Kansas Geological Survey, Bulletin 102, Part 2, originally published in 1953
Originally published in 1953 as Kansas Geological Survey Bulletin 102, Part 2. This is, in general, the original text as published. The information has not been updated.
Surficial characters and internal structures of a new species of ammonoid opercula, Spinaptychus sternbergi, discovered in the Cretaceous Niobrara formation of western Kansas, are described and illustrated. The Kansas specimens are better preserved and larger than any previously described species of the genus and represent the first record of the genus from North America. Orientation of growth lamellae leads to the conclusion that Spinaptychus was not a strictly external structure, because the outer margins of its exterior surface at least evidently were covered by mantle tissues.
Mr. George F. Sternberg, of the Fort Hays Kansas State College Museum, during years of collecting by himself and others, has obtained a number of ammonoid opercula from the Cretaceous Niobrara formation of western Kansas. Most of these aptychi are fragmentary, but two pairs are nearly complete. All belong to the genus Spinaptychus Trauth, previously recorded only from the Cretaceous of England and of Palestine. The material which Mr. Sternberg kindly turned over to us for study consists of two nearly complete pairs of aptychi, one fragmentary valve of extraordinarily large size, and numerous fragments.
Genus Spinaptychus Trauth, 1927
Records of the genus--The genus Spinaptychus was established by Trauth (1927), with Aptychus spinosus Cox (1926) as its type. At the time of Trauth's publication, and seemingly to the present day, the type species is the only one referred to Spinaptychus. The genus is distinctively characterized by its ornamentation, the exterior surface being covered with tubercles which bear a pit at their apex.
Cox's specimens came from the Micraster coranguinum zone (Santonian, lower Senonian) of Kent, England. Cox illustrated an exterior view of a rather incomplete pair of valves, another likewise fragmentary exterior of a single valve, and the interior of a nearly complete valve. He also described, but did not illustrate, the internal microstructure.
In 1929, the genus Spinaptychus was recorded by Picard from the Upper Cretaceous (Senonian, probably Campanian) of Palestine, near Jericho. The single specimen, a fragment showing the exterior surface and mold of the interior, was referred to Aptychus (Spinaptychus) spinosus Cox. The aptychus is covered with tubercles, but two possibly notable differences from A. spinosus are mentioned: (1) the pore at the top of the tubercle is elliptical rather than circular and (2) the tubercles are arranged rather loosely in concentric rows parallel to the lateral margin.
To our knowledge, these are the only documented occurrences of Spinaptychus. Therefore, it is especially interesting to find a distinctive aptychus of this type in the Smoky Hill chalk member of the Niobrara formation of the Cretaceous interior seaway of North America.
General morphology--Like other aptychi, Spinaptychus occurs in bilaterally paired valves, which are thought to have served as opercula for the shell of some ammonoid. Each valve is slightly arched. The concave surface, which is marked only by growth lines (Pl. 2B), is interpreted to be the inner surface of the operculum attached to the flesh of the mantle.
The outer convex surface of the aptychus shows only faint growth lines, but is studded with tubercles, most of which bear a craterlike pit (Pl. 1B and D). The outline of each valve is roughly triangular. The short side of the triangle forms part of the dorsal side of the aptychus, which presumably fitted against the dorsal margin of the aperture. The inner, or harmonic, margins are relatively straight but sufficiently curved so that the valves, when laid out flat, are in contact along less than half of their length, gaping rather widely at the ventral end (Pl. 1B). The lateral and ventral margins form a continuous sweeping curve. The growth lines on the inner surface of the valves show that growth of the aptychus took place along the lateral and ventral margins only.
