![]()
![]()
![]()
Prev--North American central-highland fishes || Next--Wisconsinan Mammalian Faunas
Natural History Museum of Los Angeles County, Los Angeles, California
This paper reviews seven previously published and two new Wisconsinan mammalian local faunas from northern Texas to northern Nebraska. Dates for these faunas span the last 18,000 yrs of the Pleistocene. These faunas are analyzed for latitudinally related trends of species composition based on historic distribution of climatically significant mammalian taxa. A very clear latitudinal trend appears in the nine faunas taken as a whole and also within temporally restricted subgroupings. Composition of arvicolid and soricid faunal components is a better latitudinal indicator than is diversity of those components. Unlike Wisconsinan mammalian faunas of the southern and eastern United States, those of the Central and Northern plains consist almost entirely of steppe and boreal taxa. Boreal taxa predominate in the northern faunas.
In 1937, Claude Hibbard published the first of many studies concerning Pleistocene vertebrate faunas of the Southern and Central plains. Many of his students have continued to contribute to this wealth of information. Hibbard's collecting of late Pleistocene vertebrates rarely extended north of Meade County, Kansas, but did include visits to Peoria loess localities in northern Kansas. At the time of the last review of Great Plains Wisconsinan vertebrate faunas (Hibbard, 1970), only two local faunas (the Robert and Jones) were known north of Texas, and none was known north of southern Kansas. The last decade has witnessed a marked increase in collecting of Wisconsinan microvertebrates in Kansas and Nebraska. A considerable amount of information has been available concerning the larger vertebrates of Wisconsinan horizons in Nebraska, but almost nothing has been published until recently concerning the microfauna (Voorhies, 1984). This brief review is intended, in part, as a summary of published and unpublished data now available on the Wisconsinan mammalian microfaunas of the Central and Northern plains. Although very informative nonmammalian microfaunas have been obtained for this region, they are beyond the scope of this work.
Graham (1976) demonstrated that a gradient is found in the diversity of both the Soricidae and Arvicolidae in the late Pleistocene faunas of the United States, with the greatest diversity in the Northeast and the least in the Southwest. Even at the time of that publication, published records of representative Wisconsinan vertebrate microfaunas north of extreme southern Kansas did not exist, and this precluded an examination of any latitudinal gradients throughout the Plains states. Recent advances in research in northern Kansas and Nebraska now permit these comparisons. Therefore, the second purpose of this report is to assess the evidence for latitudinal effects in Plains faunas and to compare these findings with Graham's conclusions (Graham, 1976) regarding the northeast-southwest gradient.
The criteria I will employ here for examining latitudinal effects on faunas are based on historic distributions of mammals, particularly insectivores and rodents. In the Great Plains, Didelphis virginiana, Cryptotis parva, and Procyon lotor are austral indicators. Sorex cinereus is a moderately boreal indicator. Sorex arcticus and S. palustris are much more so. Lepus americanus occurs at higher elevations and latitudes than do jackrabbits (L. californicus and L. townsendii). Heteromyid rodents have an austral connotation. Perognathus fasciatus is somewhat of an exception. Geomys generally occurs at lower elevations and in more southerly areas than does Thomomys. Sigmodon hispidus is an austral cricetid. Among arvicolid rodents, Pedomys ochrogaster is the most austral of the Plains taxa. Occurring at progressively higher latitudes are Synaptomys cooperi, Microtus pennsylvanicus, Clethrionomys gapperi, Phenacomys intermedius, Synaptomys borealis, Microtus xanthognathus, and Dicrostonyx torquatus. Zapus hudsonius is often cited as a boreal taxon (Hoffmann and Jones, 1970; Semken, 1984; Voorhies and Corner, 1985). Yet Zapus princeps, while overlapping the range of Z. hudsonius, has a more northerly distribution. Species today restricted to cordilleran regions are used here as montane indicators. The Spermophilus richardsonii complex requires special comment. Two sibling species exist today: Spermophilus richardsonii occupies the Northern Plains, and three subspecies of Spermophilus elegans occur in montane and intermontane-basin habitats in many western states. A member of this complex inhabited vast areas of the Central and Northern plains during the middle and late Pleistocene. Morphological evidence suggests that the Pleistocene population is not assignable to either species (Neuner, 1976; Neuner and Schultz, 1979) but may be ancestral to both extant species. Although the Pleistocene population cannot be deduced to have strictly the same ecological requirements of either extant taxon, its requirements were not radically dissimilar. Hall (1981) illustrates ranges for all the extant taxa referred to in this paper.
