![]()
![]()
![]()
Prev--Wisconsinan sediments on the High Plains of Kansas || Next--Wisconsinan Mammalian Faunas
University of California, Los Angeles, California
Rivers of the central highland regions of North America contain one of the most diverse assemblages of fishes of the continent. Each region is distinctive, with a high degree of endemism and only a few species shared between any two of the areas. For many years biologists have noted this pattern of diversity and commented on possible explanations for its origin. They noted similarities between regions either with allopatric populations of conspecifics or hypothesized close relatives in two or more areas. Previous explanations have adopted the philosophy of evolutionary biogeography. Centers of origin were recognized and species were hypothesized to have dispersed outward to unoccupied regions. This philosophy is compared with that of vicariance biogeography for central-highland fishes and found to be inferior with respect to its ability to provide a general explanation of common patterns. The known geological history of the region and observed pattern of relationships and distributions of central-highland fishes are consistent with the tenets of vicariance biogeography and necessitate fewer ad hoc explanations with this model. Although the three major highland regions are separated by lowland environments, geological evidence suggests that the intervening lowland areas are recent formations since the Pleistocene and that the isolated areas were once a single expansive highland region. Highland topography, and presumably rivers, extended north of the existing highland areas into the central lowlands. The impact of continental glaciers on this region and the fauna was tremendous. Ice sheets destroyed highland habitats in all areas of coverage and essentially separated this expansive region into large western and eastern components. The western component, interior highlands, underwent vicariance at sometime prior to the Sangamonian interglacial, resulting in the existing separate Ozark and Ouachita highlands. Species relationships of highland-fish groups suggest several patterns with one pattern being common. That is, species in the Ouachita and Ozark highlands are more closely related to each other than either is to species in the eastern highlands. Species of the upper Mobile Bay drainages form the sister group to interior and eastern highland species. The Mobile Bay separation is the oldest event, perhaps dating to the Miocene. The separation of the interior highlands can minimally be dated as pre-Sangamonian. Speciation events between the eastern and interior highlands predate the Pleistocene. Only with the methods and philosophy of phylogenetic systematics and vicariance biogeography can the temporal sequence of these historical events be uncovered. Further, hypotheses presented herein form general explanations that may be tested with additional geological and phylogenetic data as they become available. Such is not the case with the previous explanations of dispersal from designated centers of origin. Dispersal is not excluded as a viable explanation in vicariance. Dispersal is a very real phenomenon and must have occurred at some stage. However, it is a difficult proposition to test since everything is possible given this philosophy. The dispersal explanation shadows other, perhaps more parsimonious, explanations in biogeographic studies and should be a conclusion in these studies, not an assumption.
Biogeography, as generally defined, is that area of biology dealing with the geographical distribution of animals and plants. Although very elementary on the surface, this area of biology is diverse in its methods and philosophy and becoming ever more important in areas of evolutionary biology. Very generally, it may be subdivided into two major areas of emphasis: descriptive and interpretive biogeography. Historical biogeography forms the interpretive portion of the field and seeks to explain the present distributions of plants and animals elucidated by the field of descriptive biogeography. Historical explanations for present species distributions are sought through the use of species relationships and ecological and geological histories of regions inhabited by them. The two areas of biogeography have been compared by Platnick and Nelson (1978), Nelson and Platnick (1980), and Wiley (1981) to the collection and analysis of data in any scientific study.
Little variation exists among biogeographers concerned with descriptive biogeography, but the same cannot be said for interpretive biogeography. The former is primarily concerned with present-day species distributions and how they are controlled, maintained, or lost. On the other hand, the latter area is one of varied thought, but common to all is their interest in the spatial and temporal distributions of species or higher taxa and what historical events provide explanations for present-day distributions. The division between ecological and historical biogeography is tenuous. It is difficult to separate the two fields because they are, in fact, interdependent and are best considered two extremes of a continuum. Although inseparable, emphasis in this paper is on the historical extreme of this continuum. This paper complements ideas presented by Cross (1970) by incorporating ideas of vicariance biogeography of other North American fishes and represents a broader view of how these distributions and distributions of other central North American fishes have changed since the Pleistocene. More specifically, this paper concerns how the Pleistocene glaciation affected those fishes thought to have been restricted to an uplifted region in east-central North America with typically clear, high-gradient streams, commonly referred to as the central highlands (Mayden, 1985; Wiley and Mayden, 1985).
As previously observed by Nelson and Platnick (1980), the field of historical biogeography is unlike many areas of biology in that many researchers interested in this field are not biogeographers per se, but are systematists primarily interested in developing hypotheses of species relationships. However, they also are interested in discovering factors influential in producing observed patterns of species relationships. As biogeographers, these researchers are concerned with species distributions, geological histories, and biotic regions and how these regions are interrelated. These latter concerns are not unique to systematics; they are of general interest to other areas of organismic biology. Only with these historical kinds of data can certain evolutionary and ecological questions be answered. For example, factors influential in producing present-day patterns of species distributions, regions, etc., are products of historical phenomena and are best estimated with historical data in the form of species phylogenies and geological histories. Thus, the field is occupied primarily by systematists, the only biologists with the data base for direct estimates of historical events.
Reconstructed species phylogenies represent an estimate of the past history of a group and are presented in the form of trees, representing genealogies. From these reconstructions the biologist can determine relative ages of species and speciation events and can superimpose species trees onto areas occupied by the species to obtain direct estimates of histories of regions (Nelson and Platnick, 1980). These predictions can then be tested using geological data (also in the form of trees), if available, and then congruence between the two independently derived data sets can be examined. Such a philosophy, that the earth and its inhabitants evolve together and can be studied in concert, was termed hologenesis by Rosa (1918).
Basically, two approaches to historical biogeography exist: evolutionary biogeography and vicariance biogeography. A third approach exists, phylogenetic biogeography, but its methodology is very similar to the vicariance approach. Vicariance biogeography is the most robust area of biogeography and is employed later in this paper. However, before further discussing the vicariance method, these other areas must be addressed.