Plate 1--Spinaptychus sternbergi, n. sp., from the Niobrara formation of Logan and Gove Counties, Kansas. A, Paratype (Mus. no. 2683). Inner view of harmonic margin of left valve showing the nature of the harmonic flange (X 0.75). B, Holotype (Mus. no. 2022). Outer view (X 0.45). C, Paratype (Mus. no. 2906). Projection print of transverse thin section of mature region, cut normal to growth lines, showing microstructure of shell. Tone of print is reversed. Arrow points to ventrolateral margin (X 8.25). D, Holotype (Mus. no. 2022). Enlarged portion of mature region of right valve near lateral margin (X 1.2). [A higher resolution version is available as an Acrobat PDF file (2.1 MB).]
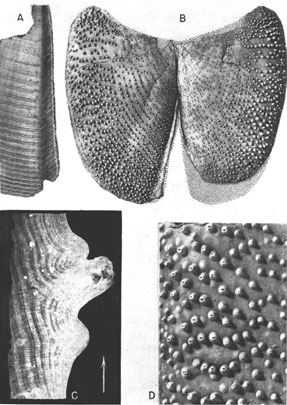
Along the harmonic margins is an outwardly convex and inwardly concave flange, which is narrow or absent in the dorsal (juvenile) region but expands progressively toward the venter (Pls. 1A, 2B).
Although Cox reported that his specimens were considerably altered by recrystallization, he distinguished three layers: (1) an inner dense layer; (2) a thick middle layer which had lost all structure owing to recrystallization; and (3) an outer laminar layer. The spines were described as hollow down to the level of the middle layer. Unfortunately, Cox published no illustrations of the internal structure. Cox's conception of internal microstructure is at variance with that observed in the Kansas specimens described below. These are similar in general surface character to Spinaptychus spinosus (Cox) but differ in outline, size, and pitting of tubercles. Clearly, our specimens represent a different species, which is here named for Mr. George F. Sternberg, the collector.
Plate 2--Spinaptychus sternbergi, n. sp., from the Niobrara formation of western Kansas. A, B, Paratype (Mus. no. 2685). A, Outer view of right and left halves of an immature specimen. B, Interior of right valve. Both X 0.65. C, Paratype (Mus. no. 2907). Projection print of transverse thin section, normal to growth lines, showing canals in tubercles. Tone reversed. Arrow points to ventrolateral margin (X 7.5). D, Paratype (Mus. no. 2908). Projection print of transverse thin section of valve, through harmonic flange (at right), parallel to growth lines. Tone of print is reversed (X 4.5). [A higher resolution version is available as an Acrobat PDF file (2 MB).]
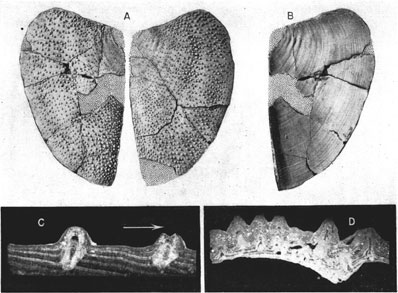
Spinaptychus sternbergi, n. sp.
Outline--Measurements of our specimens are given in Table 1. The earlier and intermediate growth stages show a distinctly triangular outline, the ventral and lateral margins forming a continuous sweep to a rather sharp mid-ventral point (Pl. 2A and B). Later stages are somewhat more quadrate in outline (Pl. 1B). The width of our smaller specimens is 65 percent of the length, and the width of a larger specimen (holotype) equals 67 percent of the length. The angle between the dorsal and harmonic margins is about 104 degrees.
Table 1--Table of comparative data.
