Kansas Geological Survey, Bulletin 170, Part 5, originally published in 1964
Originally published in 1964 as Kansas Geological Survey Bulletin 170, Part 5.
Shale samples were obtained at one-foot intervals throughout a composite section of the Council Grove Group (Lower Permian) in north-central Kansas. Each sample was treated and thoroughly searched for microfossils, which were identified and their abundance in each sample recorded. All available microfossils were utilized for study of paleoenvironments in preference to reliance upon only one kind. Eleven kinds of foraminifers and 21 genera of ostracodes, as well as conodonts, holothurian sclerites, fish remains, fragments of inarticulate brachiopods, and charophytes were recognized and their abundance plotted. Stratigraphic distribution of taxa was determined and fossil assemblages were named after commonly occurring forms. Distinct assemblages of microfossils can be recognized which reflect cyclothemic deposition of the beds and which show a regular alternation that is judged to have paleoecologic significance. Paleoecology of the microfossils is interpreted by comparison with living representatives of the fossils (where possible), fossil associations, and containing shale types. A salinity gradient is postulated from fresh water through brackish to marine water, each category being represented by a distinct suite of microfossils. Within beds laid down under marine conditions different assemblages of microfossils are judged to have been controlled either by depth of water, salinity, or other important ecologic factors.
Plate 1--Relative abundance and distribution of microfossils in shales of the Council Grove Group (Lower Permian). Available as an Acrobat PDF file (1.3 MB).
The purpose of this study was to examine carefully microfossils found in the shales of a composite section of the Council Grove Group, Lower Permian, of north-central Kansas. The shales were sampled at one-foot intervals and microfossils obtained from each sample were identified and their relative abundance determined. (The location of measured sections is given under Localities Sampled.) Microfossil assemblages are described and their vertical distribution within the shales is discussed. The information gathered provides a basis for the interpretation of paleoenvironments of the shales and the relating of these ancient environments to Lower Permian cyclothems. Paleoecological interpretation of the assemblages is based primarily upon comparison with living relatives of some of the microfossils, shale lithologies, associated megafossils, and consideration of vertical sequential changes in assemblages throughout the Group.
The Council Grove Group is the middle group of the Gearyan Stage in Kansas, being overlain by the Chase Group and underlain by the Admire Group. The Council Grove Group is about 310 feet thick across Kansas and consists predominantly of limestones and shales. The rocks of the Group are divided into 14 formations which consist of an alternation of seven limestone and seven shale formations. None of the shale units are divided into members, but the lower five limestones (Bader, Beattie, Grenola, Red Eagle, and Foraker) are divided into three or more members. Four of the five limestone formations listed above consist of two limestone members and a middle shale member. The Grenola Limestone comprises three limestones with two intervening shales. The upper two limestone formations (Crouse, Funston) are not divided into members. The formations and members of the group are shown on Plate 1.
The shales of the Council Grove Group exhibit wide variation in color--red, green, gray, brown, and black. Some shale formations may enclose beds of coal (Eskridge, Stearns), or gypsum (Easly Creek). There has been little study of the grain size or mineralogy of the shale formations, although shale members of limestone formations have been studied (Imbrie, 1955; McCrone, 1964). Sandstone is a minor lithologic type in the Group, and no sandstone beds were encountered in the sections that provided the basis for this study.
The most topographically conspicuous beds in the Council Grove Group are the limestones because they are more resistant to weathering than intervening shales. The thickest and most persistent limestones are found in the lower part of the Group, and several limestones have been given formational or member rock-stratigraphic designation. In addition to thicker limestone beds that have been given names, thin, nonpersistent layers of limestone commonly are included in the shale formations. A wide variety of lithologic types of limestone are present in the Group, and some of the lateral variations in lithology and fossil content of the limestones have been studied in detail, especially in recent studies by Imbrie, et al. (1959), Laporte (1962), and McCrone (1964).
In the first part of the study the major kinds of microfossils found are discussed and their stratigraphic distribution and arrangement into assemblages noted. The second part of the study comprises paleoecological interpretations.
Seven sections within the Council Grove Group were measured during the summer of 1958, and the shales contained in them were sampled at one-foot intervals. These sections afford a composite section through the Group, encompassing 14 formations. Samples were treated and fossils isolated and identified during 1958-59; reference sections were checked and additional samples collected in August, 1959.
Each sample was divided and one-half was soaked in kerosene, then boiled in water with a small amount of sodium carbonate, and the fine fractions decanted. The samples were then dried and sieved. The fraction between US no. 35 and 115 sieves was treated further. Samples that did not disaggregate easily by the above method were pulverized with a rubber-tipped pestle and treated again. The sieved fraction was then placed in a 50 ml beaker with a small amount of wetting agent (commercial detergent) and water and put in an ultrasonic vibrating tank for one to five minutes, then decanted and dried. It was found that the vibrator was not useful in the first stages of breaking down the sample, but that it was extremely valuable in removing small bits of clay and silt from fossils after they had been released from the matrix.
Each sample was examined under the binocular microscope and specimens mounted on cardboard slides. Initially, a weight of one gram of treated sample was used and an attempt was made to pick all the fossils from that amount. This method soon proved unsatisfactory. It was impossible to remove or break down all the fine shale particles in the samples. Some samples consisted almost entirely of small shale particles with relatively few fossils, whereas other samples, which broke down completely, were composed almost entirely of fossil debris. This difference in proportion of shale and fossil material is due to the ease and completeness with which the shale is disaggregated and not to actual differences in abundance of microfossils. It was found by examination of the entire treated sample that rare microfossils frequently are not present in a given one-gram sample. Consequently, this method was abandoned and the entire prepared sample (5 to 40 g) from each one-foot interval was picked thoroughly on three separate occasions. It was found that some rare or inconspicuous forms were overlooked on first or even second inspection of the sample. Additional microfossils doubtlessly could be discovered with additional collecting. This method of repeated picking is judged to be more reliable than the first-mentioned one, although more time consuming. After the samples were picked, the aggregate number of specimens in each taxon on the slides was tabulated and entered on the chart showing distribution of microfossils in the Group (Plate 1).
The following sections constitute a complete composite section throughout the shales of the Council Grove in north-central Kansas in the vicinity of Manhattan, from which samples were obtained which yielded the microfossils used in this study.
Grateful acknowledgment is due Daniel F. Merriam, of the Kansas Geological Survey, for continued encouragement, the paleontologic staff of the Geology Department, University of California, Los Angeles, for helpful discussion of some of the problems, and E. W. Fager, Scripps Institution of Oceanography, who gave advice concerning the statistical methods used here. Jay Smith aided in preparation of the samples, and Mrs. Peter Kurtz drafted the illustrations.
Thirty-seven kinds of microfossils were recognized in the samples used in this study. These include 10 genera of Foraminifera (other than long-axis fusulinids, which were noted but not identified), 21 genera of ostracodes, fragments of the brachiopod Orbiculoidea, conodonts, holothurian sclerites, fish teeth and scale fragments, and charophytes. No attempt was made to identify the genera of the latter four categories. The microfossils are discussed below.
Remains of the calcified oogonia (gyrogonites) of these algae were found in abundance in the basal Eskridge Shale and Johnson Shale and scattered throughout the Blue Rapids Shale. Many specimens are crushed or distorted; the majority are black, although some light brown and a few clear white gyrogonites were found. Apparently several types are represented. Only Catillochara moreyi (Peck) has been reported from the Lower Permian of Kansas (Peck and Eyer, 1963). Fossil charophytes are generally believed to have lived in brackish or fresh water, as do the living genera (Peck, 1953).
Fusulinids--Long-axis fusulinids were found extensively in the Hughes Creek Shale Member of the Foraker Limestone, but only a single occurrence was noted above this shale, that being in the lower part of the Neva Limestone Member of the Grenola Limestone. These large protozoans occur only in limestones up through the Beattie Limestone (Thompson, 1954). Many of the specimens obtained were small, immature individuals of the same size range as other microfossils. No attempt was made to identify genera.
Globivalvulina--This foraminifer is the most abundant and widespread protozoan in shales of the Group. It reaches peak abundance in the dark gray shales of the Hughes Creek. It occurs typically with a variety of other microfossils in beds that contain a conspicuous fauna of larger invertebrates. Globivalvulina was not found in thick shales between prominent limestone units. Where the genus is found in abundance, there is considerable range in its size and number of chambers.