The field of phylogenetic biogeography was principally developed by Hennig (1960) and Brundin (1966, 1972) and does not differ extensively from vicariance biogeography. When applied, however, it is most frequently with a single group of organisms and general explanations usually are not sought. The birth of this field of biogeography was in the development of phylogenetic systematics. The biogeographic history of a group is inferred from its phylogenetic history. The philosophy of phylogenetic biogeography is employed in vicariance biogeography, except the latter school of thought deals with multiple groups of organisms in search for common patterns and general explanations.
The philosophy of evolutionary biogeography is embodied by a dispersalist viewpoint. Here researchers place emphasis on centers of origin of groups and from these centers new species evolve and displace more primitive species to the periphery. This philosophy primarily is derived from ideas of progressive evolution. New species are better adapted and distributional patterns of organisms can be explained in terms of dispersal. Although dispersal is a very real phenomenon in nature, it is difficult to test. Further, in assuming that all patterns can be explained in terms of dispersal, a researcher is likely to lose valuable information. Even if only a small proportion of speciation events and geographic distributions are the result of some mode (i.e., vicariance) other than dispersal, these processes will never become known when one only employs a dispersalist explanation. With an increasing percentage of vicariance or other modes, we would only see an increase in lost information. If we assume that disjunctions in distributions are the result of dispersal, then we will never uncover cases to the contrary. Thus, the philosophy of evolutionary biogeography is not employed in this paper.
Vicariance biogeography as a discipline represents the union of the principles of phylogenetic systematics (Hennig, 1966) and track analysis (Croizat, 1952, 1958, 1964). It is a method whose basic tenets are to furnish a general method of biogeographic analysis applicable at any level and to seek general explanations for patterns (see Nelson and Rosen, 1981). Thus, it minimizes the number of unnecessary explanations for a common pattern. The method attempts to explain common distributional patterns with a general explanation such as the fracturing of a once widespread biota into smaller, distinctive biotas. Also, it allows one to separate these common patterns from those which require additional and unique explanations such as long-distance dispersal. This approach to biogeography is less dependent on ad hoc dispersal explanations.
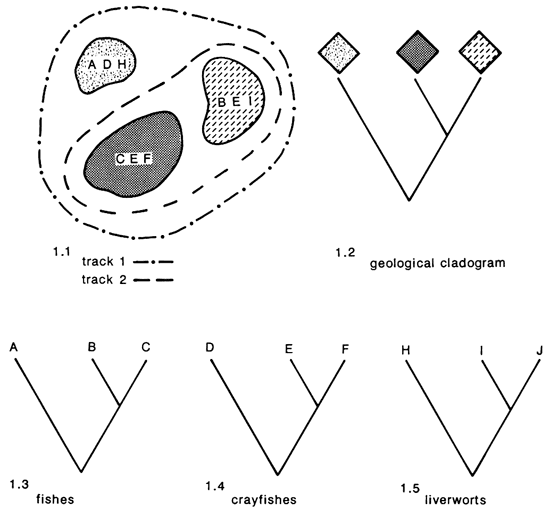
The vicariance biogeographer seeks distributional data from several monophyletic groups (all descendents of a common ancestor) inhabiting a region, and the replicated patterns of distributions of these groups are termed tracks (fig. 1.1). From these tracks, areas of endemism are identified and relationships of these areas, in terms of species inhabiting them, are studied. The relationships of the organisms found in areas of endemism are compared to the geological relationships of the regions for concordance. The degree to which we have congruence between the geological and species cladograms is related to the frequency of common factors affecting the evolution of the groups. If we see congruence between the geological cladogram (fig. 1.2) and one or more of the biological cladograms (fig. 1.3-1.5), then we may hypothesize that factors responsible for producing the former pattern were responsible for the latter phylogenetic relationships. These methods are very powerful and offer considerable predictive potential for both geologists and biogeographers. Given that one of these two historical data sets (geological or biological cladogram) is missing, one may make predictions concerning the missing component based on available data. As an example, in fig. 1 if only species relationships are available, then one may predict that areas contained in track 2 share a more recent history than either has with the third area contained in track 1. These hypotheses may then be tested later when additional data on these areas are available. The vicariance approach is superior to other existing areas of biogeographic analysis and is employed here for studying the biogeographic history of central-highland fishes.
Figure 1--Distributions of three areas of endemism, two biogeographic tracks, a geological cladogram of areas of endemism, and phylogenetic relationships of three monophyletic endemic groups of organisms.
Information presented below represents an abbreviated review and synthesis of contributions by several researchers interested in this topic. It summarizes, with a few examples, our present knowledge of the history of the fishes and the region presently inhabited by them. For more detailed information concerning the descriptive aspects of this fauna, the reader is referred to references in Mayden (1985, in press) and Wiley and Mayden (1985). Information concerning the geological history of the region is scattered but is more fully outlined in Thornbury (1965), Bretz (1965), and other references in Mayden (1985, in press). Interpretive studies of highland biogeography include Dowling (1956), Metcalf (1966), Cross (1970), Pflieger (1971), Mayden (1985, in press), Wiley and Mayden (1985), and Cross et al. (1985). Below, I discuss the biogeography of these fishes and how the Pleistocene glaciation has affected their present distributions. The methodology of vicariance biogeography is employed in determining the temporal history and relationships of the regions. Further, the distributions of some central Kansas fishes, which are typically highland species, are evaluated in terms of their probable origin.