For access to collections in their care, I wish to thank Michael R. Voorhies and R. George Corner. I acknowledge the Army Corps of Engineers, Kansas City district, for support of field work and permission to collect at Harlan County Reservoir. Funds provided by Philip V. Wells and a grant-in-aid of research from Sigma Xi, the Scientific Research Society, made possible some of the radiocarbon dates. Lawrence J. Barnes, John H. Harris, and Larry D. Martin critically read this manuscript. Susan Barnes kindly typed this paper.
In order to facilitate faunal comparisons, table 1 shows a compilation of the occurrences of most of the austral or boreal or montane taxa in the local faunas discussed in this section. Hibbard (1940) first reported mammalian taxa from the Jones local fauna (fig. 1, no. 3) of Meade County, Kansas, and supplemented this account with later reports (Hibbard, 1942, 1949). Davis (1975) recorded additional taxa. The only taxa from this local fauna which could be said to have boreal or montane affinities are Sorex arcticus, S. cinereus, Thomomys sp., Microtus pennsylvanicus, and a member of the Spermophilus richardsonii complex. Davis (1975) also deduced the presence of Microtus "alpha," having a dentition similar to Microtus longicaudus or M. montanus. Geomys is present in addition to Thomomys. Pedomys ochrogaster and one heteromyid are present. Mollusks for this horizon yielded dates of 26,700±1,500 yrs B.P. (I-3461) and 29,000±1,300 yrs B.P. (I-3462), placing it in the Farmdalian substage of the Wisconsinan stage.
Figure 1--Map showing locations of Wisconsinan faunas discussed in this work. The sites are 1) Howard Ranch local fauna, 2) Robert local fauna, 3) Jones and Classen local faunas, 4) Duck Creek local fauna, 5) Trapshoot local fauna, 6) Coon Creek local fauna, 7) Red Willow quarries, 8) North Cove local fauna, 9) Litchfield local fauna, and 10) Smith Falls local fauna.
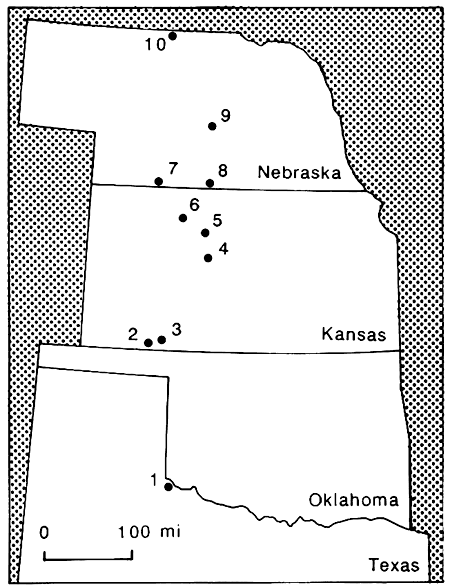
Table 1--Distribution of some austral and boreal taxa in nine local faunas discussed in text. Site numbers refer to same sites as in fig. 1; they also are listed in approximate order of increasing latitude.