Endothyranella(?)--This foraminifer is most common in the Hughes Creek Shale and was not found above the lower part of the Neva Member. [Note: This foraminifer was examined by Dr. Doris E. Nodine Zeller, State Geological Survey of Kansas, The University of Kansas, who stated: "The external form of this foraminifer resembles Endothyranella, but the internal structure and secondary deposits, which can be seen in thin section, resemble those of Endothyra. It cannot be related to any previously described genus. The two specimens seen in thin section were highly recrystallized. The stratigraphic occurrence of this form is higher than that ever recorded for Endothyra or Endothyranella."]
Nodosinella--This genus occurs commonly in the Hughes Creek Shale but was not found in younger rocks of the Group. Specimens dissolved in dilute hydrochloric acid yielded no residue of non-calcareous material.
Tetrataxis--Tetrataxis is the next most abundant smaller foraminifer in the Council Grove Group. It was found in more samples (45) than any other foraminifer, but many of these occurrences consisted of one or two specimens only. Isolated specimens of the genus commonly appear slightly worn, with rounded edges, perhaps indicating transportation prior to incorporation in the sediment. The genus ranges throughout the Group but is most common in the Florena Shale Member of the Beattie Limestone and shales of the Neva and Eiss Limestones. There is little variation in size of specimens, and small, clearly immature specimens are rare.
Textularia--Although Textularia was found in 11 samples, most of these occurrences consisted of only one or two specimens. Only samples from the Neva Limestone and the Hamlin Shale contained more than eight specimens. [Note: Specimens recorded as Textularia from the lower Neva shale (samples 115 and 116, Pl. 1) have been examined by Dr. Helen Tappan Loeblich, Department of Geology, University of California at Los Angeles, who believes that "they are the juvenile stages of the Climacammina that is found in the same sample, rather than a species of Textularia."]
Glomospira--Only two specimens were discovered in the middle part of the Hughes Creek Shale, and both of these are similar to Glomospira diversa Cushman and Waters.
Deckerella--Specimens of Deckerella were found in the Hughes Creek Shale and Neva Limestone. Individuals in the Neva are over 1 mm in length, and are consistently larger than specimens from the Hughes Creek. The later, uniserial part of the test is commonly crushed.
Climacammina--This large foraminifer was found only in the shale unit of the lower Neva. Specimens 2 mm in length occur there in association with many other microfossils. The uniserial portion of the test is commonly flattened and distorted.
Ammodiscus--This genus is especially common in a single sample from both the Eskridge Shale and Funston Limestone. No specimens were obtained from units older than the Eskridge. The specimens are typically light brown or ivory in color and exhibit considerable range in size within a single sample. When these specimens are digested in acid, the test disintegrates, and a fine residue of silt- and clay-sized particles remains. Specimens of Textularia, Glomospira, Deckerella, and Climacammina, when similarly treated, also, disintegrate completely. Except in shales of the Eiss Limestone, beds containing Ammodiscus typically do not yield a great variety of other microfossils.
Ammovertella--Specimens assigned to this adnate genus exhibit a wide variety in form. Some specimens. are still attached to fragments of brachiopod spines or shell fragments. Others have broken loose, and the open underside of the test can be seen. All specimens retain their shape after treatment in hydrochloric acid, although some calcareous cement is removed, because all specimens effervesced. Ammovertella is common in shales just above or below thin limestones containing abundant Osagia--a form-genus of intergrown calcareous algae and Ammovertella (Lane, 1958)--indicating that environments were suitable for Ammovertella both before and after peak algal growth occurred. The genus has been found associated with similar algal limestones in Pennsylvanian rocks of Kansas (Ireland, 1956).
In terms of number of recognizable genera, ostracodes are the most abundant microfossils in the Council Grove Group. Specimens consisting of both complete carapaces and single valves are common. Each fossil, whether single or complete, was counted as a single specimen. Some genera, e.g., Hollinella, were found only as single valves. Others, e.g., Cavellina, ?Coryellites, Healdia, consisted almost exclusively of complete carapaces. Still others, e.g., Geisina, Carbonita, occurred in some samples predominantly as complete carapaces and in other samples as single valves. Immature instars are common in some samples and rare in others. Such moult stages could be identified to genus and were included in counts of abundance.
The large number of known genera of late Paleozoic ostracodes presents a problem in the generic assignment of such a large number of individuals as are reported here. Diagnostic characters separating closely similar genera are commonly variable in nature and gradational forms are not uncommon. A conservative approach is used here, with relatively few genera recognized.
Carbonita--This ostracode is the most abundant one in the shales of the Council Grove Group. Large numbers of specimens were obtained from many samples, commonly in association with only a few other microfossils. The genus has been recognized for many years in the European Carboniferous as a fresh- or brackish-water form. Scott (1944) first pointed out the presence of "fresh-water" ostracodes in Pennsylvanian cyclothems of Illinois and lower Permian beds of Pennsylvania and West Virginia.
The isolated abundant occurrence of this genus clearly indicates a habitat distinct from that of the majority of the microfossils which are associated with larger marine invertebrates or foraminifers.
?Candona--These specimens seem to be related to Candona planidorsata Cooper from the Pennsylvanian of Illinois. The genus is found in abundance only in the Hooser Shale Member of the Bader Limestone and the Eskridge Shale and is invariably associated with Carbonita.
Gutschickia--Specimens referred to this genus do not occur below the Eskridge Shale and are found commonly only in the Hooser Shale. Most specimens are preserved with both valves intact.
Darwinula--Individuals identified as Darwinula are similar to specimens of D. pungens illustrated by Cooper (1946). The genus is not abundant in the Council Grove samples, but it is most common in the Stearns Shale.
Geisina--Ostracodes identified as Geisina are commonly associated with Carbonita. Kloedenellid ostracodes from Pennsylvanian and Permian rocks were largely referred to a single genus (Jonesina) in older literature, but species now are assigned to many genera. Jonesina howardensis Kellett is the only described species from Kansas similar to specimens here referred to Geisina. The genus does not occur below the Grenola Limestone; it is especially common in the Salem Point Shale Member of the Grenola Limestone and the Hooser Shale.
Hollinella.-This large, distinctive ostracode was found throughout the Group. Hollinella typically occurs as isolated valves and commonly only a few specimens are obtained from each sample. The valves usually are broken, crushed, or show evidence of wear, suggesting pre-burial transportation.
Bairdiacypris--This genus typically occurs in beds that also contain Bairdia, and it is not commonly found except in association with the latter genus.
Bairdia--This genus was found in many samples throughout the section, but is rare or absent in shales such as the Johnson and Eskridge, which lie between limestone formations.
?Coryellites--Identification of this genus is questionable because most, but not all, species assigned to Coryellites by Cooper (1946) have been reassigned to Bythocypris (Cordell, 1952). In this study populations of bythocyprid ostracodes, especially in the Hughes Creek Shale, that accord with Cooper's original description of the genus are tentatively assigned to Coryellites.
Sansabella--Specimens of this genus are similar to Jonesina bolliaformis (Ulrich & Bassler) figured by Kellett (1933), which has been placed in Sansabella (Cooper, 1946). The genus is common throughout the Council Grove Group in shales containing other marine microfossils.
Oliganisus--Ostracodes of this type are not common in the samples and were recognized definitely in only two samples.
Monoceratina--This small distinctive ostracode occurs sparsely in but four samples.
Pseudobythocypris--This genus occurs only in the Legion Shale Member of the Grenola Limestone and in shales of the Easly Creek. [The old name Bythocypris is not used here, as it applies to a different kind of ostracode (Shaver, 1958).]
Amphissites--This ostracode is common throughout the Group and especially in shales of the Neva Limestone, Florena Shale, and Eiss Limestone Member of the Bader Limestone. [The genus Ectodemites Cooper, which is closely similar to Amphissites, is considered to be a junior synonym of the latter (Echols and Creath, 1959).]
Kirkbya--This ostracode was found in only six samples. It is especially common in shales of the Neva Limestone Member.
Knightina--Knightina was found in seven samples, its greatest abundance being in the Eiss Limestone Member. Like Kirkbya, this genus occurs only rarely below the Neva Limestone.
Silenites--This ostracode was collected from shales in the upper Eskridge, the Bennett Shale Member of the Red Eagle Limestone, and the Hughes Creek. Silenites does not occur in abundance in any sample.
Cavellina--Abundant carapaces of Cavellina occur in the Hughes Creek Shale, where the genus is the most common ostracode. Specimens are also plentiful in shales of the Neva, but only a few specimens were obtained from samples higher than the Neva. Cavellina occurs in only one sample in the Florena and is absent in the Eiss, although both units contain a wide variety of other ostracodes. This genus is almost always found with both valves intact, and single valves are rare.