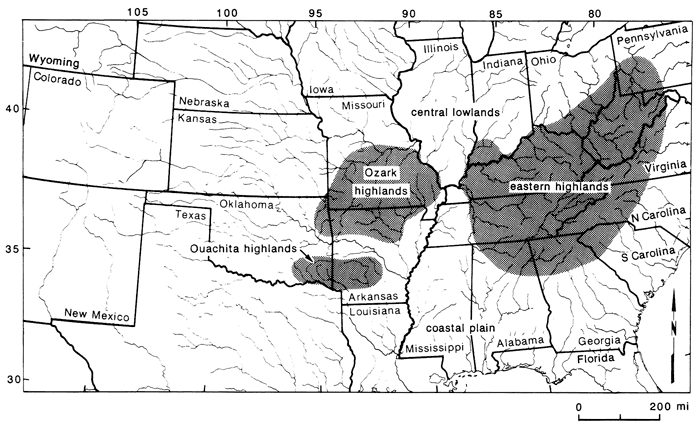
The central highland region includes three areas east and west of the Mississippi Valley (Mayden, 1985, in press). Included are the Ozark highlands, Ouachita highlands, and eastern highlands (fig. 2). These geological formations are described in more detail in Mayden (1985, in press). Presently, the areas are typically drained by high-gradient and clear streams, all part of the Mississippi River system. Separating the areas are low-gradient habitats with typically silt-loaded streams, generally not occupied by fishes typical of the highland regions. To the south, the highlands are bounded and partially dissected by the Coastal Plain province. The Ozark and Ouachita highlands (interior highlands) also are divided by this province and the floodplain of the Arkansas River. Between the interior highlands and the eastern highlands is the floodplain of the Mississippi. North of the highlands lies the characteristically low-gradient Central Lowlands province, an area displaying the impact of Pleistocene glaciation. West of the interior highlands are the unglaciated central lowlands and Great Plains. The Ouachita highland region is the smallest of the three regions and is drained by the Ouachita, Kiamichi, and Little rivers. The Ozark highlands are intermediate in size and are drained by portions of the Missouri, White, and Arkansas river systems. The eastern highlands is by far the largest area and has the largest number of species. This region is drained by portions of the Ohio, Cumberland, and Tennessee rivers, and the upper Mobile Bay and Appalachicola rivers of the Gulf Coast.
Figure 2--Major central highland regions in North America (after Mayden, in press).
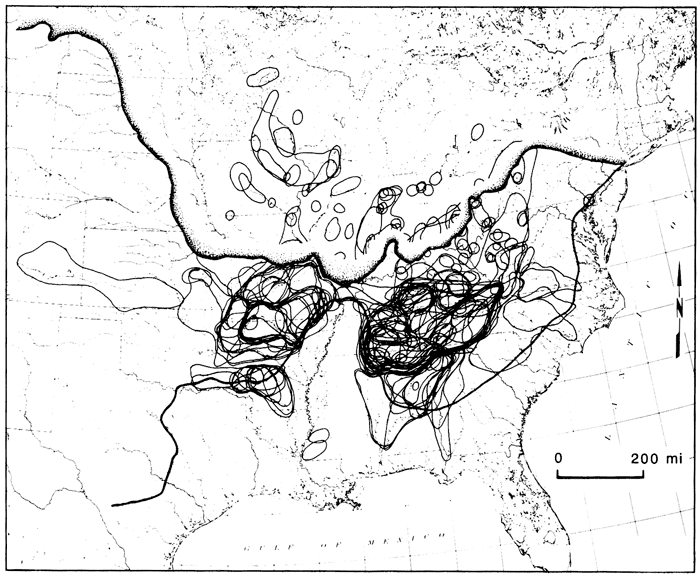
The three major regions of the central highlands contain one of the most diverse faunas in North America. As a whole, it contains greater than half the total diversity of the Mississippi River system. Each highland area is known for its diverse and endemic fauna (fig. 3; Mayden, in press). The highland areas also are known to have several exclusively shared species or species groups between two or three of the regions. Fish species groups involved include darters of the Etheostoma variatum and E. maculatum species groups; the subgenera Ozarka, Swainia, Littocara, Odontopholis, Ericosma, and Imostoma; minnows of the genera Erimystax, Phenacobius, Nocomis, Pimephales; the Notropis zonatus-coccogenis, N. leuciodus, N. telescopus, N. spectrunculus, N. venustus, and N. xaenocephalus species groups; and the sucker genus Hypentelium. Single species illustrating this pattern include the topminnow Fundulus catenatus, the darter Etheostoma blennioides, the sculpin Cottus carolinae, the cavefishes Typhlichthys subterraneus, Ambylopsis, and others listed by Mayden (in press).
Figure 3--Distributions of central highland fishes (dark lines) and composite of maximum glacial advances superimposed on present-day drainage patterns in North America, identifying areas of endemism, northern disjunct populations of some central highland fishes, and the impact of glaciation on the previous widespread ichthyofauna. Figure modified from Mayden (in press).
With endemic faunas found in one or more of the highland regions, ichthyologists have noted a faunal pattern unique to this area and in need of explanation. Also of interest, but rarely commented on, are populations of some species in the central and western Plains region, an area west of the interior highlands.
For many years the explanation advanced for the observed distribution pattern in the central highlands was one of centers of origin and dispersal, commonly proposed by advocates of the school of evolutionary biogeography. Given the diverse nature. of the eastern highlands, most species were thought to have evolved here and dispersed to the other two areas where they would become isolated and perhaps differentiated. Given that this could have happened several times, we would then have the diverse and endemic faunas of the regions. This explanation was not completely without merit or logical basis. Proponents believed that species dispersed from east to west or west to east along the front of an ice sheet during the Pleistocene. Alternately, dispersal could have occurred at some stage during the Pleistocene when the base level would have been lowered enough to give the Mississippi River in southern Illinois and Missouri the high-gradient characteristics required for the survival and successful movement of highland species. Such an explanation was used to explain dispersal of surface species as well as subsurface cavefishes. For the latter, drainages beneath the Mississippi were hypothesized to have connected the two major highland areas. Thus, under this model the present diversity of the central highlands has been in existence since the Pleistocene.
More recently, an alternative hypothesis, that of vicariance, was proposed by Mayden (1985, in press), Wiley and Mayden (1985), and, in part, by Pflieger (1971) as a general explanation of this pattern of distributions and species relationships. These authors proposed that the present highland regions and faunas represent the remnants of a fragmented, once-widespread highland region. They proposed that prior to the Pleistocene the highlands were all interconnected and drained by river systems different from those today. These drainages contained a diverse fauna that was dissected by advancing glaciers and associated glacial processes to produce the existing pattern. Thus, a single general explanation is employed to explain the observed patterns, not multiple dispersals in various directions. Further, this hypothesis is testable. If closest relatives of species in one of the highland regions are not in another highland area, but are outside of the area, then this hypothesis may be falsified. If, on the other hand, species from one region have their closest relatives in another region, then this hypothesis is corroborated and remains to be falsified.