| Length (dorsoventral) mm |
Width mm |
Inflation mm |
Thickness (maximum) of shell mm |
Apical angle |
Angle of taper of harmonic groove |
Width of largest spines mm |
Height of largest spines mm |
Unpitted spines |
Shape of pits in spines |
|
|---|---|---|---|---|---|---|---|---|---|---|
| S. spinosus (Cox) | 48-65 | 38-52 | 9 | thin | 90° | 5° | 2 1/2 | 3 | ? | circ. |
| S. spinosus, Picard | 55 | 48 | ? | ? | 80° | ? | 1 1/2 | 1 | ? | ellipt. |
| S. sternbergi, n.sp. | ||||||||||
| Holotype (2022) | 150 | 82 | 11 | ? | ? | 4° | 2 | 2 | yes | ellipt. |
| Paratype (2685) | 90 | 58 | 5 | 1 | 104° | 5° | 1 | 1 | yes | ellipt. |
| Paratype (2683) | 170 | 100-105 | 10 | 2 1/2 | 104° | 4° | 2 1/2 | 2 1/2 | yes | ellipt. |
Inner surface--The inner surface of the valves is dull, partly discolored by carbonaceous matter in one of the specimens, and distinctly marked by growth lamellae. In the early stages these growth lamellae are broad and are separated by rather faint lines. The later growth lamellae, found near the margins of larger specimens, are narrower but are sharply separated, so that the periphery takes on a rugose appearance. The inner surface bears a well-defined harmonic flange (Pls. 1A, 2D).
Outer surface--The outer surface is dense and lacks well-defined growth lines. It bears numerous tubercles generally distributed at random and varying in degrees of spacing. The tubercles on the juvenile portion near the corner between the dorsal and harmonic margins are nearly microscopic and are widely spaced. They become more closely spaced and larger toward the periphery, the largest tubercles reaching a height of 2.5 mm. Along the peripheral margins the tubercles are small, for these are not yet fully formed. Actual size and spacing vary from one specimen to another. Only one, the holotype, shows a concentric arrangement of tubercles in the immature parts of the valves. In more mature regions of the aptychi, the large pitted tubercles are not alone, for between them may lie irregularly scattered domelike mounds and unpitted tubercles of intermediate size. The more prominent tubercles in any region of the aptychus contain an elliptical pit at their tip, commonly surrounded by an elevated rim (Pl. 1D, Fig. 1A-C). The long axes of the pits are radially oriented, that is, normal to the growth lines. The region surrounding the pit seems to have been somewhat worn away as though the tips of the spines suffered abrasion.
Microstructure--Thin sections of our well-preserved specimens (Pls. 1B; 2C and D) show a structure at variance with that described by Cox. The Kansas aptychi are essentially uniform in structure, composed of growth bands or laminae which were laid down successfully at the ventral and lateral margins. Each of these laminae slants peripherally from its termination at the outer surface to the place of disappearance on the inner surface of the aptychus (Pls. 1C, 2C; Fig. 1C). Each lamina terminates at the outer surface in a wedge-edge, which is hardly discernible in exterior view of the surface; it increases in thickness inwardly, so as to form a broad band on the inner surface of the aptychus. The laminae are alternately clear and turbid; the clear ones form ridges on the inner surface, whereas the turbid ones form the intervening depressions. Toward the exterior, the contrast between these laminae is lost because of prevailing turbidity of the shell structure. Both types of laminae are subdivided into thinner lamellae, which are crossed at right angles by the calcite fibers composing them. The orientation of growth laminae leaves no doubt that they were laid down on the outer, convex side of the valves.
Cross sections of the tubercles show that they are produced by upward doming of the laminae (Pl. 2C). As is also evident from surface inspection, newly formed tubercles at the edge of the aptychus are relatively small. Growth of the tubercles occurs by superposition of successive laminae.
Cox described the spines of his specimens as hollow down to the "middle layer" of the aptychus--that is, deep into the valve. The depth to which the pits extend in the tubercles of our Kansas specimens is not certain. We cut numerous sections in an effort to determine this matter and concluded that an open tube is probably restricted to the upper parts of the large tubercles (Pls. 1C, 2C and D; Fig. 1A-C). Sections needed for solution of this problem must be cut parallel to the axis of the spine. Since these axes generally are inclined and may be curved, one is likely to cut them obliquely. We believe that Plate 1C, Plate 2C (right tubercle), and Figure 1A and B are essentially parallel to the axes of some of the tubercles shown.