Kellettina and Roundyella--These two genera are found only in the upper and lower shales of the Neva Limestone Member, in association with many other microfossils.
Healdia--Healdia is most common in the Hughes Creek Shale, but ranges throughout the Group. The genus typically occurs sparsely in most samples and is restricted to shales that also contain a variety of other microfossils.
A total of nine shale samples contained conodonts, but eight of these yielded fewer than five specimens. Individual conodonts commonly are broken and most specimens are of the platform type, similar to Idiognathodus. Two specimens each of a Hindeodella- and a Prioniodus-like form were found in the lower shale of the Neva. Conodonts were found in black shales in the Hughes Creek, but they also occur in dark gray shales and are most abundant in brown shales of the lower Neva. Specimens were not found in black shales in the Legion and Bennett Shale Members. All but one of the conodont-bearing samples are from the Grenola Limestone or older beds in the Group. A single specimen was found above the Grenola in the uppermost Speiser Shale, at the very top of the Group.
Calcareous ossicles or sclerites of holothurians are common in many of the marine shales of the Council Grove Group. Four distinct types of sclerites can be recognized, of which the small wheels called Protocaudina are most common. These are especially abundant in the Hughes Creek Shale, Bennett Shale, Neva Limestone, and Florena Shale members. In some beds small wheels are associated with hooks, such as Ancistrum, which are less widely distributed and less common than the wheels. A third type of sclerite consists of convex disks, flattened on one side and strongly convex on the other. The convex side has a cribrate, imperforate surface and a central depression at the apex. This form is similar to Etheridgella, but it lacks the elevated apex of that genus.
The fourth type includes small, flat, pentagonal disks that have a fine reticulate ornamentation on both sides and are imperforate. These are abundant in the Eiss Member, but they were not found elsewhere. No known genus of sclerite seems to be similar to them. The latter two types do not occur in association with wheels or with each other and presumably represent remains of different holothuroids.
The Etheridgella-type sclerite is common in the Eskridge Shale (Samples 90 and 91), where it is associated with hooks, but not wheels; this type is also common in one bed (Sample 48) where it occurs with no other sclerites but with common Ammodiscus and Hollinella. The flat-disk sclerites were found only in the upper Eiss shales, without other sclerites in association.
The sclerite associations can be grouped as follows: wheels and hooks, Neva, Speiser, and Hughes Creek; wheels alone, Florena and Hughes Creek; Etheridgella alone, Easly Creek; Etheridgella and hooks, Eskridge; flat pentagonal disks alone, Eiss.
These different associations probably represent remains of several distinct holothuroids that lived in different environments.
Bony fragments of fish are found in many beds of the Council Grove. These fossils consist of small, whole scales, broken larger scales, denticles, and a variety of teeth, including small, blunt teeth and sharp, elongate ones. A few teeth of the pleurocanth shark type were encountered, and many of the teeth and scales probably belong to paleoniscid fish. Although these vertebrate fossils were found in 44 samples, they consist commonly of only one or two specimens. Abundant fish remains were found only in shale samples from the Legion Shale, lower Neva Limestone, Funston Limestone, and Blue Rapids Shale. Three samples containing abundant fish remains also contain Orbiculoidea and the other one (Blue Rapids) contains only two charophyte gyrogonites.
Although the fish fossils are too fragmentary to be identified as to genus, an occurrence of similar fish remains in the Roca Shale, discovered by the author in 1959, is noteworthy. At a locality 1.4 miles east of Fairview, Kansas, in a road cut on the north side of Kansas Highway 36, a thin stringer of limestone one inch thick and three feet long yielded large quantities of small fish teeth and scales that were removed from the matrix with dilute acetic acid. The limestone matrix consisted largely of innumerable carapaces of the ostracodes Carbonita and Geisina.
The vertebrate material was submitted to T. H. Eaton, Museum of Natural History, The University of Kansas, who reported:
"This material (UKMNH No. 1756) contains many scales of paleoniscid fish, with isolated teeth and bone fragments of the same, but the genus is not determinable (perhaps Elonichthys); one piece is a fragment of the occipital portion of a braincase. The paleoniscides were primarily fresh-water fishes.
"Also there are a few very small teeth of Xenacanthus, a fresh-water pleuracanth shark, and at least two Cladodus teeth (this name refers to a type of shark tooth, but the genus is uncertain, as several different ones have such teeth). Thus it is highly probable that the deposit is from fresh water rather than a marine environment."
Many fossils besides the ones discussed here were observed in the shale samples. These are not included on the distribution chart but deserve brief mention. Fragments of echinoid plates and spines, small crinoid columnals, portions of fenestrate and ramose bryozoans, productid brachiopod spines, and shell fragments of a variety of macroscopic invertebrates, especially brachiopods, were found. These are most common in the Hughes Creek, Neva, Florena, and Eiss samples.
Black shale in the lower part of the Legion Shale (Sample 128) yielded numerous small, brown, twiglike fragments of plant material that could not be identified.
Molluscan remains are present in a number of samples including ones that do not contain other macrofossils. These fossils consist of small pelecypods and gastropods. Most specimens are fragmentary, crushed, or are internal molds that could not be identified. Some of these were probably immature individuals. Internal limonitic molds of high-spired gastropods and small nuculoid pelecypods are present in dark gray shales of the Hughes Creek. A variety of small pelecypods, including ones that show hinge structure, and high- and low-spired gastropods are present in the Roca Shale in association with Carbonita. Fragments of very thin-shelled, extremely low-spired, discoidal gastropods are present in association with Carbonita. Internal molds of small high-spired gastropods are present in beds that also contain Ammovertella, such as in the Hooser Shale Member (Sample 51).
Janesville Shale
Hamlin Shale Member--Seven one-foot samples were obtained from the upper part of the Hamlin Shale to provide information about microfossils in beds just below the Council Grove Group. The lowest three samples were unfossiliferous; the next sample contained Carbonita and Darwinula. The upper three feet of the Hamlin contains a microassemblage comparable to that in the Hughes Creek Shale above.
Foraker Limestone
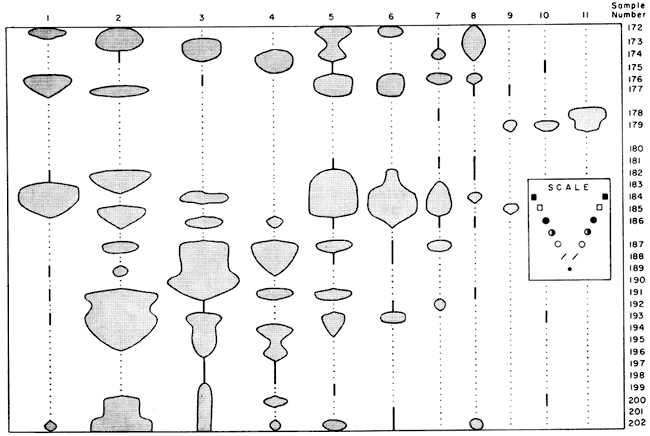
Hughes Creek Shale Member--At the locality sampled the Hughes Creek provides the thickest and best documented record of marine microfossil distribution. Fusulinids, Globivalvulina, Cavellina, ?Coryellites and Bairdia are abundant (Fig. 1). ?Coryellites occurs in 14 samples in this Member and in only eight other samples in the remainder of the Group. The dark gray shales contain an abundant macrofauna of invertebrates, as well as an extensive microfauna. Black shale near the top of the Member has a fauna restricted to fragments of Orbiculoidea, fish teeth and scales, and conodonts. Limy shale near the base of the Member (Samples 197 and 198) yielded only a few specimens largely because of difficulty in preparation of the calcareous samples. The characteristic assemblage of this shale is virtually continuous up through the Member, beginning next above the American Limestone Member of the Foraker Limestone and continuing to the base of the Long Creek Limestone Member of the Foraker Limestone. The microfauna is essentially similar above and below thin limestones within the shale of the Hughes Creek, except where black shales intervene. Fusulinids are more common and widely distributed in this Member than in any other shale in the Group. The Hughes Creek becomes increasingly limy farther south in Kansas, until it becomes a fusulinid-bearing, cherty limestone (Moore, et al. 1951).
Figure 1--Distribution of common microfossils in the Hughes Creek Shale. Width of "balloons" indicates relative abundance according to scale above. Relative abundance symbols and sample numbers are the same as on Plate 1. Microfossils: 1, fusulinids; 2, Globivalvulina; 3, Cavellina; 4, ?Coryellites; 5, Bairdia; 6, Tetrataxis; 7, Bairdiacypris; 8, Sansabella; 9, conodonts; 10, fish teeth and scales; 11, Orbiculoidea.