Hence, we have two potential explanations for the historical biogeography of central-highland fishes, one from each of the two competing philosophies of historical biogeography. One explanation invokes centers of origin and dispersal and a second invokes vicariance. Which, if either, of these hypotheses forms the most parsimonious explanation of the pattern? Which is consistent with the phylogenetic histories of the species and the geological history of the region and supplies us with a general theory? Were these regions ever connected to produce the observed faunal patterns without invoking long-distance dispersal? We may examine the evidence below.
All available data suggest that the three highland areas are very old. The Ozark province itself has been above ground since the Pennsylvanian and had its last episode of uplift in the Eocene or before (Bretz, 1965). The Ouachita Mountains and a large portion of the eastern highlands represent an old geomorphic formation extending beneath the Coastal Plain province. This formation dates to the orogeny of the Appalachian Mountains (Thornbury, 1965). Although these three regions are now independent of each other, it is now known that they are, in fact, remnants of a once more widespread highland region. It is now believed that this region extended north (and perhaps west) of the existing highlands and was dissected by the several glacial advances of the Pleistocene (Mayden, 1985, in press; Wiley and Mayden, 1985). Before the advance of glaciers over north-central North America in the Pleistocene, a highland topography and presumably streams with highland characteristics ranged from the Appalachians to at least the Great Lakes region, south to the Coastal Plain, and west to eastern Kansas, perhaps even farther.
The Central Lowlands province is a result of this glacial advance south to the present highland areas. Prior to the Pleistocene, the topography of this area was presumably like that of the existing central highlands. Studies of preglacial bedrock topography from Ohio, Illinois, and Indiana, all areas now in the central lowlands, indicate that the Tertiary geomorphic history of the areas paralleled that of the central highlands to the south (Ver Steeg, 1934, 1936; Hornberg, 1950; Wayne, 1952, 1956). Further supporting the theory of a once-continuous highland region are studies of erosional surfaces from driftless areas throughout the central lowlands which support an expansive highland region (Thornbury, 1965). With the glacial advance, much of the pre-Pleistocene highland topography was destroyed and converted into the existing topography characterized by relatively low relief.
The impact of Pleistocene glaciation on highland habitats goes beyond the destruction of the northern highland areas. Two major events may be attributed to the Pleistocene glaciation. The first is the direct result of moving ice and the second is more indirect, related to sea-level fluctuation. First, advancing glaciers allowed total separation of the large eastern highlands from the western interior highlands, and second it divided the interior highlands into the Ozark and Ouachita provinces (Mayden, 1985, in press). Because of the predominate shape of glacial fronts (Flint, 1957), with maximum advance near the Mississippi Valley, the multiple glacial advances, together with the northward extension of the Coastal Plain province in the Mississippi Valley, effectively severed the highland area into eastern and western regions (fig. 2). A small highland "bridge" in southern Illinois (Thornbury, 1965) still remains, but connection between the two regions is prohibited by the Mississippi River and associated floodplain. Separation of the interior highlands into the Ozark and Ouachita regions was made possible with fluctuating sea level and the development of the Arkansas River system (Quinn, 1958). This event occurred in the Pleistocene and has been minimally dated as pre-Sangamonian.
In summary, although our reconstructed knowledge of past geological events is weak, we may infer from a collection of geological data that the present central highland regions were once part of an old, widespread highland area dating at least to the Eocene. This region was dissected by events of Pleistocene glaciation. First, it was split into two areas, one east and one west of the Mississippi River. The western region later became dissected with the development of the Arkansas River whose floodplain produced lowland habitats similar to those of the central lowlands and Mississippi Valley (see fig. 5 in Mayden, 1985).
Using methods of phylogenetic systematics outlined by Hennig (1966), the historical relationships of species in the central highlands may be examined. This method employs only shared-derived characters to infer relationships within monophyletic (single origin) groups and is superior to other methods when answers to genealogical questions are being sought (Wiley, 1981). Primitive characteristics are those inherited by species through ancestry, are found also outside the group of interest, and are not characteristics indicative of a unique ancestry.
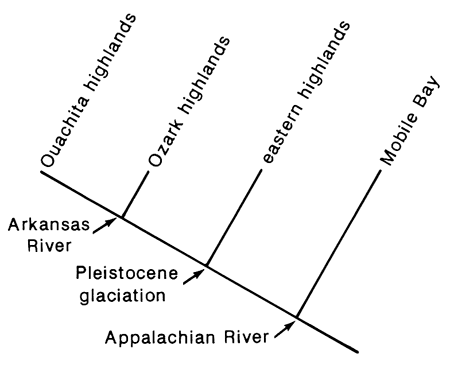
Species relationships in the highland regions do, in fact, suggest a common pattern but not a single pattern. Stated briefly, in the most frequently occurring pattern (fig. 4), species in the highlands of the upper Gulf Coastal drainages form the sister group to the remaining highland species and thus are the result of the oldest speciation event for the groups. This event is presumably correlated with the breakup of the Appalachian River (Starnes and Etnier, 1985; Mayden, in press). Next, species of the eastern highlands form the sister group to species of the interior highlands (fig. 4). This relationship is slightly more complicated than appears in that it is not always a species from the same drainage which is most closely related to the western members. This topic will be addressed more fully below. Within the interior highlands, if a species of a central highland clade is present in the Ouachita highlands, its closest relative is in the Ozark highlands. Thus, we have a generalized temporal sequence of speciation events for central-highland fishes (fig. 4) which is consistent with what we know of the geological history of the area. Some potential problems of extinction in the eastern highlands remain, however, and will be addressed below.
Figure 4--Area cladogram for major central highland regions.
Other biogeographic tracks are observed for central-highland fishes. These patterns are much less common, are not inconsistent with the general Pleistocene-central-highland hypotheses presented here, and need only be listed. Some species of the Ouachita highlands have their closest relatives to the south of the mountains in the Coastal Plain province from eastern Texas to the Atlantic slope. Others have sister species with a widespread distribution over eastern North America (Mayden, 1985). Some species of the eastern highlands have their closest relatives in Atlantic-slope drainages east of the Appalachians (Mayden, in press). In all probability, other patterns will be elucidated once relationships of other highland species are determined, but the temporal sequence of highland areas listed above will remain a common pattern.
