the tempo of burning after the advent of man in North America is a retrodicted possibility with intriguing consequences for the late Pleistocene biota of the Great Plains (Sauer, 1944).
Direct macrofossil evidence of cordilleran-boreal conifers (spruce, limber pine) at four scattered sites on the central Great Plains in western Kansas and southwestern Nebraska documents a taigalike vegetation during the latest Wisconsinan glacial (14,000-18,000 yrs ago) in habitats now dominated by grassy steppe of semiarid character. Closely associated fossil landsnail and small-mammal faunas also are cordilleran-boreal in their modern distributions, and most are regionally extinct in the Great Plains. The ecological requirements of the loess landsnails strongly indicate forest vegetation and more specifically suggest a cold-temperate, moist, broadleaf-deciduous phase, probably aspen (Populus tremuloides).
Some small-mammal fossils indicate a coniferous phase, while others suggest aspen-parkland or grassland-opening phases. Occurrence of spruce charcoal at four sites is direct evidence of fire in the coniferous phase. Hence, the diverse ecological indications at all of our sites are in agreement with the expected successional sequence in taiga vegetation following fire: 1) herbaceous phase with fireweed, sedges, grasses; 2) aspen-parkland phase; 3) aspen/spruce phase; and 4) mature spruce-forest phase. Fire in the mature-conifer stands reinitiates the cycle on a time scale that appears penecontemporaneous in the local fossil record, telescoping the various seral stages. The unbroken flatness of the vast interfluves on the Plains would permit a regional ravaging by fire, maintaining the vegetation in a state of successional flux that would maximize the coexistence of ecologically disparate fauna and flora among a kaleidoscopically shifting mosaic of seral niches in time and space. Thus, the central Great Plains was not a "loess steppe" during the most recent glaciation of the Pleistocene, but rather a biotically diverse, taigalike biome, with numerous species that now survive in the cooler and summer-rainy Rocky Mountains. The pleniglacial climate of the Great Plains was undoubtedly more equable than that of today, with much cooler, cloudier, and therefore effectively moister summers, minus the modern droughts.
We thank Wayne Ostlie for laboratory and field assistance. Research supported by National Science Foundation grants DEB 78-11187 and BSR-8320529 and by General Research Fund grants from the University of Kansas in 1983-84 and 1984-85 are all gratefully acknowledged.
Ahlgren, I. F., and Ahlgren, C. E., 1960, Ecological effects of forest fires: Botanical Review, 26, p. 483-533.
Baker, R. G., and Wain, K. A., 1985, Quaternary pollen records from the Great Plains and central United States; in, Pollen Records of Late Quaternary North American Sediments, V. Bryant, Jr., and R. Holloway, eds.: American Association of Stratigraphic Palynologists Foundation, Dallas, Texas, p. 191-203.
Bequaert, J. C., and Miller, W. B., 1973, The mollusks of the arid southwest: University of Arizona Press, Tucson, 271 p.
Frest, T. J., and Rhodes, R. S., 1981, Oreohelix strigosa cooperi (Binney) in the Midwest Pleistocene: Nautilus, v. 95, p. 47-55.
Frye, J. C., and Leonard, A. B., 1952, Pleistocene geology of Kansas: Kansas Geological Survey, Bulletin 99, p. 1-230. [available online]
Frye, J. C., and Leonard, A. B., 1957, Studies of Cenozoic geology along eastern margin of Texas High Plains: Texas Bureau of Economic Geology, Austin, Report of Investigations 32, p. 1-62.
Gruger, J., 1973, Studies on the late Quaternary vegetation history of northeastern Kansas: Geological Society of America, Bulletin, v. 84, p. 239-250.
Halliday, W. E. D., and Brown, A. W. A., 1943, The distribution of some important forest trees in Canada: Eeofogy, v. 24, p. 353-373.
Kapp, R. O., 1965, Illinoian and Sangamon vegetation in southwestern Kansas and adjacent Oklahoma: Contributions Museum of Paleontology, University of Michigan, v. 19, p. 167-255.
Kapp, R. O., 1970, Pollen analysis of pre-Wisconsin sediments from the Great Plains; in, Pleistocene and Recent Environments of the Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: University of Kansas Press, Lawrence, p. 143-155.
Karlin, E. J., 1961, Ecological relationships between vegetation and the distribution of landsnails in Montana, Colorado, and New Mexico: American Midland Naturalist, v. 65, p. 60--66.
King, J. E., 1973, Late Pleistocene palynology and biogeography of the western Missouri Ozarks: Ecological Monographs, v. 43, p. 539-565.
Kralka, R. A., 1986, Population characteristics of terrestrial gastropods in boreal forest habitats: American Midland Naturalist, v. 115, p. 156-164.
LaRocque, A., 1966-1970, Pleistocene mollusca of Ohio: Ohio Geological Survey, Bulletin 62.
Leonard, A. B., 1952, Illinoian and Wisconsin molluscan faunas in Kansas: University of Kansas, Paleontological Contributions, Mollusca, Article 4, p. 1-38. [available online]
Leonard, A. B., 1959, Handbook of gastropods in Kansas: University of Kansas, Museum of Natural History, Miscellaneous Publication 20, 224 p.