| Taxa | Sites | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| South | North | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 8 | 9 | 10 | ||
| ↑ boreal/ montane |
Dicrostonyx torquatus | x | ||||||||
| Ochotona princeps | x | |||||||||
| Gulo gulo | x | |||||||||
| Tamiasciurus hudsonicus | x | x | ||||||||
| Marmota flaviventris | x | ? | ||||||||
| Eutamias sp. | x | ? | ||||||||
| Lepus americanus | x | x | x | |||||||
| Microtus xanthognathus | x | x | x | x | ||||||
| Phenacomys intermedius | x | x | x | x | x | |||||
| Zapus princeps | x | x | x | |||||||
| Synaptomys borealis | x | x | x | |||||||
| Clethrionomys gapperi | x | x | x | x | x | |||||
| Sorex arcticus | x | x | x | x | ||||||
| Microtus montanus | ? | ? | x | x | x | x | x | |||
| Sorex palustris | x | x | ||||||||
| Thomomys sp. | x | x | x | x | x | x | x | x | x | |
| Spermophilus richardsonii complex | ? | x | x | x | x | x | x | x | x | |
| Sorex cinereus | x | x | x | x | x | x | ||||
| Microtus pennsylvanicus | x | x | x | x | x | x | x | x | ||
| Zapus hudsonius | x | x | ||||||||
| Synaptomys cooperi | x | x | x | ? | ||||||
| austral ↓ |
Geomys sp. | x | x | x | x | x | x | |||
| Pedomys ochrogaster | x | x | x | ? | x | x | ||||
| Cryptotis parva | x | x | x | |||||||
| Perognathus sp. | x | x | x | |||||||
| Perognathus hispidus | x | x | ||||||||
| Dipodomys ordii | x | |||||||||
| Sigmodon hispidus | x | |||||||||
| Procyon lotor | x | |||||||||
| Didelphis virginia | x | |||||||||
Shells from the only Woodfordian site in southern Kansas, the Classen local fauna (fig. 1, no. 3) of Meade County, produced a date of 16,100±250 yrs B.P. (I-4930). Davis (in press) described the vertebrate remains of that fauna. Also occurring within this county is a fauna of Twocreekan age, the Robert local fauna (fig. 1, no. 2), for which Schultz (1967) reported a date of 11,100±390 yrs B.P. (SM-762). Mammalian taxa with boreal affinities include Sorex cinereus, S. palustris, Thomomys cf. T. talpoides, Microtus pennsylvanicus, Synaptomys cooperi, Zapus cf. Z. hudsonius, and a member of the Spermophilus richardsonii complex. Geomys also is present, although Schultz (1967) did not signify whether it is more abundant than Thomomys. Likewise, he did not indicate the abundance of Pedomys ochrogaster relative to Microtus pennsylvanicus. No heteromyids are known from this fauna.
Moving 197 km (118 mi) north, we next encounter the Duck Creek local fauna from Ellis County, Kansas (fig. 1, no. 4). It has been the basis of several articles (Zakrzewski and Maxfield, 1973; Kolb et al., 1975; McMullen, 1975, 1978; Holman, 1984). All these studies considered the local fauna to be of "Illinoian" age; geomorphologic and biostratigraphic evidence, however, corroborate a Woodfordian radiocarbon date of 17,700±350 yrs B.P. (GX 9354), obtained from shells of Sphaerium cf. S. striatinum. Two of its arvicolid taxa never occur in Wisconsinan faunas of the Southern Plains. These are Clethrionomys gapperi (Zakrzewski and Maxfield, 1973) and Synaptomys borealis (McMullen, 1978). Sorex arcticus, S. palustris, Microtus pennsylvanicus, a member of the Spermophilus richardsonii complex, and either Mustela ermina or M. nivalis also are present. No heteromyids are present. McMullen (1978) also reported two M2's, which he assigned to Pedomys ochrogaster. The presence of a four-element M2 in a microfauna is insufficient basis for an identification of P. ochrogaster (Stewart, 1978). Microtus montanus has such an M2 and occurs in all described Wisconsinan microfaunas north of Ellis County. In light of this fact and also the observation that arvicolid M1's are more frequently preserved than M2's (no P. ochrogaster M1's were found), these teeth probably belong to M. montanus. The geomyids of this site include both Geomys and Thomomys, although the latter is markedly more numerous. This local fauna comes from fluvial sediments.