Another frequently observed pattern for these highland fishes, relating to the most common pattern mentioned above, is that some species with the common pattern also have disjunct populations to the north of the Ozarks or eastern highlands in the central lowlands (fig. 3; Mayden 1985, in press). The origin of some of these populations may be debatable, but for others it is not. Some are found in driftless areas, regions presumably never glaciated and represent glacial relicts. Others are found in regions previously covered with at least one continental ice sheet. These populations either represent refugia left over from the Pleistocene, or they represent long-distance dispersal after the last glaciation. Because of the disjunct nature of the appropriate habitat of many of these refugia, separate from the nearest highland region by adverse environments for highland species, dispersal seems an unlikely explanation for the origin of these populations unless, however, these species dispersed north following the last melting ice sheet and settled into preferred areas. Alternatively, they may actually represent northern relict populations of species left by glaciation, like Notropis nubilus (fig. 5) and Campostoma oligolepis in the driftless region, and are indicative of a diverse pre-Pleistocene fauna (Mayden, 1985). Because each glacial advance left many areas unglaciated, considering the potential for many "driftless" areas to be present with each advancing glacier is not unreasonable.
Figure 5--Distributions and phylogenetic relationships of species in the Notropis leuciodus group (see Mayden, in press, for species relationships). The solid circle in southwestern Kansas represents fossil locality for N. nubilus.
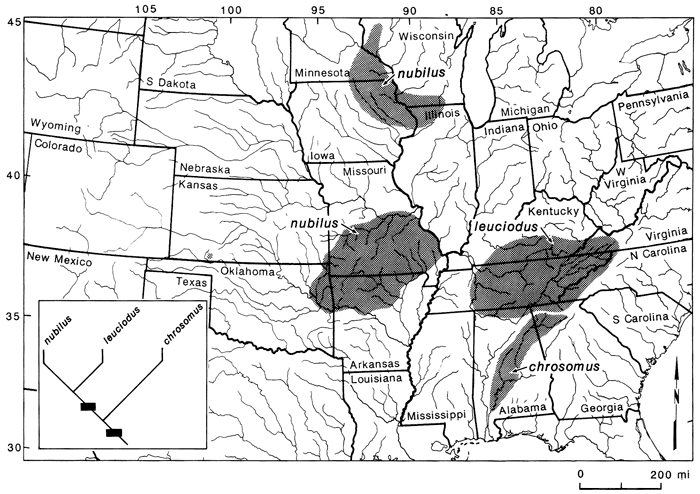
Notropis nubilus is an important species in the overall picture of the biogeography of central-highland fishes (fig. 5). The significance of this species includes its relationships to other members of the N. leuciodus species group, a unique morphological trait possessed by the species, and its distribution. N. nubilus represents one of the best supporting examples of an intact pre-Pleistocene fauna in the highlands.
The Ozark minnow is found in the Ozark highlands, the driftless region, and as a fossil in southwestern Kansas. Populations of this species have a derived coiled intestine, a characteristic unique to this group. Thus, the species evolved this morphological characteristic at the time of the speciation event responsible for its existence. Relationships of N. nubilus are with N. leuciodus of the eastern highlands, and these two together are most closely related to N. chrosomus of the upper Mobile Bay drainage (fig. 5). The significance of these particular findings is that if the populations of N. nubilus present in the northern unglaciated region represent Pleistocene relicts, then the origin of the species must predate the Pleistocene, because both isolated recent populations have the unique coiled intestine. Other species, some with disjunct northern populations, have distributions and relationships also in support of an intact pre-Pleistocene fauna (see Mayden, 1985, in press).
As seen above, the predominate pattern of phylogenetic relationships of central-highland fishes suggests that closest relatives of a species found in one highland region are with species in another highland region. Closest relatives of these highland fish groups are located outside the central highlands. Further, the temporal sequence of speciation events of some groups suggests that the fauna was diverse, widespread, and in existence before the Pleistocene. Distribution patterns and relationships of all other highland species are consistent with this hypothesis. The impact of the Pleistocene glaciation on the central-highland fishes was tremendous but probably had little significance with respect to speciation of these fishes; the only exception being the division of the interior highlands province by the Arkansas River. This event is correlated with the evolution of closest relatives in the Ouachita and Ozark highlands (Mayden, 1985). Massive continental ice sheets were responsible for the vicariance between the interior and eastern highlands, but events responsible for species patterns between the two regions predate the Pleistocene in river systems only partially reconstructed today (see Hocutt and Wiley, 1985).
The hypothesis of centers of origin and dispersal seems unjustified in light of the phylogenetic and geological data presented above and in previous papers. It is true that this explanation can account for all of the observed patterns; it has the potential to account for any and all patterns. However, it is not a general explanation. As mentioned earlier, the center of origin and disperal hypothesis is difficult, if not impossible, to test. Acceptance of such an explanation would result in the loss of valuable information to geologists and organismal biologists concerned with historical events occurring over a landscape of North America only partially reconstructed today. This hypothesis seems to be further unjustified in light of Pleistocene findings presented by Dort (1983). Presently no data are available to support the hypothesis that base level was lowered enough during the Pleistocene to alter the habitat characteristics of the Mississippi River near southern Illinois (see Mayden, 1985).
As mentioned previously and discussed by Cross (1970), disjunct populations of some species typically thought of as ozarkian are found in the Plains region west of the interior highlands in central and western Kansas and eastern Colorado. The question is, do these populations represent independent dispersal events westward into the Central Plains, or could this region have been at one time more favorable for highland fishes, such that the interior-highland fauna primarily extended farther west? Geologically we have little information from this area concerning previous environmental conditions. However, using historical biogeography, can we make predictions as to the origins of these populations?
