Kansas Geological Survey, Current Research in Earth Sciences, Bulletin 248, part 1
Prev Page--Results || Next Page--Conclusions
![]()
![]()
![]()
Kansas Geological Survey, Current Research in Earth Sciences, Bulletin 248, part 1
Prev Page--Results ||
Next Page--Conclusions
![]()
Climatic factors (precipitation and PDSI) and residual tree-ring widths display a close relationship, such that the initiation of drought (1988 and 1991) results in major reduction of tree-ring growth (see fig. 24). In contrast, the initiation of drought had minimal effect on Landsat NDVI values for the study forests (see fig. 17). In fact, the July 1988 bottomland forest experienced a slight increase in its NDVI value compared to 1987. NDVI values reached their minima in 1990 for all categories of analysis--whole-study scene, upland forest, and bottomland forest. NDVI values declined from 1989 to 1990, even though precipitation and tree-ring growth increased substantially in 1990. A similar, but smaller decline in NDVI values occurred in 1992, a year after the beginning of drought in 1991. Again, this change in NDVI was out of phase with tree-ring growth.
Landsat NDVI values for northeastern Kansas do not reflect climatic conditions for the current growing season nor for the preceding 12-month period. Instead, changes in NDVI values of forests lag one to two years behind significant climatic events. Furthermore, changes in NDVI values take place a year after variations in tree-ring growth for the upland forest. These results are consistent with earlier preliminary findings for the study area (Aber et al., 1998). Similar NDVI lag effects have been noted in other forest studies in Kansas (Wang et al., 2001) and in East Africa (Davenport and Nicholson, 1990). Thus, satellite-derived NDVI values should be used cautiously when interpreting impacts of current climate on growth of deciduous forest in similar settings. The NDVI values most likely represent climatic events of the previous one or two years.
Landsat NDVI values are a reflection of the forest canopy, namely leaf area index (LAI). The growth of leaves in deciduous forest is based on food stored in tree limbs and twigs. The amount of food available for leaf growth in the spring is a function of favorable or unfavorable climatic conditions of the previous growing season. Thus, the forest canopy in a given year is a reflection of antecedent climatic events. In contrast, growth of wood in the body of the tree depends primarily on moisture and nutrients available to the tree roots (Fritts, 1976). This involves a combination of shallow soil moisture and deeper ground water. The former often depends on recent precipitation events during the winter and early portion of the growing season, whereas the latter responds to multi-year climatic fluctuations. Thus, tree-ring growth may have a rather complicated relationship to short- and long-term climatic events.
The upland study forest, for which tree-ring data were collected, depends entirely on soil moisture to support tree growth. Moderate to steep slopes promote rapid runoff of precipitation, and ground water is far below the zone of tree roots. In this situation, growth of tree rings in oak species is sensitive to climatic events, especially drought episodes (Fritts, 1976). It is not surprising to find close correlation between residual tree-ring width, precipitation, and PDSI on an annual basis. The bottomland study forest, in contrast, depends almost entirely on ground water, which lies only a few meters below the surface in most years. Growth of cottonwood tree rings has little connection to annual climatic events. However, long-lasting droughts could impact bottomland forest growth because of lower ground-water levels.
The bottomland forest experienced an anomalous increase in vegetation cover in July 1988, as indicated by NDVI values, image differencing, and the band 5:4 ratio. This increase in vegetation cover is contrary to normal expectations for drought effects and is opposite to the slight reduction in vegetation cover in the upland forest. Field observations during the drought of 2000 provide a possible explanation. During the first year of a drought episode, the floodplain surface dries up; small water bodies in meander scrolls disappear, and wetland soil begins to lose water. This allows grass and herbaceous plants to rapidly spread over the dry hollows and potholes on the forest floor. Meanwhile, ground water remains relatively close to the surface and available to tree roots. The net result is an increase in vegetation cover within the bottomland forest. Continuation of a drought during subsequent years, however, leads to soil-moisture deficit and lowered ground water, with a reduction in understory vegetation and reduced tree growth. This appears to be what happened during the drought of 1988-1989 in the bottomland forest, when NDVI values first increased slightly in 1988 and then declined abruptly in 1989.