A lesser known Wisconsinan fauna, the Trapshoot local fauna, comes from the next county to the north, Rooks County (fig. 1, no. 5). In contrast to most Plains microfaunas, it lies essentially within a paleosol. The local fauna may be a correlative of the "Citellus zone" fauna of the basal paleosol of the Wisconsinan loess of Nebraska. Upland organisms strongly dominate the faunal list as a result of this geological context (Stewart and Rogers, 1984). In accordance with this fact, it lacks any record of aquatic mollusks, fishes, Rana, Synaptomys, or Ondatra. Another consequence is that the soricid record is very poor, represented by only a single femur. Species present here, but not known from faunas to the south, include Zapus princeps, Perognathus fasciatus, Phenacomys intermedius, and Sorex nanus (Stewart, 1978; Hoffmann and Owen, 1980). The second most common species at this site is a member of the Spermophilus richardsonii complex. The montane vole, Microtus montanus, also is well represented here. This is probably the identity of the Microtus "alpha" of Davis (1975) from the Jones local fauna (Stewart, 1978). Microtus pennsylvanicus, the most common species in the Trapshoot local fauna, outnumbers M. montanus by a factor of two to one. A single individual documents the presence of Pedomys ochrogaster in the Trapshoot local fauna. Another fauna of this age and upland character comes from central Rooks County, but it lacks any arvicolids. In both these faunas, Thomomys is the only known geomyid, and the only Lepus species are L. californicus and L. townsendii. No absolute dates are available for these faunas, but faunistic evidence indicates a Wisconsinan age. A more detailed account of this local fauna is forthcoming (Stewart, in press).
A third fauna, which continues this increasingly boreo-montane trend, is the Coon Creek local fauna, 50 km (30 mi) to the west-northwest in Graham County, Kansas (fig. 1, no. 6). As in all local faunas discussed thus far, Microtus pennsylvanicus and a member of the Spermophilus richardsonii complex occur here. Also present is Mustela nivalis. Like the Duck Creek and Trapshoot local faunas, the Coon Creek produces Clethrionomys gapperi. It also shares Phenacomys and Zapus princeps with the Trapshoot local fauna. Microtus montanus furnishes a montane aspect and outnumbers M. pennsylvanicus by a factor of two to one. The yellow-cheeked vole, Microtus xanthognathus, here marks its southernmost occurrence in the Plains. Neither Pedomys ochrogaster nor heteromyids are present. Again, Thomomys is the only geomyid. A date obtained from Bison bones was 17,930±550 yrs B.P. (GX 9355; Wells, 1983). The local fauna occurs in a redeposited loess. Numerous remains of Rana, Ambystoma, cyprinids, and catostomids indicate that this deposit represents alluvium rather than colluvium.
In extreme southern Nebraska, 88 km (53 mi) north of the Trapshoot local fauna and 87 km (52 mi) northeast of the Coon Creek local fauna, lies the recently discovered North Cove local fauna in Harlan County (fig. 1, no. 8). Taxa which continue the trend of boreal affinities include Sorex cinereus, S. arcticus, a member of the Spermophilus richardsonii complex, Phenacomys, Clethrionomys, Synaptomys borealis, Microtus xanthognathus, Zapus princeps, and Mustela nivalis. The trend is further accentuated by Lepus americanus and the southernmost Plains records of Tamiasciurus hudsonius, Eutamias minimus, and Marmota cf. M. flaviventris. An extinct marten, Martes nobilis, and Microtus montanus emphasize the montane influence on this local fauna. As in the Coon Creek and Trapshoot local faunas, Thomomys is the only geomyid. As in the former, Pedomys ochrogaster and heteromyids are absent. This is the first fauna in this survey which has arboreal taxa. Wood of Picea glauca provided a date of 14,700±100 yrs B.P. (Beta 12286) for this site. All evidence indicates a fluvial origin for this deposit. The megafaunal component of this local fauna is rather limited but probably would be similar to the fossils recovered from the Red Willow quarries just 80-114 km (48-68 mi) upstream on the Republican River (fig. 1, no. 7). Taxa collected there include Panthera atrox, Ovis canadensis, Symbos cavifrons, Ovibos moschatus, and Rangifer tarandus (Corner, 1977). No date is available for the Red Willow quarries, but the bulk of the assemblage accords with a Wisconsinan age.