Johnson Shale
In sharp contrast to the uppermost beds of the Hughes Creek, the basal shale of the Long Creek Limestone contains a charophyte-Carbonita-Orbiculoidea assemblage. Charophytes are restricted to the lower six feet of the unit, but no fossils were obtained from the lowest foot. Carbonita reaches peak abundance in the middle part of the Johnson, and here occurs with rare to common fish fragments. Both Carbonita and fish remains continue up to the base of the Glenrock Limestone Member of the Red Eagle Limestone.
Red Eagle Limestone
Bennett Shale Member--The mottled brown and black lower part of the Bennett contains fish remains and fragments of inarticulate brachiopods. These fossils are replaced in the upper part of the shale by abundant holothurian sclerites, Bairdia, and Silenites. Although fusulinids are common in the Glenrock Limestone below, none were found in the Bennett.
Roca Shale
The Roca Shale is one of the least fossiliferous of the shales sampled. Apart from Carbonita in the middle of the shale, few other microfossils were encountered. The most abundant fossils are fragments of small, probably immature, pelecypods and gastropods (abundant in Sample 136; rare in Sample 140). It was impossible to identify these molluscan fragments.
Grenola Limestone
Legion Shale Member--The Legion Shale, where sampled, consisted of a lower black and an upper brown shale. The lower part contains fish remains and orbiculoid brachiopod fragments and a few Tetrataxis. The upper brown part contains Sansabella, and Hollinella, together with Bairdia and Ammovertella in the upper one foot. Ammovertella carries over into the Burr Limestone Member of the Grenola Limestone as part of the form genus Osagia, an association of algae and this adnate foraminifer.
Salem Point Shale Member--This shale member marks the reappearance of the Carbonita assemblage within the Grenola. Carbonita and Geisina are both abundant, especially in the upper half of the Member. Fish teeth and scales are the only common microfossils in the upper one foot.
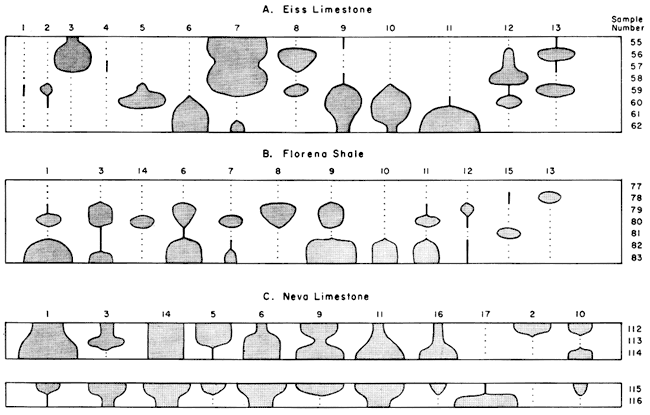
Neva Limestone Member--Although this Member is largely limestone, it includes a two-foot shale near the base and a three-foot shale near the top. Both of these shales contain an abundant and varied microfauna (Fig. 2C). The lowest sample in the Neva (No. 116), consisting of brown shale, contains abundant conodonts, fish teeth, and orbiculoid fragments similar to the assemblage in the black shales in the Hughes Creek. The upper three feet of shale contains a varied ostracode-foraminifer assemblage dominated by Globivalvulina and Bairdia. Orbiculoidea and conodonts were found in only one sample above the Neva.
Figure 2--Distribution of common microfossils in shales of the Eiss Limestone, the Florena Shale, and the Neva Limestone. For explanation see Figure 1. Microfossils: 1, Globivalvulina; 2, Healdia; 3, holothurian ossicles; 4, Textularia; 5, Knightina; 6, Amphissites; 7, Sansabella; 8, ?Coryellites; 9, Bairdia; 10, Bairdiacypris; 11, Tetrataxis: 12, Hollinella; 13, Ammodiscus; 14, Cavellina; 15, fish teeth and scales; 16, Kirkbya; 17,conodonts.
Eskridge Shale
The Eskridge Shale samples begin with microfossils, among which charophytes and Carbonita are especially abundant. Ammodiscus is first found in the lower Eskridge Shale. A thin shale bed (No. 100) between two thin limestone beds contains Ammodiscus and Hollinella. ?Candona and fish teeth are associated with abundant Carbonita above this thin shale. The upper Eskridge contains a more varied assemblage, including holothurian sclerites, Deckerella, Cavellina, and Silenites. The highest beds in the Eskridge contain Bairdiacypris, Hollinella, and abundant Ammovertella, clearly associated with a thin algal limestone at the base of the Cottonwood Limestone Member of the Beattie Limestone.
Beattie Limestone
Florena Shale Member--An abundant and varied assemblage of microfossils is present in the Florena (Fig. 2B), including Globivalvulina, Bairdia, Amphissites, and Tetrataxis. There is a gradual decrease in abundance and variety of the assemblage from bottom to top of the shale and the upper part of the Florena contains only a few fish teeth, Ammodiscus, Geisina, Darwinula, and Carbonita, foreshadowing the typical Stearns assemblage above.
Stearns Shale
Just above the algal Morrill Limestone Member of the Beattie Limestone the Stearns Shale contains rare Cavellina, ?Coryellites, Bairdia and Ammodiscus, which do not occur farther up in the formation. Carbonita and fish remains typify the Stearns microfauna, as well as associations of Carbonita, Geisina, Gutschickia, and Darwinula.
Bader Limestone
Eiss Limestone Member--There is an abrupt faunal change from the highest Stearns Shale to lowest shale in the Eiss, on either side of a thin limestone marking the base of the Bader. The Eiss is predominantly shale at the exposure sampled and contains a microfauna dominated by Sansabella, which increases in abundance upward in the Member (Fig. 2A). Bairdiacypris, Tetrataxis, and Amphissites are confined to the lower half of the unit and all decrease in abundance upward. Holothurian ossicles and Ammodiscus occur most commonly in the upper part.
Hooser Shale Member--Thin brown and gray shales of the Hooser contain a typical Carbonita assemblage. Red and green shale are present in the Hooser at other localities. The microfossils increase in variety, but not abundance, upward in this Member. Gutschickia and ?Candona are present only in the upper half. Hollinella and Ammovertella occur in the upper foot just below algal limestones of the Middlesburg Limestone Member of the Bader Limestone.
Easly Creek Shale
With the exception of the Roca, the Easly Creek contains the most sparse micro-assemblage in the Group. Of 16 samples only two yielded microfossils. The lowest two-foot shale of the formation contains holothurian remains, Oliganisus, Sansabella, Hollinella, Ammodiscus, and fish remains. The middle part of the Easly Creek is largely red and green unfossiliferous shale, and in Marshall County, Kansas, the basal portion of the unit contains an eight-foot bed of gypsum.
Blue Rapids Shale
Rare to moderately common microfossils in the Blue Rapids Shale include charophytes, Carbonita at the base, and scattered Tetrataxis and fish teeth.
Funston Limestone
A thin two-foot green shale in the Funston carries abundant fish remains and scattered Orbiculoidea fragments in the lower foot and abundant Ammodiscus in the upper foot.
Speiser Shale
Carbonita dominates the microfauna of the Speiser, except for the upper two feet, which yielded an assemblage consisting of Globivalvulina, Tetrataxis, Amphissites, and holothurian sclerites, just below the Threemile Limestone Member of the Wreford Limestone, the basal unit of the Chase Group. Ammovertella is restricted to a one-foot shale at the base of the formation, just below a thin Osagia limestone.
It was hoped that after the microfossils in each sample had been picked, identified, and their relative abundance and stratigraphic occurrence plotted, they could be grouped into fossil assemblages based on common occurrences that would have paleoecologic significance. A number of preliminary methods for deciding which fossils consistently occurred together in (or were both absent from) the same samples, and which utilized some sort of index of affinity or association, were tried and found wanting. The grouping method proposed by Fager (1957) and employed by Johnson (1962) finally was used, although the index of association suggested by each author above was not utilized. Fager and McGowan (1963) have suggested an index of the form:
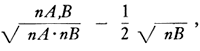
where nA,B is the number of joint occurrences of taxa A and B, nA is the total number of occurrences of taxon A, nB the total number of occurrences of taxon B, such that nA≤nB can be used as a basis for this grouping method. Fager found a cutoff value of 0.50 appropriate for the material with which he has been working. It was found that this cutoff value led to only one group in the Permian microfossils. As this was unsatisfactory, other values were tried and finally a cutoff value of 0.250 was accepted. At the 0.250 level there was a total of 81 pairs of taxa with association indices at this level or higher. These pairs involved 23 of the 37 categories of microfossils used in this study, which were then grouped according to Fager's method (Fig. 3B).