Changes in vegetation occurred mainly along the edges of both the upland and bottomland study forests, as demonstrated by image differencing and principal-component analysis (figs. 27 and 28). Boundary or edge effects are well known in ecological studies of vegetation. In the case of forests, the edges are more exposed to microclimatic impact of insolation, wind stress, and temperature, which affect evapotranspiration and, thus, soil moisture. The forest interior is more protected from these microclimatic factors, which helps to preserve soil moisture. Another factor that undoubtedly affected the bottomland forest was flooding of the Missouri River in 1993. Erosion and deposition of sediment caused permanent changes in soil and topography of the floodplain. It is not surprising that the greatest changes in the bottomland forest took place next to the Missouri River channel, along the northern margin of the forest (fig. 29). Changes also happened on the western margin of the forest, where it meets the levee around the airfield.
Fig. 27--Digital orthophotograph showing the western margin of the upland study forest near Bell Point. This edge experienced significant decline during the period of study. Compare with fig. 18.

Fig. 28--Digital orthophotograph showing the upland maple-basswood forest along the Missouri valley bluff. The western margin of this forest experienced significant decline during the period of study. Compare with fig. 18.
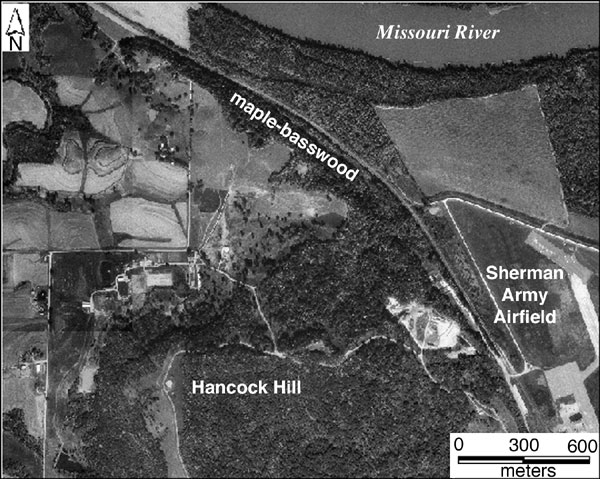
Fig. 29--Digital orthophotograph showing the bottomland study forest. The northern margin of this forest experienced significant decline during the period of study. Changes also took place along the western margin next to the levee around the airfield. Compare with fig. 19.
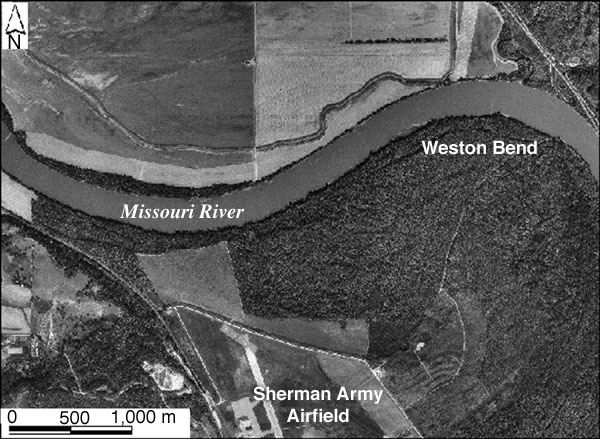
Overall, NDVI values declined from 1987 to 1997 for all areas studied: upland and bottomland forests as well as whole-study scenes (see tables 2, 3). Human impacts--such as mowing, clearing, and construction--could be a local factor, particularly along forest margins. However, this seems an unlikely explanation for long-term decline in whole-study scene NDVI values. A more-probable explanation is connected to long-lasting climatic trends, in which the average yearly precipitation of 74 cm (29 in) for the period of study is well below the long-term average of 90 cm (35 in). Following the severe drought of 1988-1989, forest cover experienced partial recovery. This recovery was interrupted by a minor drought in 1991-1992 and major flooding in 1993. As of 1997, forest cover had not quite recovered to the level of 1987. Given the drought of 2000, it seems likely that another decline in forest NDVI may take place in 2001.