Figure 1--Thin sections and restorations of aptychi. Arrows indicate direction toward ventrolateral margin. A, Mus. no. 2684, section across a gerontic aptychus near ventrolateral margin (X 15). Large tubercle showing canal in upper part and structureless calcite crossed by occasional growth laminae in middle and lower parts. B, Mus. no. 2684, thin section of tubercles of a gerontic aptychus close to ventrolateral margin which in this specimen is curled under (X 15). C, Mus. no. 2907a, radial section cut to the edge of the ventrolateral margin of an immature specimen. Note imbrication of growth lamellae and cross section of nonperforate blisterlike tubercle (X 24). D, Diagrammatic dorsoventral section of aptychus and restored tissues. Tissues of attachment are crosshatched, secreting tissue stippled. E, Restoration, head extended. F, Restoration, head withdrawn.
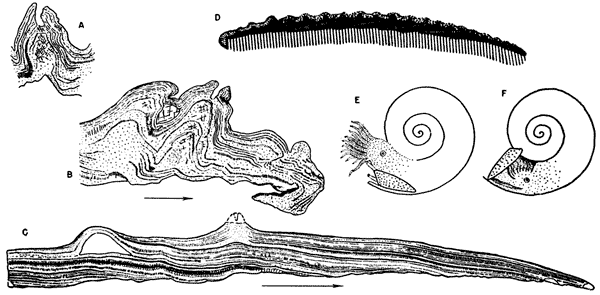
The small spines show a core of clear or cloudy calcite, crossed at intervals by single growth laminae. The structure suggests a pulpy, poorly calcified, perhaps vesicular core which became recrystallized and filled during diagenesis. Large tubercles are similarly built in the lower part, but are topped by a nipplelike protuberance, which contains an open canal (Fig. 1A).
The low nonperforate tubercles are revealed as blisterlike structures filled with coarsely crystalline calcite of secondary origin (Fig. 1C).
To sum up differences between observations of Cox on the English Spinaptychus and those made by us on the Kansas specimens. (1) We have not found a threefold division of the shell, but rather a uniform structure of imbricate laminae. (2) Furthermore, we are not convinced that the pores in tubercles of our specimens extended as tubes deep into the aptychus in the manner described by Cox. The question rises as to whether these differences may be sufficient to warrant generic distinction. We have not seen Cox's sections, and, lacking figures of them, we are doubtful as to actuality of difference in original shell structure of the British and American specimens. If the mid-portion of the aptychus sections illustrated in Plates 1 and 2 had been recrystallized, it is probable that we would have described our material as three-layered. Therefore, until proved otherwise, it seems reasonable to interpret major microstructure differences between Cox's description of Spinaptychus and our own observations of presumed Spinaptychus from Kansas as attributable to differences in preservation.
Although our specimens are judged to be congeneric with those described by Cox and by Picard, they differ from Cox's specimens in size, proportions, and the nature of the tubercles, and from Picard's specimens at least in size and shape.
As shown by their dimensions (Table 1), the Kansas specimens greatly exceed the others in size. They are narrower (laterally) and longer (dorsoventrally) than the form described by Cox and differ from both the British and Palestinian material in the nature of the dorsal margin. Picard's and Cox's forms show an angle of approximately 90 degrees between the harmonic and dorsal margins, whereas our specimens show an angle of 104 degrees. The dorsal margins of their specimens form a nearly straight line; the dorsal margins of ours form a concave embayment, designed to fit against a more deeply impressed whorl.
The tubercles of our two paratypes and of all the available fragments show a random distribution, as do those of Cox's species. The holotype of Spinaptychus sternbergi clearly differs from the type of S. spinosus from England but resembles Picard's specimen in having the tubercles of intermediate growth stages aligned parallel to the lateral and ventral margins. This alignment is not present in early stages and is lost in the adult. In the absence of other distinguishing characters, we believe that such alignment may be a matter of individual variation.