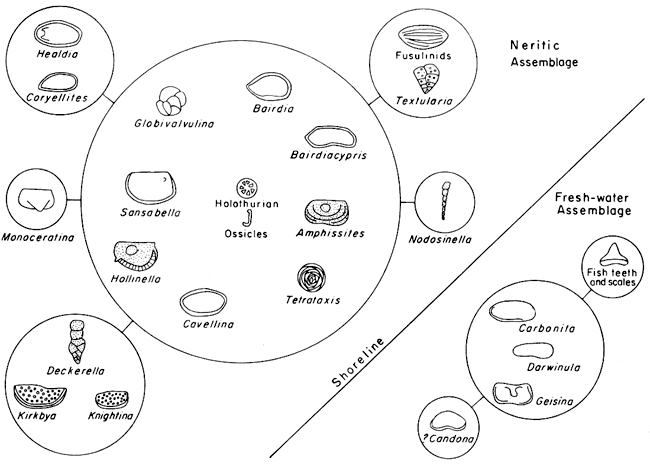
The groups of mutually associated taxa include one large group of nine that consists of Cavellina, Bairdia, Tetrataxis, Bairdiacypris, Amphissites, Hollinella, Globivalvulina, Sansabella, and holothurian ossicles (Fig. 4). Exclusively associated with members of this group were Nodosinella and Monoceratina. Two groups of three each were formed that included Deckerella, Knightina, and Kirkbya, as well as Carbonita, Geisina, and Darwinula, of which the latter group was exclusively associated with fish teeth and scales and ?Candona. Two groups of two each consisted of fusulinids and Textularia as well as ?Coryellites and Healdia.
Figure 3--A, Number of joint occurrences (within box) and total number of occurrences (diagonal) of microfossils from 202 shale sample stations (Plate 1) in the Council Grove Group. Numbers along left margin and bottom correspond to numbers of taxa on Plate 1; B, Joint occurrences of taxa that had indices of affinity of 0.250 or greater, using Fager's (1957) index.

The following samples (see Plate 1) are considered representative of each of the groups defined above:
Among the 14 identified taxa that had no association indices of 0.250 or higher are included a number of forms that are based upon their distribution and occurrence, to have significance for environmental interpretation (Fig. 5). Consequently, all taxa not included in the groups above that occur in four or more samples are listed below with the other two taxa with which they had highest and next highest indices of affinity. The remaining six genera occur in only one or two samples. The eight taxa listed below will be considered in the next section on interpretation of the assemblages.
The preceding discussion has set forth salient features of the kinds of microfossils present in shales of the Council Grove Group, their stratigraphic distribution through the Group, and the assemblages of small fossils present. This data provides the raw materials for paleoecological interpretation of the microfossils and consideration of past environments.
Environmental interpretation may be drawn from a number of lines of evidence. Comparison of the occupied habitats of living plants and animals that are also recorded as fossils together with evidence derived from fossil associations and their enclosing sediments may provide clues to ancient environments. Earlier paleoenvironmental reconstructions based on the same kinds of fossils found in other areas or in rocks of differing ages may be useful. The vertical distribution of individual taxa and assemblages throughout the Group and their relationship to each other allow inferences as to past environmental conditions. These lines of evidence provide the bases for the paleoecological part of this study.
One of the fruitful lines of paleoecological investigation of fossils in Cenozoic and, to some extent, in Mesozoic rocks has been the comparison of environments inhabited by living plants and animals with that postulated for their fossil analogues. The validity of this inductive method rests on the assumption that organisms have not appreciably changed their environmental requirements through long spans of time. In older and older rocks comparisons become increasingly difficult, partly because increasingly larger categories of organisms have become extinct and because the prerequisite assumption of non-evolution of habitat requirements must be made for increasingly long periods of time.
If such an approach is attempted in a study of early Permian microfossils, the assumption must be made that whatever taxonomic level is considered, whether genus, family, or order, its habitat has not been appreciably altered through the last 200 million years. At present there is no adequate test that can be applied to such an assumption to determine its validity. Most paleontologists probably would agree that community evolution and consequent shifting of environmental requirements must have taken place continuously throughout geologic time, just as organic evolution has proceeded, apparently without interruption.
Charophytes--Living charophytes are found principally in fresh-water ponds and lakes, in quiet water on soft bottoms. A few live in brackish water, e.g., in the upper reaches of the Baltic Sea, where they are found in water of salinity up to about 6‰ (Segerstråle, 1957). The assumption is made that the early Permian charophytes reported in this study also lived in fresh water, although it is certainly a possibility that the oogonia might have been carried out into saline waters before deposition.
Foraminifers--Three genera of foraminifers found in the Council Grove Group are still living--Textularia, Ammodiscus, and Glomospira. Living species of Textularia have been reported from water as shallow as 12 feet (Bandy, 1953) to as deep as 12,000 feet (Cushman, 1911). Ammodiscus has been reported from depths of 18 feet (Cushman and Valentine, 1930) to depths of 18,000 feet (Cushman, 1911). The genus is typically found on a mud or sand bottom. Glomospira has been recovered from depths of about 50 feet to over 10,000 feet. Clearly, the total bathymetric range of extant genera of foraminifers that are also found in Lower Permian rocks is of little significance.
Ostracodes--Among early Permian ostracodes identified the following genera are still extant: Candona, Darwinula, Bairdia, and Monoceratina. Bairdia is typically a marine ostracode and is commonly found at depths of 20 to 50 meters (Grekoff, 1956), but it may also be found in littoral environments. Monoceratina is benthonic, living in neritic marine waters. These two genera then should be indicative of deposition under marine conditions.
Candona is common today in fresh-water lakes and streams, but it is found also in shallow brackish water of salinity up to 2 or 3 parts per thousand (Wagner, 1957). In Europe Darwinula occurs in much the same range of salinity and depth as Candona, but Van den Bold (1958) reports Candona and Darwinula in Trinidad, the former as typical of upper courses of rivers, 400 to 2,000 feet above sea level, and Darwinula 50 to 600 feet above sea level.
Both Candona and Darwinula are considered to be typically fresh-water ostracodes today. They occur in shales of the Lower Permian together with charophytes, which are also mainly fresh-water, as well as with teeth and scales of paleoniscid and pleuracanth fishes, both of which are considered to be fresh-water in habitat. Consequently, the Geisina-Carbonita-Darwinula assemblage with its association of ?Candona and fish teeth and scales is interpreted to represent a fresh-water group of organisms in shales of the Council Grove Group.
The fresh-water Carbonita assemblage is not entirely excluded from slightly brackish-water habitats. Presumably, however, these ostracodes, along with charophytes and fishes, lived in waters not so closely related to the strand line, perhaps in shallow pools and ponds on a broad backshore area in which there were fresh- or brackish-water lagoons. The commonly imbricated nature of Carbonita valves is suggestive of quiet, shallow water in which the carapaces became dissociated and then washed to and fro, resulting in many valves stacked one within the other. The Carbonita assemblage is more closely associated with red and green shales than any of the other groups, although it is much more common in green than in red shales. This would suggest a broad subaereal backshore of fine red muds with semipermanent ponds of fresh or slightly brackish water scattered over the flat. The red muds, by reduction, became green muds in these bodies of water. The entire area probably was only slightly above sea level but may have been very broad, so that ponds on the landward side could have been a relatively great distance from the open sea.
The large group of nine taxa that includes Bairdia, and holothurian sclerites, as well as the three smaller groups of foraminifers and ostracodes, are judged to have been marine organisms (Fig. 4).
Figure 4--Assemblages of microfossils obtained by using Fager's (1957) grouping method and their interpretation in terms of a salinity gradient.
The presence of groups of organisms that are interpreted to have lived in fresh-water habitats versus those that lived in marine waters leads to the postulate that within the shales studied one of the factors affecting distribution of microorganisms is a salinity gradient from fresh water at one end to water of normal marine salinity at the other end. It follows from this that ancient shorelines would be represented within the rock record of the Council Grove Group, and that some of the shales would represent brackish-water conditions of deposition intermediate in salinity between fresh and completely marine waters.