Residual values for the TM band 5:4 ratio showed different fluctuations from the NDVI values. Residual 5:4 values increased markedly in 1989 and 1994 for both study forests (see fig. 23). The 1989 increase corresponds with the lowest mean annual precipitation and most negative PDSI value in the study period. The 1994 increase, however, is consistent with reduced tree-ring width and principal-component analysis, which suggest that both forests suffered limiting growth factors. The preceding year, 1993, experienced highest mean annual precipitation, major flooding, and lowest mean annual temperature during the period of study. On this basis, it appears the residual values for TM band 5:4 ratios may reflect current conditions as well as climatic events of the past year. The climatic significance of changes in the TM band 5:4 ratio is difficult to evaluate, as either drought or flood appears to increase residual values. However, the band 5:4 ratio is consisently higher for hardwood (upland) forest than for the cottonwood (bottomland) forest (see fig. 22). As these forest types are visually indistinguishable on the Landsat images, this ratio difference could be useful in separating and classifying similar types of forests elsewhere.
The photographs taken by KAP over the upland forest show a rough canopy with many shadows at the scale of meters (see fig. 25). These shadows would reduce values for reflected solar energy in the visible, near-infrared, and mid-infrared spectra. Such shadows are not depicted in Landsat TM imagery, for which sensor resolution is 30 m (100 ft). We surmise that meter-sized shadows depress the overall reflectivity of the upland forest canopy, which would result in lower values for individual TM bands (1-5 and 7) and lessen the possibility for TM sensor saturation. A similar effect is likely for water-filled hollows on the floor of the bottomland forest. However, band ratios tend to remove such variations in overall brightness (Jensen, 1996), so resulting NDVI values and the band 5:4 ratio should not be adversely affected by small shadows. Although canopy roughness was considered to be more or less constant throughout the study period, differences in canopy roughness should be taken into account in comparisons of the study forests with similar forests elsewhere.
The results of intentional burning of the bottomland prairie illustrate the importance of human management techniques (see fig. 26). Controlled fire is commonly used to maintain native (warm-season) grassland ecosystems in the Great Plains region (Fitch et al., 2001). Such prairies are fire adapted and must burn occasionally to retain their natural character. Spring grassland burning has several important benefits: old, dead grass is removed, which allows rapid growth of new grass; nutrients in the old grass are returned to the soil to fertilize new growth; and woody brush and trees are inhibited from spreading into the prairie. A prairie should be burned once every three to seven years to promote maximum biodiversity and to prevent establishment of trees and brush. The bottomland prairie at Fort Leavenworth is maintained primarily by annual mowing. It had not been burned for twenty years prior to the deliberate fire of April 2000. A large volume of litter and dry vegetation had accumulated, and much brushy vegetation had grown up.
KAP depicts trees injured or killed by the fire on the edge of the bottomland forest. The fire destroyed or damaged basal bark of these trees, which cut off supply of water and nutrients from the roots, and killed some trees outright. Other injured trees were able to grow leaves in their upper branches, based on food stored in the body of the tree. Most of these trees probably did not survive a second year, although some roots may survive to sprout new trees. Field observations in June 2001 confirmed that many trees injured by fire had died (fig. 30). It is unusual for mature trees to be so adversely affected by an understory fire. In fact, it is common practice to burn the understory of upland, hardwood forests in the Chautauqua Hills (southeastern Kansas) and Missouri Ozarks. The authors have observed, however, that trees are much more susceptible to fire damage during drought years, such as 1988-1989 and 2000. Once again, it is notable that the impact of burning is most pronounced at the forest margin.
Fig. 30--Color-infrared kite aerial photograph of the bottomland forest, June 2001. Dead trees (bare trunks) are evident at the forest margin toward the near left side of scene. Compare with fig. 26.

The KAP images demonstrate the effectiveness of fire for controlling tree growth and forest expansion into areas that are managed for grassland habitat. Given the rapid growth rate for cottonwood trees, it would be necessary to burn frequently (at least every third year) to prevent the bottomland forest from spreading farther into the prairie area. Burning in wet years would minimize damage to mature trees, but would still suppress growth of brush and young trees. Early spring burning, prior to tree flowering in early April, could also reduce potential injury to mature trees.
Prev Page--Results || Next Page--Conclusions
Kansas Geological Survey
Web version January 25, 2002
http://www.kgs.ku.edu/Current/2002/aber/aber5.html
email:webadmin@kgs.ku.edu