The next Wisconsinan microfauna that we encounter along our northward transect is the Litchfield local fauna of Sherman County, Nebraska (fig. 1, no. 9). This site lies 120 km (72 mi) north of the North Cove fauna. Certainly this local fauna, reported by Voorhies and Corner (1985), exhibits some of the boreal characteristics observed in the previous faunas. Sorex cinereus, a member of the Spermophilus richardsonii complex, Microtus montanus, M. xanthognathus, Phenacomys, Clethrionomys, Thomomys, and Lepus americanus occur there. If this fauna were to follow all the trends we have observed, we would expect to see no Pedomys, no Geomys, and Synaptomys borealis rather than S. cooperi. However, this is not the case. Geomys is five times more abundant than Thomomys, and Pedomys is more numerous than Microtus pennsylvanicus and M. montanus combined. The only bog lemming is Synaptomys cooperi, and Zapus hudsonius is the only zapodid present (personal observation). I believe that these findings may indicate that the Litchfield local fauna is either significantly older (early Wisconsinan) or later (latest Wisconsinan) than the other faunas being considered in this region, as will be detailed below. This local fauna comes from alluvium capped by loess and apparently incised into older loess.
The Smith Falls local fauna (fig. 1, no. 10), which is the northernmost local fauna in this survey, actually comprises samples from 11 sites along the Niobrara River in Cherry, Brown, and Keya Paha counties, Nebraska (Voorhies and Corner, 1985). These sites are approximately 210 km (126 mi) north-northwest of the Litchfield local fauna. Boreal or montane mammalian taxa seen in other local faunas surveyed include Lepus americanus, Tamiasciurus, Phenacomys, Microtus montanus, and M. xanthognathus. Four taxa that are, within the context of the Plains, unique to the Smith Falls local fauna include Ochotona, Dicrostonyx torquatus, Martes americana, and Gulo gulo. As in most of the more northerly faunas, Pedomys ochrogaster and heteromyids are absent. Conspicuous by its absence is Microtus pennsylvanicus, which is common in every other Plains Wisconsinan microfauna from the Texas panhandle northward. Although Thomomys is not the only geomyid, it outnumbers Geomys by a factor of four to one. This is the only Plains local fauna which has what might be termed tundra components. No radiocarbon date for this local fauna is available, but Voorhies and Corner (1985) suggested that it is somewhat older than the Litchfield local fauna. As these authors demonstrated, the fluvial sediments that produce this fauna constitute a high terrace, presumably of the Niobrara River.
The climate during the Wisconsinan glacial stage was not static. Thus, testing for environmental gradients without regard for heterochroneity is inappropriate. The majority of the dated sites in this study are of Woodfordian age, falling within the dates of 15,000-20,000 yrs B.P. These include the Duck Creek, Coon Creek, and North Cove local faunas. The Smith Falls local fauna probably belongs to this group. The Trapshoot local fauna may be of this age or may be older. The published Meade County Wisconsinan local faunas are of Farmdalian and Twocreekan ages. Voorhies and Corner (1985) suggest that the Litchfield local fauna is younger than the Smith Falls local fauna. Some aspects of the Litchfield local fauna certainly indicate less boreal conditions than in the nearby Woodfordian faunas. Among these aspects are the high proportion of Geomys to Thomomys, the presence of Zapus hudsonius rather than Z. princeps, and an extremely high representation of Pedomys ochrogaster. These characteristics could accord with either a pre- or post-Woodfordian date. However, the presence of several mammalian taxa including Microtus xanthognathus and mollusks such as Discus shimekii and Columella alticola indicates that its age does not antedate the Woodfordian. These species could possibly persist for some time after the glacial maximum. For example, Microtus xanthognathus occurs as late as 11,300 yrs B.P. at the New Paris No. 4 site in Pennsylvania. However, no date for this taxon is older than 20,530 yrs B.P. in the coterminous United States.