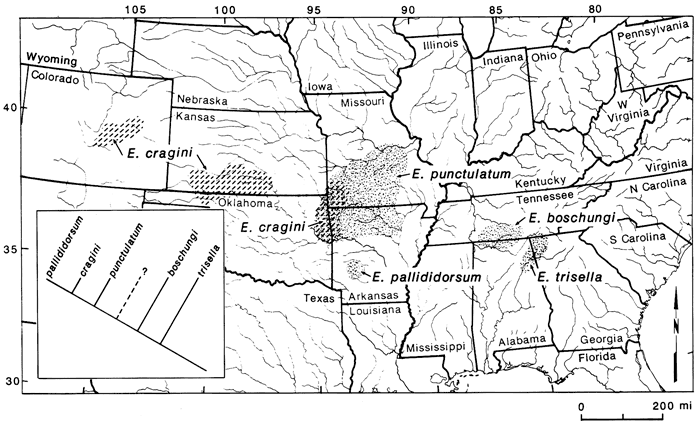
Species of primary concern here include the minnows Phoxinus erythrogaster, Nocomis asper, Notropis pilsbryi, N. nubilus, Hybopsis amblops, and the darters Etheostoma flabellare and E. zonale (fig. 6). These species are common in the Ozark highlands and have disjunct populations in the Flint Hills region of central Kansas. Other taxa to be considered here include the minnows Erimystax x-punctata and Notropis camurus, the madtom Noturus placidus, and the darter Etheostoma cragini (fig. 6). These species have populations in central Kansas and some species are known from western Kansas and eastern Colorado. Erimystax x-punctata and Noturus placidus extend only up the Neosho River, but Notropis camurus and E. cragini are distributed farther west. These species differ from those listed above in that their distributions are more continuous and that recent populations exist between the central and western populations and the Ozarks, although disjunctions between the populations are common. So, we may ask if there is a general explanation for the distributions of these species in the Central Plains region.
Figure 6--Disjunct populations of fishes with highland habitat preferences in the Central Plains region. Stippled pattern represents area of co-occurrence of Ozark species with western disjunctions. Area of Neosho River in Kansas enclosed by shaded area demarks distributions of Noturus placidus and Erimystax x-punctata.
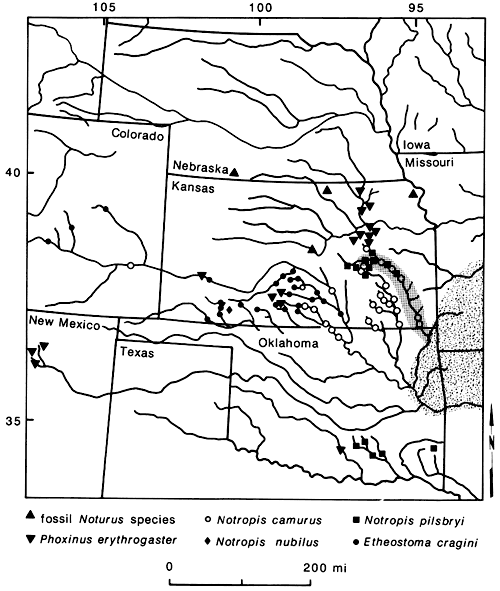
For Notropis camurus, Noturus placidus, and Erimystax x-punctata, dispersal from the Ozarks to some western regions in recent times seems plausible. Individuals of these species can be found in the Neosho River. Thus, the river apparently is not a barrier to their movement. Dispersal to areas further west seems improbable, however, because of adverse habitats in intervening areas. For the remaining taxa, habitat restrictions preclude their long-distance dispersal within historic time. The present distributions of these species suggest a previously more widespread distribution of the taxa with later extinction. This was discussed in more detail for some species by Cross (1970).
From these data the most parsimonious explanation is that a highland environment must have extended farther west than the existing interior highlands at some time in the past, at least in the Pleistocene, if not before. Geological data are weak for reconstructing this history, but fossil data add some insight. Late Pleistocene fossils of Notropis nubilus are known from southwestern Kansas (Meade County), indicating that at least at this time conditions were favorable here for this species (Smith, 1963; Cross, 1970) and perhaps other highland taxa. Further supporting a more widespread highland environment are several Pleistocene fossils of a Noturus species in the furiosus group reported by Eshelman (1975), Neff (1975), Bennett (1979), and Cross et al. (1985) from Nebraska and Kansas (fig. 6). Four species are presently in this group, the closest member geographically being N. placidus. These fossils support a more widespread distribution of the furiosus group in the Pleistocene, whether or not they are eventually determined to be N. placidus.
Thus, based on fossil materials, distributional data, and environmental restrictions of recent populations of these species, we may predict that these species and possibly others were more widely distributed west of the interior highlands before and during the Pleistocene. In light of present-day habitat selectivity of many species, especially those of the central highlands, we may predict that this region also was highland in character. Cross (1970) reached a similar conclusion concerning some of these species, remarking that the region of the central plains must have had more water and that the streams would have been cooler. These characteristics, combined with clear streams would probably have been acceptable to many highland fishes.
Given the pattern of isolated populations of highland fishes in the Central Plains region, the differential in numbers of species present in the central-highland regions and those present in the glaciated central-lowland province, together with the hypothesis that the central highlands were once continuous with an intact fauna before the Pleistocene speculating that extinction occurred during the Pleistocene is not unreasonable. This is not a new idea. Previous researchers have hypothesized that extinction (species and/ or populations) during the Pleistocene was a real phenomenon. The magnitude of this natural process is, however, very difficult to determine.
Examination of distribution patterns and available fossil data of the Plains fishes listed above indicate extinction at least on the population level and perhaps at the species level (e.g., Noturus sp.) of some taxa occurring in this region during the Pleistocene and/or later (also see Cross, 1970). The same may be said for populations of highland fishes which previously inhabited areas north of the interior and eastern highlands. However, at present we have no fossil data to suggest that species extinction occurred in the central lowlands during the Pleistocene. Is this really the case? I would suggest not. Based on distributions of extant fishes and their phylogenetic relationships, it seems very likely that some species extinctions probably occurred during the Pleistocene, never to be recorded in the fossil record. As mentioned earlier, the closest relative to interior-highland fishes is an eastern-highland species, but the distribution of this taxon is variable. The species may be from any or all of the three major drainages, the Ohio, Cumberland, and Tennessee rivers. This pattern, together With species relationships, may be an indication that extinction has occurred in fish groups of this region. We may examine the evidence below.