Among fossils that could indicate the brackish-water conditions hypothesized above, the two most likely are Ammodiscus and Orbiculoidea. Neither of these two genera is closely enough associated with any marine or freshwater form to have been included in groups of clearly fresh-water or marine habitat. Ammodiscus is most closely associated with Carbonita, on the one hand, and Hollinella, on the other hand, and so can be inferred to have lived in brackish water. Orbiculoidea has the closest index of affinity with fish teeth and scales and Carbonita, but since no brachiopods, so far as is known, live or have lived in fresh water, this genus is interpreted to have lived in brackish waters close to the shoreline (Fig. 5). If Ammodiscus and Orbiculoidea do represent part of a brackish-water fauna, the preserved elements of this fauna are clearly less diverse than are preserved parts of the fresh-water or marine faunas. In other words, diversity increases from brackish into fresh water or from brackish to marine water, a situation that is also true today (Gunter, 1947, 1947a). Intertidal or littoral environments and near-littoral lagoons, estuaries, and periodically submerged mud flats are the environments that are characterized by such conditions today, and the Orbiculoidea-Ammodiscus group may have lived in such habitats as these.
Figure 5--Grouping interpretation of microfossils that had no indices of affinity of 0.250 or greater.
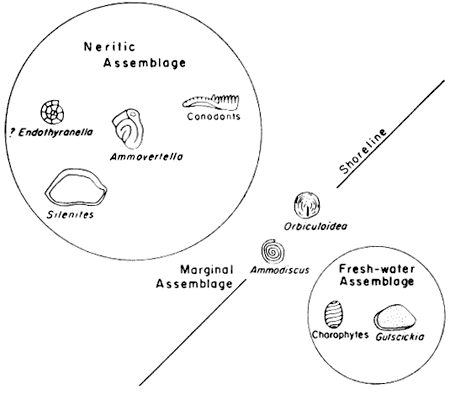
The assemblages found in shales of the Council Grove Group have their closest described counterpart in ostracode-foraminifer assemblages described from the Namurian and Westphalian of the Ruhr District of Germany (Kremp, 1952). Kremp was able to recognize three principal microfaunal assemblages that occur between spore-bearing beds closely associated with coal on the one hand, and Lingula and marine megafossil-bearing beds on the other hand. Succeeding fossil phases, from top to bottom, of the German cycles are: a. micro- and megaspores; b. ostracodes of the Carbonita group (Carbonita, Candona, and Gutschickia); c. ostracodes of the Jonesina group (Jonesina, Sansabella, and Sulcella); d. arenaceous foraminifers (Ammodiscus, Glomospira, Glomospirella, and Hyperammina); e. Lingula; and f. small goniatites, brachiopods, pelecypods, and gastropods.
The typical sequence in these European beds clearly represents an almost complete record of the regressive hemicycle; transgressive phases are confined to beds just above coals and are poorly represented or absent. Commonly minor fluctuations within the regressive hemicycle are recorded. The Carbonita group of ostracodes is essentially the same as the assemblage recorded here from the Council Grove strata. The Jonesina group is sulcate and seemingly corresponds to Geisina as used in this report, which is commonly found in association with Carbonita. The Lingula phase corresponds to the Ammodiscus-Orbiculoidea brackish-water assemblage as interpreted here and marine megafossils to the marine microfaunal assemblages of the Council Grove. The rocks under study here present a much better record of marine microfossils than is found in the Carboniferous of the Ruhr. However, the similarity of the ostracode assemblages is strikingly similar, considering the difference in the ages of the rocks. In both cases the controlling factor is judged to have been salinity of water.
Distribution of microfossils in the Council Grove Group (from bottom to top) will be considered in this section. Two aspects of vertical distribution are considered. First, the median sample station occurrence of each microfossil within the Group; second, the vertical sequence of microfossils within more or less continuous, thinner, shale intervals.
Casual inspection of the stratigraphic distribution of microfossils within the Group (Plate 1) reveals that there are conspicuous differences in occurrence of specific microfossils. Fragmentary Orbiculoidea, conodonts, ?Coryellites, and fusulinids are largely restricted to the lower half of the Group. Nodosinella and Glomospira occur only in the Hughes Creek Shale Member. Ammodiscus is present only in the Eskridge Shale and higher beds, and Ammovertella is not found below the Legion Shale Member. In order to provide a more complete picture of vertical distribution of all taxa, the median sample station occurrence of each form within the Group was determined (Table 1). The shale sample numbers on Plate 1 are used to record the median occurrences, which are based on total number of feet of rock in the Group in the sections measured. When a fossil occurred in an even number of samples, the middle of the interval between the two samples on either side of the median was listed. The median bed of shale, excluding limestones, is between samples 101 and 102 in the Eskridge Shale. This is close to the median bed (Sample 98) of the Group, including the limestones. A total of ten taxa have median occurrences above the median thickness of the group, with Ammodiscus and Gutschickia having the highest medians. Twenty-one taxa have median occurrences below the median interval of the group, six fossils not being considered because they occurred in only one or two samples.
Table 1--Median sample station occurrences of microfossils in the Council Grove Group, Lower Permian. Sample numbers same as on Plate 1.
| Taxa | Median Sample Number |
|---|---|
| Gutschickia | 57 |
| Ammodiscus | 59 |
| Darwinula | 74 |
| Geisina | 77 |
| Fish teeth | 78 |
| Amphissites | 83 |
| Ammovertella | 84 1/2 |
| Hollinella | 87 1/2 |
| Monoceratina | 95 1/2 |
| ?Candona | 97 1/2 |
| 98--Median bed of Group | |
| Carbonita | 99 1/2 |
| 101 1/2--Median shale sample | |
| Sansabella | 105 |
| Charophytes | 106 |
| Holothurians | 112 |
| Knightina | 113 |
| Kirkbya | 113 1/2 |
| Tetrataxis | 113 1/2 |
| Bairdiacypris | 115 |
| Textularia | 116 |
| Deckerella | 146 1/2 |
| Bairdia | 148 |
| Silenites | 148 |
| Conodonts | 158 1/2 |
| Orbiculoidea | 160 |
| Endothyranella | 163 1/2 |
| Globivalvulina | 173 |
| Cavellina | 179 1/2 |
| Fusulinids | 183 1/2 |
| Healdia | 186 |
| ?Coryellites | 187 1/2 |
| Nodosinella | 189 |
Most of the taxa in the fresh-water assemblage have median occurrences high in the Group. Many of the fossils in the main and subsidiary neritic assemblages have low median occurrences, including especially the Coryellites-Healdia group. There is a distinct gap in medians between Textularia (median sample, 116) and Deckerella (median sample 146 1/2). There are less distinct breaks in the distribution of medians between Ammodiscus and Darwinula, Hollinella and Monoceratina, Silenites and conodonts, and an Endothyranella-like foram and Globivalvulina.
The median sample occurrences of all 31 taxa considered here range through a total of 133 feet of shale samples and through 174 feet of rock, including limestones. This range of medians, including approximately 60 percent of the total thickness of the Group, strongly suggests that there is a gradual change in gross composition of the microfauna through the Group apart from the cyclic occurrences of individual taxa through a number of units. This gradual faunal change includes reduction in number of occurrences of some taxa from bottom to top, as well as appearance or increase in abundance of other taxa in higher beds. The conclusion is that superimposed on the cyclic arrangement of microfossils and rock types there was a gradual environmental change taking place during deposition that did not obviously affect the gross physical character of the rocks. In Permian rocks above the Council Grove Group (e.g., Sumner and Nippewalla groups) there is an increasing abundance of unfossiliferous clastics and evaporites, including redbeds, gypsum, salt, and shales, that may represent fresh-water, marginal marine, and hypersaline deposits. Gradual faunal changes observed in the Council Grove Group are probably related to gradual restriction of the marine basin of deposition, expansion of nonmarine depositional areas, including shallowing of, as well as gradually increasing salinity of, marine water in which the sediments were laid down. Some of the microfossils were not able to adjust to increasing salinity and shoaling, and so are mainly confined to lower beds in the Group; other microfossils were able to tolerate such changes and made their first appearance or increasingly flourished in the younger strata. In order to test this hypothesis it would be necessary to study microfossil assemblages in the Admire Group (below) and the Chase Group (above).