Within the Woodfordian faunas, we see a definite correlation between increasing latitude and increasingly boreal affinities of the local faunas. With the exception of the North Cove local fauna, each has one or two arvicolid rodent taxa not seen in the local faunas to the south. All have some mammalian taxa not found in faunas to the south. Likewise, austral elements such as heteromyids and Pedomys ochrogaster drop out with increasing latitude. All these faunas contrast starkly with Woodfordian faunas no farther south than the Texas-Oklahoma border (Dalquest, 1965). If we assume a Twocreekan age for the Litchfield local fauna (possibly an underestimate), we may compare it to the Robert local fauna in southern Kansas. Nearly all the taxa of the Robert local fauna occur in the Litchfield local fauna. However, many additional boreal elements appear in the latter.
Graham (1976) formulated five conclusions resulting from his study of Wisconsinan microfaunas from Pennsylvania to Texas. I wish to examine each of these conclusions in relation to the faunas of the Central and Northern plains.
Graham first concluded that most late Wisconsinan mammalian local faunas were composed of more boreal and deciduous species than are the modern faunas in these areas. This is certainly not true for the area of the present study. Hoffmann and Jones (1970) noted that the Jones local fauna, in contrast with the Texas local fauna, was composed almost entirely of steppe taxa, with a few boreal taxa. The fauna of southwestern Kansas today has many more deciduous elements and almost no boreal elements. The Woodfordian local faunas of northern Kansas and Nebraska accord with the Jones local fauna in this respect. However, they have a higher percentage of boreal taxa. The relative proportions of steppe and boreal components seem to be related to latitude. In some of the northern Kansas local faunas, the boreal component becomes predominant. Not far south of the Kansas-Nebraska border, the boreal component is larger than the steppe and nondiagnostic components combined.
Graham's second conclusion was that arvicolid and soricid species diversity was greater during the late Wisconsinan, but formed a gradient as in the modern fauna. While sampling of the soricids of the Plains Wisconsinan faunas may be incomplete, no well-defined gradient in soricid diversity has been found in either the Woodfordian or Twocreekan faunas. In fact, there are as many soricid taxa (4) in the Woodfordian Howard Ranch local fauna of Hardeman County, Texas (Dalquest, 1965), as in any of the Central and Northern plains faunas. Wisconsinan diversity is as great or slightly greater than that of the modern fauna at any given site. The Twocreekan fauna seems to have a slightly greater soricid diversity than does the Woodfordian fauna. The arvicolids show a different pattern. A slight latitudinal increase in diversity occurs in the Woodfordian fauna, but we see an apparently stronger gradient in the Twocreekan fauna. In all cases, the Pleistocene diversity of arvicolids (4-7) is significantly greater than the current diversity at those sites (1-3). Clearly, the composition of soricid and arvicolid faunal components is of greater significance than the diversity of these components.
Further, Graham (1976) concluded that the soricid and arvicolid gradients in the second conclusion indicate moderated longitudinal and latitudinal gradients in temperature and moisture during the late Pleistocene. As observed above, I have not detected as much evidence of Pleistocene diversity gradients in the Plains. However, even if this were not so, how greater Pleistocene faunistic gradients translate into moderated climatic gradients is not clear. I would not, however, contest a moderated Pleistocene climate.
The fourth conclusion that Graham listed was that, although species restricted today to boreal regions were wide-ranging during the late Wisconsinan, resident species were not displaced further to the south. Certainly the Plains sites show that such taxa as Sorex arcticus, the Spermophilus richardsonii complex, Thomomys talpoides, Microtus montanus, and Microtus xanthognathus were more widespread in the Wisconsinan than they are today. However, examination of the modern faunal lists of those areas readily show that many current residents did not range as far north as they do today. Among soricids and arvicolids, Cryptotis parva, Pedomys ochrogaster, and in one case, Microtus pennsylvanicus are examples. Didelphis, Procyon, heteromyids, Zapus hudsonius, Oryzomys palustris, and Sigmondon hispidus are other mammalian examples of the case in point. Dozens of additional examples could be cited among the herpetofaunas.