The primary basis for the extinction statement involves the distributions and relationships of members of the Etheostoma variatum group relative to other species groups found in the highlands. Members of the E. variatum group are limited to the central highlands (fig. 7). Two species are in the interior highlands, four are in the eastern highlands, and one is endemic to the northern Atlantic slope. Etheostoma blennius is the sister to all members of the group and is found in the Tennessee River system. The two interior-highland species form the sister group to the three members in the Ohio River system and E. sellare of the east coast is most closely related to these five taxa (fig. 7). If we compare this group with other central-highland groups, we can see where extinctions may have occurred. Examples include the Notropis zonatus-coccogenis group of the subgenus Luxilus (fig. 8) and the subgenus Ozarka of Etheostoma (fig. 9). In these groups we see that the Tennessee representatives are present in all three, as well as the interior-highlands representatives. However, in the latter two groups, Ohio River and northern Atlantic-slope members are missing. One explanation is that they were never present. If so, then one would have to hypothesize long-distance dispersal between the eastern and western highland regions to explain existing relationships. However, given that we have ample data to suggest a previous widespread fauna which was subsequently altered by glaciation, these two groups suggest some degree of extinction has occurred, specifically those members of the subgenera Luxilus and Ozarka that would have been present in the upper Ohio River system. Etheostoma sellare and Notropis cerasinus illustrate fairly unique patterns for central-highland fishes, perhaps peripheral isolates (Wiley and Mayden, 1985), and can essentially be ignored for this comparison. Following Rosen's (1978) work on area cladograms of Central American fishes, the significant comparisons here are those common to both the interior and eastern highlands. If E. variatum, E. kanawhae, and E. osburni (fig. 7) were to be removed from their phylogeny, relationships in the variatum group would be like those of the zonatus group (fig. 8) and the subgenus Ozarka (fig. 9). Thus, with reference to the phylogenies of these groups (figs. 7-9), it is possible that other species were present between the interior-highland members and those in the Tennessee River system. This scenario also may be the case for several other species groups (see Wiley and Mayden, 1985; Mayden, in press). If true, extant taxa in the two highland regions may represent apparent rather than actual sister groups.
Figure 7--Distribution and relationships of species in the Etheostoma variatum group (see Wiley and Mayden, 1985, for species relationships). Etheostoma sellare of the Atlantic Coast is not included.
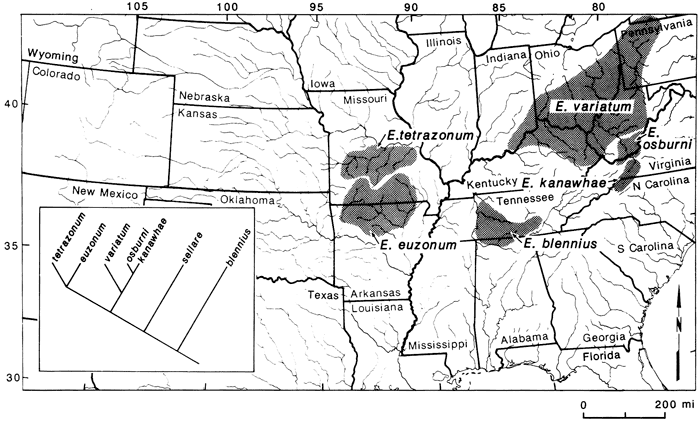
Figure 8--Distributions and relationships of species in the Notropis zonatus-coccogenis species group (see Mayden, in press). Dashed line in phylogenetic hypothesis represents proposed extinct taxa or taxon. Notropis cerasinus of the Atlantic Coast is not included.
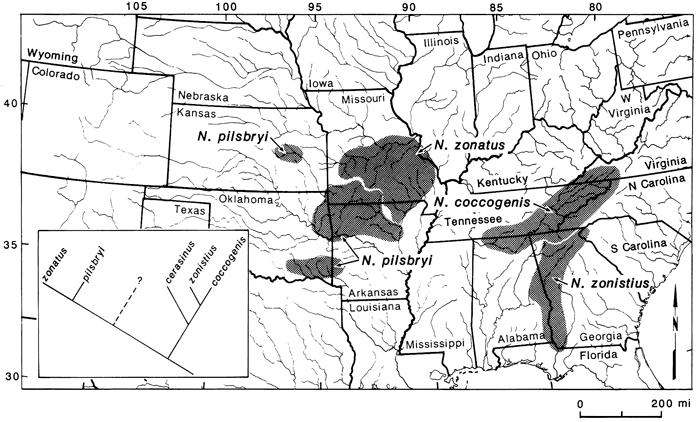
Figure 9--Distributions and relationships of species in the subgenus Ozarka (see Mayden, in press, for species relationships). Dashed line in phylogenetic hypothesis represents proposed extinct taxa or taxon.
The frequency of missing representatives of Ozark sister species in the Ouachita highlands deserves comment. Although extinction via glaciation cannot be directly attributed to this pattern since the Ouachita highlands were never glaciated, one would predict, based on data from other groups, that more species related to Ozark taxa should be present in the Ouachita highlands. Two possible explanations seem obvious. First, the existing Ouachita sister taxa may represent independent dispersal events from the Ozarks across the Arkansas River valley and not a vicariant event. This seems rather unlikely since these two areas were once a continuous region dissected by the Arkansas River in the Pleistocene (Quinn, 1958) and the present-day Arkansas River floodplain contains unfavorable habitats for highland fishes. Alternatively, the "missing" elements represent examples of extinction due to the small size of the region, relative to the size of other highland areas. Extinction rates are known to be greater on smaller oceanic islands than on larger islands (MacArthur and Wilson, 1967). Whether the principles of island biogeography apply to these highland "islands" has yet to be demonstrated.
Bennett, D. K., 1979, Three late Cenozoic fish faunas from Nebraska: Transactions, Kansas Academy of Science, v. 82, p. 146-177.
Bretz, J. H., 1965, Geomorphic history of the Ozarks of Missouri: Missouri Geologic Survey and Department of Water Resources, v. XLI.