Our specimens agree with the Palestinian one and differ from the British form (as described by Cox) in possessing elliptical rather than circular craters. Neither Cox nor Picard mentions the presence of nonperforate tubercles, which are abundant in our form.
It may be concluded, therefore, that our specimens are specifically distinct from the British and Palestinian specimens referred to Spinaptychus spinosus (Cox). The question may be raised as to whether all our specimens from Kansas are conspecific. Despite some variation in shape and ornamentation, we believe that they do belong to the same species. Size, distribution, and arrangement of tubercles may vary widely on a single specimen; subquadrate shape and the development of a great harmonic flange are clearly expressions of ontogeny.
Orientation and growth of Spinaptychus--Descriptions of aptychi and attempts at explaining their zoological affinities and function have been made by many writers since the beginning of paleontology. Trauth (1927, 1928, 1931) summarized everything known about aptychi up to that time and added a wealth of new information. Trauth's conclusions on orientation and growth of aptychi are summarized below.
Fortunate finds of aptychi in position occupied during life have proved beyond doubt that they are skeletal structures of ammonoids which served as opercula to close the aperture. They have further substantiated the conclusion, previously reached on the basis of aptychus structure, that the convex side of the valves is the outer one and the concave side the inner one.
It has become so customary to interpret ammonoids by analogy with the modern Nautilus that one is tempted, at first glance, to interpret the aptychus as homologous to the Nautilus hood, the tough mass of tissue lying above the head and largely closing the aperture when the head is withdrawn. Trauth and others have shown that while the aptychus served the function of the nautiloid hood, it is not a morphological equivalent of this structure, for it was placed differently. The majority of aptychi found in association with shells are located within the living chamber, lying along its ventral margin. The dorsal margin of these aptychi points forward and the ventral margin toward the rear. The external side is down, the internal side up. This orientation is so common and constant that it has become known as the normal position. It led some early observers to the erroneous conclusion that the aptychi functioned as protective shields over the female reproductive organs.
It seems only logical to conclude that the aptychi were carried normally in this position. When the head was retracted, the valves slid forward and upward to close the aperture. When the head was extended, the valves of the aptychus were presumably retracted below into a ventral mantle pocket.
This hypothesis is supported by other lines of evidence. It explains why the majority of ammonoid opercula are of the two-valved aptychus type rather than single shields (anaptychi), for in compressed shells a two-valved construction was necessary to permit folding of the operculum, which otherwise could not have been accommodated in the narrow ventral part of the living chamber. The hypothesis is further supported by observation that in many aptychi shell matter was added on the external surface of the valves (as in Spinaptychus). If the valves had been located in the position of the Nautilus hood, such deposits could only have been formed by a special mantle flap over their exterior. In the position envisaged by Trauth , this deposition could be accomplished by the ventral tissue lining of the pocket in which the aptychus rested when withdrawn.
Affinities--The aptychi here described have not been found attached to, or in close association with, ammonoid conchs. We are not able to identify with certainty, therefore, the species or genus of ammonoid conch for which Spinaptychus sternbergi served as an operculum. Whatever its identity, the ammonoid which bore this aptychus must have been very large, and the cross section of its whorls must have been something between heart-shaped and subquadrate. Cox has suggested that Spinaptychus may belong to the so-called Mortoniceras, which subsequently has been renamed Texanites Spath.
Ammonoid conchs have been reported from the Niobrara formation (Morrow, 1935), but specimens evidently are rare and in this formation are found preserved only as molds. Morrow lists a questionable Pachydiscus, and the University of Kansas collections contain a Texanites from the Niobrara formation of Trego and Ellis Counties, Kansas. The latter specimen has a whorl cross section which seems to be an excellent fit for Spinaptychus sternbergi. Accordingly, we concur with Cox in judging that in all probability Spinaptychus constitutes the operculum of "Mortoniceras" (=Texanites).