Graham (1976) finally concluded that the modern patterns in distribution and diversity of small mammals are the result of a more continental post-glacial climate. I support this conclusion for the Wisconsinan microfaunas of the Plains.
Hibbard (1970) mentioned that today's extreme winter temperatures developed during Wisconsinan times. To say that some of the organisms then living in the Plains, today live in regions with cold winters is not a demonstration that these taxa indicate extreme winter temperatures. They may merely require moderate summer temperatures. If Hibbard were correct in his contention, then a most remarkable climatic gradient would have had to exist to accommodate Geochelone at that time only approximately 700 km (420 mi) to the south in Oklahoma, Texas, and New Mexico.
Recent discoveries in the Central and Northern plains enable us to discern definite latitudinal effects in the Wisconsinan mammalian faunas of the Plains. Arvicolid diversity generally reflects this trend, but soricid diversity does not. Composition of arvicolid and soricid faunal components are much better indicators of latitudinal effects than is diversity of those components. In contrast to Wisconsinan faunas of the south and east, those of the Central and Northern plains consist almost entirely of steppe and boreal taxa. Boreal taxa predominate in those to the north.
Corner, R. G., 1977, A late Pleistocene and Holocene vertebrate fauna from Red Willow County, Nebraska: Transactions of the Nebraska Academy of Sciences and Affiliated Societies, v. 4, p. 77-93.
Dalquest, W. W., 1965, New Pleistocene formation and local fauna from Hardeman County, Texas: Journal of Paleontology, v. 39, no. 1, p. 63-79.
Davis, L. C., 1975, Late Pleistocene geology and paleoecology of the Spring Valley basin, Meade County, Kansas: Ph.D. thesis, University of Iowa, 170 p.
Davis, L. C., in press, The mammalian record for Late Pleistocene and Holocene environmental changes in the Central Plains of the United States; in, Late Quaternary Mammalian Biogeography and Environments, R. W. Graham, H. A. Semken, Jr., and M. A. Graham, eds.: Illinois State Museum Scientific Papers.
Graham, R. W., 1976, Late Wisconsinan mammalian faunas and environmental gradients of the eastern United States: Paleobiology, v. 2, p. 343-547.
Hall, E. R., 1981, Mammals of North America: New York, John Wiley & Sons, 2 vols., 1,181 p.
Hibbard, C. W., 1937, A new Pitymys from the Pleistocene of Kansas: Journal of Mammalogy, v. 18, p. 235.
Hibbard, C. W., 1940, A new Pleistocene fauna from Meade County, Kansas: Transactions of the Kansas Academy of Science, v. 18, no. 2, p. 235.
Hibbard, C. W., 1942, Pleistocene mammals from Kansas: Kansas Geological Survey, Bulletin 41, part 6, p. 261-269. [available online]
Hibbard, C. W., 1949, Pleistocene stratigraphy and paleontology of Meade County, Kansas: Contributions from the Museum of Paleontology, University of Michigan, v. 7, no. 4, p. 63-90.
Hibbard, C. W., 1970, Pleistocene mammalian local faunas from the Great Plains and Central Lowland provinces of the United States; in, Pleistocene and Recent Environments of the Central Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: University Press of Kansas, Lawrence, University of Kansas, Department of Geology, Special Publication 3, p. 395-433
Hoffmann, R. S., and Jones, J. K., Jr., 1970, Influence of late-glacial and post-glacial events on the distribution of Recent mammals on the northern Great Plains; in, Pleistocene and Recent Environments of the Central Plains, W. Dort, Jr., and J. K. Jones, Jr., eds.: University Press of Kansas, Lawrence, University of Kansas, Department of Geology Special Publication 3, p. 355-394.