Not only was the vertical distribution of individual taxa of microfossils throughout the entire group a consideration, but the nature of sequential changes in distribution of contained fossils through smaller intervals of shale were considered and were deemed to have significance. Some shales contain essentially the same microfossils from bottom to top, whereas others exhibit gradual changes in the types of fossils present. Four sequential patterns of distribution of microfossils can be observed as follows:
The four distributive patterns outlined above can be interpreted as follows: (1) It seems obvious that some shales (pattern 1) record long-continuing deposition of shale in approximately the same environment. Shales that show examples of this pattern commonly contain Carbonita and other members of the fresh-water assemblage. (2) Some thin limestone layers (limestones between samples 2 and 3, and 177 and 178) within thicker shale sequences seem to mark relatively sudden, temporary, changes in the environment of deposition, as denoted by microfossils, whereas other thin limestone layers (within the Eskridge Shale, for example) present within shale sequences have similar microfaunas just above and below. These latter limestones seemingly were deposited in the same environment as the shales enclosing them, and perhaps record temporary cessation of clastic sedimentation but continuing deposition of carbonate materials. The first-mentioned thin limestone beds may have been formed during relatively abrupt changes in the environment of deposition, or accumulated during a longer period of time marked by non-deposition of shale, during which the environment gradually changed, so that when shale began to be deposited again, the microfossil assemblage had changed in response to environmental change. (3) Either the abrupt change from clearly marine limestone to shale with a very different (Carbonita assemblage) microfossil aspect is due to a relatively rapid change in depositional environment, not recorded at the limestone-shale interface, or to a gradual change within the limestone bed. (4) A gradual change in the environment of deposition is recorded by deposits containing a gradually altering microassemblage.
Patterns (1) and (4), then, are taken to indicate long stability and gradual environmental change respectively. If the other two patterns also involve long-continuing gradual changes, then patterns (2) and (3) may be taken to be characteristic of sequences of rock that indicate less stability and abrupt environmental changes. Many of the examples of patterns (2) and (3) involve the Carbonita assemblage. This assemblage has been interpreted to have lived in fresh or brackish water. It seems entirely possible that the apparent changes in sedimentation rates and microfossil assemblages commonly occur where the change is from non-marine to marine, or vice versa.
Interpretation of microfossil groupings must take into account recent studies by several workers who also have been concerned with the paleoecology of the Council Grove Group and other Lower Permian rocks in Kansas. Extensive review of the factual data and diverging interpretations based upon that data is not pertinent here, but an introduction to the problem can be gained from Elias (1937), Hattin (1957), Imbrie, et al. (1959), Laporte (1962), Elias (1962), and McCrone (1964).
Imbrie and associates have accumulated lithologic, faunal, and paleogeographic data that support the conclusion that during the deposition of the Beattie Limestone turbulence and salinity of water, and rate of terriginous influx were more important causal factors in controlling vertical and lateral distribution of lithology and fossils than were changes in depth of water or distance from shore. They also concluded that the water in which the Beattie Limestone was deposited was shallow--30 feet or less in depth. By inference, it is possible to extend their conclusions to other lithologically and paleontologically similar, generally marine, limestone formations in the Council Grove Group.
On the other hand, Elias (1937, 1962) argued that water depths fluctuated regularly during regressions and transgressions of the shallow seaway, although the rates of withdrawal and flooding of the sea may have been slow or rapid at different times. Consequently, in his opinion, water depth was a primary controlling factor in the distribution of marine organisms and, to a lesser extent, of lithologies. He postulated that maximum depth of water was about 150 to 180 feet and that most fusulinids lived at those depths.
Much of the important data upon which these differing views are based have been collected from studies of limestones, which have been explicitly excluded from this study. Because the limestones are interbedded with the microfossil-bearing shales reported here, and the above interpretations must be taken into account in any further conclusions, I would like to comment on the above conclusions.
The studies cited above raise a question in regard to two aspects of the paleoecological interpretation: what were the general habitat requirements of fusulinids, and what does abundance of marine algae in limestones signify?
There is no doubt, from studies of Pennsylvanian rocks by Harbaugh (1959, 1960) and of the Beattie Limestone by Imbrie, et al. (1959), that fusulinids and algae commonly do occur together, or in closely contiguous beds. From physical evidence of associations of algal breccias, current-bedded algal limestone, and oolitic limestones, much of the algae and associated fusulinids probably accumulated in the zone of greatest wave action. However, the depth of water to which waves are capable of agitating bottom sediments varies widely. Such factors as wind velocity, direction, and duration, and fetch determine the water depth to which wave action is effective. These factors are almost impossible to estimate, and if there were occasional violent storms they could have stirred bottom sediment at depths that were far below the reach of non-storm waves. So, postulation of deposition of sediments within the "zone of active wave action" does not indicate precise estimates of water depths, although it probably does mean that the water was not very deep.
While it is true that all known Recent marine algae live in the photic zone, that zone itself has variable limits. Most of the fossil algae (Osagia, Ottonosia, Anchicodium), which have been found in limestones of the Council Grove Group, presumably were simple green or blue-green algae, although it should be emphasized that their taxonomic affinities with living groups of simple algae are not known. Blue-green marine algae can survive, but probably not flourish, in light intensities as low as 10 foot-candles, which in clear water could mean depths of about 200 meters. However, optimum light intensity for their growth is about 500 foot-candles, which might be characteristic of water 5 to 15 meters deep, depending upon the clarity of the water. [Note: Information on living algae presented here was kindly furnished to me by Dr. Bruce C. Parker, Department of Botany, University of California, Los Angeles.]
The occurrence of a green algae growing on the underside of cobbles in a completely dark or poorly lighted environment, cited by Elias (1962), deserves comment. Many terrestrial species of green algae can live in complete darkness in soils down to depths of a meter or more. These plants are able to cease ordinary photosynthetic activity (although they retain chlorophyll) and, instead, to obtain their food from surrounding organic nutrients and to take on an existence (organotrophic) more like saprophytes or animals. This organotrophic condition is rare in blue-green terrestrial algae--only two or three species are known that adapt to total darkness--although many species, because of their contained pigments, can thrive under conditions of very dim light. No living marine algae has ever been reported to have attained this organotrophic state, although it is possible that there are such algae living today that are as yet unknown to us, or that marine algae might have, at one time or another in the past, adapted to this condition. Considering the optimum conditions in which simple marine algae thrive today, it seems safe to say that they flourish in water depths not in excess of about 50 or 60 feet, probably less, depending upon the latitude, the clearness of water, and the species. Therefore, because of the abundant evidence of flourishing fossil algae in Pennsylvanian and Lower Permian limestones in Kansas, associated physical evidence of wave action, and the occurrence of fusulinids in algal limestones, these rocks are judged to have been deposited in shallow water.
The limits of depth of water for certain phases of the cyclothems of the Big Blue series proposed by Elias (1937) are open to question on the basis of more recent studies. For example, Elias' depth limit of 90 feet between his molluscan and mixed phases is based mainly on the supposition that most living inarticulate brachiopods are found above that depth and most articulates below. His source is Schuchert's (1911) paper; Cooper (1937) has shown that Schuchert's conclusions can no longer be maintained in light of present knowledge of living brachiopod distribution. The maximum depth of water set by Elias for the fusulinid phase is 180 feet. This is based upon the observation that living larger foraminifers most commonly are not found below this depth. However, most of these Recent foraminifers live in clear tropical waters and contain commensal algae (zooxanthellae) that must live in the photic zone (Yonge, 1957). Consequently, in order to validly assume a similar depth restriction for fusulinids, it seems necessary to make the correlative assumption that these extinct protozoans also contained commensal algae. This assumption is not explicitly stated by Elias, nor has it been made by any student of fusulinids to the writer's knowledge. The fusulinids may have lived at depths considerably shallower or deeper than those proposed by Elias. It can be postulated that those species of fusulinids that are characteristically found in association with fossil algae may have contained zooxanthellae in their cytoplasm and because of this were restricted to the photic zone.
One must conclude that absolute depth values are almost meaningless in attempting to reconstruct the paleoecology of fossils as old as those found in early Permian beds. The accumulated data seems to favor relatively shallow water deposition for most of the marine rocks in the Council Grove Group, but whether the water reached maximum depths of 10, 30, 60, or 100 feet is open to question, and it can only be hoped that further careful research may provide some more concrete answers.
Finally, it is judged that the microfaunal marine assemblages reported here from the Council Grove Group do not provide unequivocal confirmation or refutation of either of the two postulated sets of primary controls discussed above.
The Globivalvulina-Bairdia assemblage, or portions thereof, seemingly is ubiquitous in marine shales of the Council Grove Group. The Healdia-?Coryellites and the fusulinid-Textularia assemblages are most common in dark gray shales, especially in the Hughes Creek Shale and in dark gray shale of Sample 59 of the Eiss Limestone. The dark color of these shales probably is due to the presence of relatively large amounts of organic carbon that was not destroyed by bacteria or other scavengers, as was the carbon in the lighter gray or brown shales of the Group. Presumably, turbulence of water, differences in amount and kind of zoo- and phytoplankton, and rate of sedimentation, rather than depth of water, could have been the controlling factors. Dark shales in the Hughes Creek contain the bulk of fusulinids reported in this study. Under Elias' hypothesis, the Hughes Creek Shale, then, would represent the sediment deposited under the deepest water conditions among the shales studied.