The discovery of more aptychi than conchs in the chalk is almost certainly a matter of preservation. The aptychi, composed of calcite, are among the best preserved fossil remains found in the chalk, whereas the aragonitic conchs were dissolved during diagenesis and the resulting molds have been partly crushed or obliterated during compaction of the chalk. The conchs are therefore more obscure and less commonly collected.
The relationships of Spinaptychus to other types of aptychi are not clear. Little is known about the structure of Cretaceous aptychi, and most comparisons therefore must be made with the much better known Jurassic forms, none of which closely resembles Spinaptychus. The pitted tubercles covering the outer surface of Spinaptychus are unique. The black layer on the concave inner surface of the valves is the remainder of a conchiolinic "inner layer," a structure characteristic of the Jurassic cornaptychi (belonging to the harpoceroids). In other Jurassic calcareous ammonoid valves, this thin layer is calcified. Unlike Spinaptychus, the main layer of the cornaptychi is cellular. The imbrication of growth layers in our specimens suggests the structure of the Jurassic laevaptychi, but in these, again, the layer is cellular.
It seems, therefore, that Spinaptychus, while unique in ornamentation, is conservative or even primitive in terms of basic shell structure. Trauth's studies indicate that this may be true for other Cretaceous aptychi as well.
All types are deposited in the Fort Hays Kansas State College Museum, Fort Hays, Kansas.
Holotype--Mus. no. 2022. Horizon: upper Smoky Hill chalk member, Niobrara formation. Locality: about 2 miles northeast of Pyramid rocks, Gove County, Kansas. Collector: George F. Sternberg in about 1942.
Paratypes--Mus. no. 2083. Horizon: upper Smoky Hill chalk member, Niobrara formation. Locality: about 2 miles northeast of Pyramid rocks, Gove County, Kansas. Collector: George F. Sternberg.
Mus. no. 2296. Horizon: upper Smoky Hill chalk member, Niobrara formation. Locality: about 4 miles southeast of Elkader, Logan County, Kansas. Collector: Robert Taylor in fall of 1949.
Mus. no. 2683. Horizon: upper Smoky Hill chalk member, Niobrara formation. Locality: 3 miles southeast of Elkader, Logan County, Kansas. Collector: Robert Taylor on October 8, 1949.
Mus. no. 2685. Horizon: upper Smoky Hill chalk member, Niobrara formation. Locality: about 4 miles southwest of Castle Rock, Gove County, Kansas. Collector: George F. Sternberg.
Mus. no. 2906. Horizon: Niobrara formation. Locality: Logan and Gove Counties, Kansas.
Mus. no. 2907. Same as no. 2083.
Mus. no. 2907a. Same as no 2083.
Mus. no. 2908. Same as no. 2683.
Cox, L. R. (1926) Aptychus spinosus, sp. n., from the upper Chalk: Ann. and Mag. Nat. Hist., London, ser. 9, vol. 17, pp. 573-580.
Morrow, A. L. (1935) Cephalopods from the Upper Cretaceous of Kansas: Jour. Paleontology, vol. 9, no. 6, pp. 463-473.
Picard, Leo (1929) On Upper Cretaceous (chiefly Maestrichtian) Ammonoidea from Palestine: Ann. and Mag. Nat. Hist., London, ser. 10, vol. 3, pp. 433-456.
Trauth, F. (1927) Aptychenstudien. I. Über die Aptychen im Allgemeinen: Ann. d. Naturh. Mus., Wien, vol. 41, pp. 170-259.
Trauth, F. Aptychenstudien. II. Die Aptychen der Oberkreide: Ann. d. Naturh. Mus., Wien, vol. 42, pp. 121-193.
Trauth, F. Aptychenstudien. VI-VII: Ann. d. Naturh. Mus., Wien, vol. 45, pp. 18-137.
Kansas Geological Survey, Geology
Placed on web Aug. 8, 2007; originally published in June, 1953.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/102_2/index.html