Hoffmann, R. S., and Owen, J. G., 1981, Sorex telenus and Sorex nanus: Mammalian Species, no. 131.
Holman, J. A., 1984, Herpetofaunas of the Duck Creek and Williams local faunas (Pleistocene-Illinoian) of Kansas; in, Contributions in Quaternary Vertebrate Paleontology-A Volume in Memoriam to John E. Guilday, H. H. Genoways and M. R. Dawson, eds.: Special Publications of Carnegie Museum of Natural History, no. 8, p. 20-38.
Kolb, K. K., Nelson, M. E., and Zakrzewski, R. J., 1975, The Duck Creek molluscan fauna (Illinoian) from Ellis County, Kansas: Transactions of the Kansas Academy of Science. v. 78. no. 1-2, p. 63-74.
McMullen, T. L., 1975, Shrews from the late Pleistocene of central Kansas, with the description of a new species of Sorex: Journal of Mamma1ogy, v. 56, no. 2, p. 316-320.
McMullen, T. L., 1978, Mammals of the Duck Creek local fauna, late Pleistocene of Kansas: Journal of Mammalogy, v. 59, no. 2, p. 374-386.
Neuner, A. M., 1976, Evolution and distribution of the Spermophilus richardsonii complex of ground squirrels in the middle and late Pleistocene--a multivariate analysis: M.A. thesis, University of Kansas, 42 p.
Neuner, A.M., and Schultz, C. B., 1979, Spermophilus from the "Citellus zone" of the late Quaternary of the Central Plains: Transactions of the Nebraska Academy of Sciences and Affiliated Societies, v. 7, p. 101-104,
Schultz, G. E., 1967, Four superimposed late Pleistocene vertebrate faunas from southwest Kansas; in, Pleistocene Extinctions--The Search for a Cause, P. S. Martin and H. E. Wright, Jr., eds.: Yale University Press, New Haven, Connecticut, p. 321--336.
Semken, H. A., Jr., 1984, Paleoecology of a late Wisconsinan/Holocene micromammal sequence in Peccary Cave, northwestern Arkansas; in, Contributions in Quaternary Vertebrate Paleontology--A Volume in Memoriam to John E. Guilday, H. H. Genoways and M. R. Dawson, eds.: Special Publication of Carnegie Museum of Natural History, no. 8, p. 405-431.
Stewart, J. D., 1978, Mammals of the Trapshoot local fauna, late Pleistocene of Rooks County, Kansas (abs.): Proceedings of the Nebraska Academy of Sciences and Affiliated Societies, p. 45-46.
Stewart, J.D., and Rogers, R. A., 1984, Analysis of pollen from the Trapshoot local fauna quarry (Rancholabrean) of Kansas: American Midland Naturalist, v. 112, no. I, p. 198-200.
Voorhies, M. R., 1984, "Citellus kimballensis" Kent and "Propliophenacomys uptegrovensis" Martin, supposed Miocene rodents, are Recent intrusives: Journal of Paleontology, v. 58, no. 1, p. 254-258.
Voorhies, M. R., and Corner, R. G., 1985, Small mammals with boreal affinities in late Pleistocene (Rancholabrean) deposits of eastern and central Nebraska: Institute for Tertiary-Quaternary Research, TER-QUA Symposium Series, v. 1, p. 125-142.
Wells, P. V., 1983, Late Quaternary vegetation of the Great Plains: Transactions of the Nebraska Academy of Sciences and Affiliated Societies, v. 11. p. 83-89.
Zakrzewski, R. J., and Maxfield. J. L., 1973, Occurrence of Clethrionomys in the late Pleistocene of Kansas: Journal of Mammalogy, v. 52, no. 3, p. 620-621.
Prev--North American central-highland fishes || Next--Wisconsinan Mammalian Faunas
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version updated March 29, 2010. Original publication date 1987.
URL=http://www.kgs.ku.edu/Publications/Bulletins/GB5/Stewart/index.html