Lutz, H. J., 1956, The ecological effects of forest fires in the interior of Alaska: U.S. Department of Agriculture, Technical Bulletin 1133, p. 1-121.
Lutz, H. J., and Chandler, R. F., 1951, Forest soils: John Wiley and Sons, New York, 514 p.
McGregor, R. L., and Barkley, T. M., eds., 1977, Atlas of the flora of the Great Plains: Iowa State University Press, Ames, 600 p.
Miller, B. B., 1975, A series of radiocarbon-dated, Wisconsinan nonmarine, molluscan faunas from southwestern Kansas-northwestern Oklahoma; in, Studies on Cenozoic Paleontology and Stratigraphy, G. R. Smith and N. E. Friedland, eds.: University of Michigan, Museum of Paleontology, Ann Arbor, p. 9-18.
Miller, B. B., and Wittine, A. H., 1972, The origin of late Pleistocene deposits at Garfield Heights, Cuyahoga County, Ohio: Ohio Journal of Science, v. 72 p. 305-313.
Moir, D. R., 1958, Occurrence and radiocarbon date of coniferous wood in Kidder County, North Dakota: North Dakota Geological Survey, Miscellaneous Series 10, p. 108-114.
Payette, S., and Gagnon, R., 1985, Late Holocene deforestation and tree regeneration in the forest-tundra of Quebec: Nature, v. 313, p. 570-572.
Pilsbry, H. A., 1939-1948. Land mollusca of North America: Philadelphia Academy of Sciences, Philadelphia.
Ritchie. J. C., and DeVries, B., 1964, Contributions to the Holocene paleoecology of west-central Canada--a late-glacial deposit from the Missouri Coteau: Canadian Journal of Botany, v. 42, p. 677-692.
Rogers, R. A., and Martin, L. D., 1984, The 12 Mile Creek site--a reinvestigation: American Antiquity, v. 49, p. 757-764.
Ruhe, R. V., 1969, Quaternary landscapes in Iowa: Iowa State University Press, Ames, Iowa, 255 p.
Sauer, C. O., 1944, A geographic sketch of early man in America: Geographic Review, v. 34, p. 529-573.
Shimek, B., 1888, Notes on the fossils of the loess at Iowa City, Iowa: American Geologist, v. 1, p. 149-152.
Shimek, B., 1930, Landsnails as indicators of ecological conditions: Ecology, v. 11, p. 673-686.
Slaughter, C. W., Barney, R. J., and Hansen, G. M., eds., 1971, Fire in the northern environment--a symposium: Pacific Northwest Forest and Range Experiment Station, U.S. Department of Agriculture, Forest Service, Portland, Oregon, 275 p.
Taylor, D. W., 1965, The study of Pleistocene nonmarine mollusks in North America; in, The Quarternary of the United States, H. E. Wright, Jr., and D. G. Frey, eds.: Princeton University Press, Princeton, New Jersey. p. 597-611.
Thorp, J., and Smith, H. T. U ., eds., 1952, Pleistocene eolian deposits of the United States, Alaska, and parts of Canada: Geological Society of America, New York, map.
Watts, W. A., and Bright, R. C., 1968, Pollen, seed, and mollusk analysis of a sediment core from Pickerel Lake, northeastern South Dakota: Geological Society America, Bulletin, v. 79, p. 855-876.
Watts, W. A., and Wright, H. E., 1966, Late Wisconsin pollen and seed analysis from the Nebraska sandhills: Ecology, v. 47, p. 202-210.
Wells, P. V., 1965, Scarp woodlands, transported grassland soils, and concept of grassland climate in the Great Plains region: Science, v. 148, p. 246-249.
Wells, P. V., 1970a, Postglacial vegetational history of the Great Plains: Science, v. 167, p. 1,574-1.582.
Wells, P. V., 1970b, Vegetational history of the Great Plains-a postglacial record of coniferous woodland in southeastern Wyoming; in, Pleistocene and Recent Environments of the Central Great Plains, W. Dort, Jr., and J, K. Jones, Jr., eds.: University of Kansas Press, Lawrence, Kansas, p. 185-202.
Wells, P. V., 1970c, Historical factors controlling vegetation patterns and floristic distributions in the Central Plains region of North America; in, Pleistocene and Recent Environments of the Central Great Plains. W. Dort, Jr., and J. K. Jones, Jr., eds.: University of Kansas Press, Lawrence. p. 211-221.
Wells, P. V., 1976. Macrofossil analysis of wood rat (Neotoma) middens as a key to the Quaternary vegetational history of arid America: Quaternary Research, v. 6, p. 223-248.
Wells, P. V., 1983, Late Quaternary vegetation of the Great Plains: Nebraska Academy of Science, Transactions, v. 11 (special issue), p. 83-89.
Wendorf, F., ed., 1961, Paleoecology of the Llano Estacado: Museum of New Mexico Press, Santa Fe. 144 p.
Wright, H. E., Jr., 1970, Vegetational history of the Central Plains; in, Pleistocene and Recent Environments of the Great Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: University of Kansas Press, Lawrence, p. 157-172.
Prev--Equability in the late Pleistocene || Next--North American central-highland fishes
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version updated March 27, 2010. Original publication date 1987.
URL=http://www.kgs.ku.edu/Publications/Bulletins/GB5/Martin/index.html