The Deckerella-Kirkbya-Knightina assemblage is characteristic of brown or light-gray, calcareous shale, as in samples 112 to 114 in the upper Neva Limestone Member, where this assemblage occurs in association with many elements of the Globivalvulina assemblage, Presumably, light-colored calcareous shales of this kind represent a different set of ecologic conditions than those that existed during deposition of dark gray shales.
Further study of microfossil assemblages in shales of other sections of the Council Grove Group, and of other groups within the Lower Permian and Upper Pennsylvanian of Kansas and adjacent states, would shed much light on the nature of microfossil assemblages. Such studies could be fruitfully combined with a study of the clay mineralogy and trace element composition of the shales, and of pollen-spore assemblages that might be obtainable by other methods of sample preparation.
Bandy, O. L., 1953, Ecology and paleoecology of some California Foraminifera. Pt. 1. The frequency distribution of Recent Foraminifera off California: Jour. Paleontology, v. 27, p. 161-182.
Cooper, C. L., 1946, Pennsylvanian ostracodes of Illinois: Illinois State Geol. Survey, Bull., v. 70, p. 9-123.
Cooper, G. A., 1937, Brachiopod ecology and paleoecology: Natl. Research Council (1936-1937), Rept. Committee on Paleoecology, p. 26-53.
Cordell, R. J., 1952, Ostracodes from the Upper Pennsylvanian of Missouri: Pt. 1. The family Bairdiidae: Jour. Paleontology, v. 26, p. 74-112.
Cushman, J. A., 1911, A Monograph of the Foraminifera of the North Pacific Ocean: U.S. Natl. Museum, Bull. 71, 596 p.
Cushman, J. A., and Valentine, W. W., 1930, Shallow-water Foraminifera from the Channel Islands of Southern California: Contrib. Dept. Geol., Stanford Univ., v. 1, no. 1, p. 1-51.
Echols, D. J., and Creath, W. B., 1959, Survey of Mississippian, Pennsylvanian, and Permian Ostracoda recorded in the United States: Micropaleontology, v. 5, p. 389-414.
Elias, M. K., 1937, Depth of deposition of the Big Blue (Late Paleozoic) sediments in Kansas: Geol. Soc. America, Bull., v. 48, p. 403-432.
Elias, M. K., 1962, Comments on recent paleoecological studies of Late Paleozoic rocks in Kansas: Kansas Geol. Soc., Guidebook, 27th. Field Conf., p. 106-114.
Fager, E. W., 1957, Determination and analysis of recurrent groups: Ecology, v. 38, p. 586-595.
Fager, E. W., and McGowan, J. A., 1963, Zooplankton species groups in the North Pacific: Science, v. 140, no. 3566, p. 453-460.
Grekoff, N., 1956, Guide Pratique pour la détermination des Ostrocades post-Paléozoïques: Inst. Français du Pétrole, Div. Sédimentologie, Protat Frères, Mason, 95 p.
Gunter, Gordon, 1947, Paleoecological import of certain relationships of marine animals to salinity: Jour. Paleontology, v. 21, p. 77-79.
Gunter, Gordon, 1947a, Extended remarks on relationships of marine animals to salinity: Jour. Paleontology, v. 21, p. 498500.
Harbaugh, J. W., 1959, Marine bank development in Plattsburg Limestone (Pennsylvanian), Neodesha-Fredonia area, Kansas: Kansas Geol. Survey, Bull. 134, pt. 5, p. 289-331. [available online]
Harbaugh, J. W., 1960, Petrology of marine bank limestones of Lansing Group (Pennsylvanian), southeast Kansas: Kansas Geol. Survey, Bull. 142, pt. 5, p. 189-234. [available online]
Hattin, D. E., 1957, Depositional environment of the Wreford megacyclothem (Lower Permian) of Kansas: Kansas Geol. Survey, Bull. 124, p. 1-150. [available online]
Imbrie, John, 1955, Quantitative lithofacies and biofacies study of the Florena Shale (Permian) of Kansas: Am. Assoc. Petroleum Geologists Bull., v. 39, p. 649-670.
Imbrie, John, Laporte, Leo, and Merriam, D. F., 1959, Beattie Limestone facies and their bearing on cyclical sedimentation theory: Kansas Geol. Soc., Guidebook, 24th Field Conf., South-central Kansas, p. 69-78.
Ireland, H. A., 1956, Upper Pennsylvanian arenaceous Foraminifera from Kansas: Jour. Paleontology, v. 30, p. 831864.
Johnson, R. G., 1962, Interspecific associations in Pennsylvanian fossil assemblages: Jour. Geology, v. 70, p. 32-55.
Kellett, Betty, 1933, Ostracodes of the upper Pennsylvanian and the lower Permian strata of Kansas, I. The Aparchitidae, Beyrichildae, Glyptopleuridae, Kloedenellidae, Kirkbyidae, and Youngiellidae: Jour. Paleontology, v. 7, p. 59-108.
Kellett, Betty, 1935, Ostracodes of the upper Pennsylvanian and lower Permian strata of Kansas, III. Bairdiidae (concluded), Cytherellidae, Cypridinidae, Entomoconchidae, Cytheridae, and Cypridae: Jour. Paleontology, v. 9, p. 132-166.
Kellett, Betty, 1936, Carboniferous ostracodes: Jour. Paleontology, v. 10, p. 769-784.
Kremp, Gerhard, 1952, Sporen-Vergesellschaftungen und Mikrofaunen-Horizonte im Ruhrkarbon: Cong. Avanc. Études stratig. et géol. Carbonifère 3rd, C.R., t. 1, p. 347-357.
Lane, N. G., 1958, Environment of deposition of the Grenola Limestone (Lower Permian) in southern Kansas: Kansas Geol. Survey, Bull. 130, pt. 3, p. 117-164. [available online]
Laporte, L. F., 1962, Paleoecology of the Cottonwood Limestone (Permian) northern Midcontinent: Geol. Soc. America, Bull., v. 73, p. 521-544.
McCrone, A. W., 1964, Paleoecology and biostratigraphy of the Red Eagle cyclothem (Lower Permian) in Kansas: Kansas Geol. Survey, Bull. 164, 114 p. [available online]
Moore, R. C., Frye, J. C., Jewett, J. M., Lee, Wallace, and O'Connor, H. G., 1951, The Kansas rock column: Kansas Geol. Survey, Bull. 89, p. 1-132. [available online]
Peck, R. E., 1934, The North American Trochiliscids, Paleozoic Chatrophyta: Jour. Paleontology, v. 8, p. 83-119.
Peck, R. E., 1953, Fossil Charophytes: Botanical Review, v. 19, p. 209-227.
Peck, R. E., and Eyer, J. A., 1963, Pennsylvanian, Permian, and Triassic Charophyta of North America: Jour. Paleontology, v. 37, p. 835-844, pl. 100-101.
Schuchert, Charles, 1911, Paleogeographic and geologic significance of Recent Brachiopoda: Geol. Soc. America, Bull., v. 22, p. 258-275.
Scott, H. W., 1944, Permian and Pennsylvanian fresh-water ostracodes: Jour. Paleontology, v. 18, p. 141-147.
Segerstråle, S. G., 1957, Baltic Sea: Geol. Soc. America, Mem. 67, v. 1, p. 751-800.
Shaver, R. H., 1958, A study of Pseudobythocypris pediformis, a new name for an old ostracode: American Midland Nat., v. 59, p. 120-137.
Thompson, M. L., 1954, American Wolfcampian fusulinids: Univ. Kansas, Paleont. Contr., Protozoa, art. 5, 225 p. [available online]
Van den Bold, W. A., 1958, Distribution of fresh-water ostracodes in Trinidad: Micropaleontology, v. 4, p. 71-74.
Wagner, C. W., 1957, Sur les ostracodes du Quaternaire récent des Pays-Bas et leur utilisation dans l'étude géologique des dépôts Holocène: Mouton & Co., 's-Gravenhage, 158 p.
Yonge, C. M., 1957, Interrelations of organisms, C. Symbiosis: Geol. Soc. America, Mem. 67, v. 1, p. 429-442.
Kansas Geological Survey, Geology
Placed on web Jan. 7, 2008; originally published Dec. 1964.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/170_5/index.html