Brundin, L., 1966, Trans-Antarctic relationships and their significance: Kungliga Svenska Vetenskapsakademiens Handlingar, v. 4, no. 11, p. 1-472.
Brundin, L., 1972, Phylogenetics and biogeography: Systematic Zoology, v. 21, p. 69-79.
Croizat, L., 1952, Manual of phytogeography: Vitgeverig Dr. W. Junk, The Hague.
Croizat, L., 1958, Panbiogeography: L. Croizat, Caracas, Venezuela.
Croizat, L., 1964, Space, time, form--the biological synthesis: L. Croizat, Caracas, Venezuela.
Cross, F. B., 1970, Fishes as indicators of Pleistocene and Recent environments in the Central Plains; in, Pleistocene and Recent Environments of the Central Great Plains: University of Kansas Press, Lawrence. Kansas, Special Publication No. 3., p. 241-257.
Cross, F. B., Mayden, R. L., and Stewart, J. D., 1985, Fishes in the western Mississippi drainage; in, Zoogeography of North American Freshwater Fishes, C. H. Hocutt and E. O. Wiley, eds.: Wiley Interscience, p. 363-412.
Dort, W., Jr., 1983, Sea-level changes and midcontinental fluvial terraces (abs.): Symposium on Late Cenozoic Continental Glacial, Periglacial, and Marine Correlations in the Great Plains-Gulf Coast, Lincoln, Nebraska.
Dowling, H. G., 1956, Geographic relations of ozarkian amphibians and reptiles: Southwestern Naturalist, v. 1, p. 174-189.
Eshelman, R. E., 1975, Geology and paleontology of the Early Pleistocene (Late Blancan) White Rock fauna from north-central Kansas: University of Michigan Museum of Paleontology, Papers in Paleontology, v. 13, p. 1-60.
Flint, R. F., 1957, Glacial and Pleistocene geology: John Wiley and Sons, New York.
Hennig, W., 1960, Die Dipteran-Fauna von Neuseel als systematisches un tiergreographisches Problem: Beiträe Zur Entomologie, v. 10, p. 221-329 (English translations, 1966, Pacific Insects Monographs, v. 9, p. 1-81).
Hennig, W., 1966, Phylogenetic systematics: University of Illinois Press, Urbana.
Hocutt. C. H., and Wiley, E. O., eds., 1985, Zoogeography of North American freshwater fishes: Wiley Interscience, New York, 866 p.
Hornberg, L., 1950, Bedrock topography of Illinois: Illinois Geological Survey, Bulletin 73.
MacArthur, R. H.. and Wilson, E. O., 1967, The theory of island biogeography: Princeton University Press, Princeton.
Mayden, R. L., 1985, Biogeography of Ouachita highland fishes: Southwestern Naturalist, v. 30, no. 2, p. 195-211.
Mayden, R. L., in press, Biogeography of North American central-highland fishes--a research program in community ecology; in, Community and evolutionary ecology of North American stream fishes, W. J. Matthews and D. C. Heins, eds.: University of Oklahoma Press, Norman, Oklahoma.
Metcalf, A. L., 1966, Fishes of the Kansas River system in relation to zoogeography of the Great Plains: University of Kansas, Museum of Natural History, Publication 13, p. 23-189.
Neff, N. A., 1975, Fishes of the Kanopolis local fauna (Pleistocene} of Ellsworth County, Kansas: University of Kansas Museum of Paleontology, Papers in Paleontology, v. 12, p. 39-48.
Nelson, G., and Platnick, N. I., 1980, A vicariance approach to historical biogeography: Bioscience, v. 30, no. 5, p. 339-343.
Nelson, G., and Rosen, D. E., eds., 1981, Vicariance biogeography, a critique: Columbia University Press, New York, 593 p.
Platnick, N. I., and Nelson, G., 1978, A method of analysis for historical biogeography: Systematic Zoology, v. 27, p. 1-16.
Pflieger, W. L., 1971, A distributional study of Missouri fishes: University of Kansas, Museum of Natural History, Publication 20, no. 3, p. 226-570.
Quinn, J. H., 1958, Plateau surfaces of the Ozarks: Proceedings of the Arkansas Academy of Science, v. 11, p. 36-43.
Rosa, J., 1918, Ologenesi: Bemparad, Firenze.
Rosen. D. E., 1978, Vicariant patterns and historical explanations in biogeography: Systematic Zoology, v. 27, p. 159-188.
Smith, G. R., 1963, A late Illinoian fish fauna from southwestern Kansas and its climatic significance: Copeia, v. 1963, p. 278-285.
Starnes, W. C., and Etnier, D. A., 1985, Drainage evolution and fish biogeography of the Tennessee and Cumberland rivers drainage realm; in, Zoogeography of North American Freshwater Fishes, C. H. Hocutt and E. O. Wiley, eds.: Wiley Interscience, New York, p. 325-361.
Thornbury, W. D., 1965, Regional geomorphology of the United States: John Wiley and Sons, New York.
VerSteeg, K., 1934, The buried topography of north-central Ohio and its origin: Journal of Geology, v. 42. p. 602-620.
VerSteeg, K., 1936, The buried topography of western Ohio: Journal of Geology, v. 44, p. 918-939.
Wayne, W. J., 1952, Pleistocene evolution of the Ohio and Wabash valleys: Journal of Geology, v. 60. p. 575-585.
Wayne, W. J., 1956, Thickness of drift and bedrock physiography of Indiana north of the Wisconsin glacial boundary: Indiana Geological Survey, Progress Report 7.
Wiley, E. O., 1981, Phylogenetics--the theory and practice of phylogenetic systematics: John Wiley and Sons, New York, 439 p.
Wiley, E. O., and Mayden, R. L., 1985, Species and speciation in phylogenetic systematics, with examples from the North American fish fauna: Annals of the Missouri Botanical Gardens, v. 72, p. 596-635.
Prev--Wisconsinan sediments on the High Plains of Kansas || Next--Wisconsinan Mammalian Faunas
Kansas Geological Survey
Comments to webadmin@kgs.ku.edu
Web version updated March 28, 2010. Original publication date 1987.
URL=http://www.kgs.ku.edu/Publications/Bulletins/GB5/Mayden/index.html