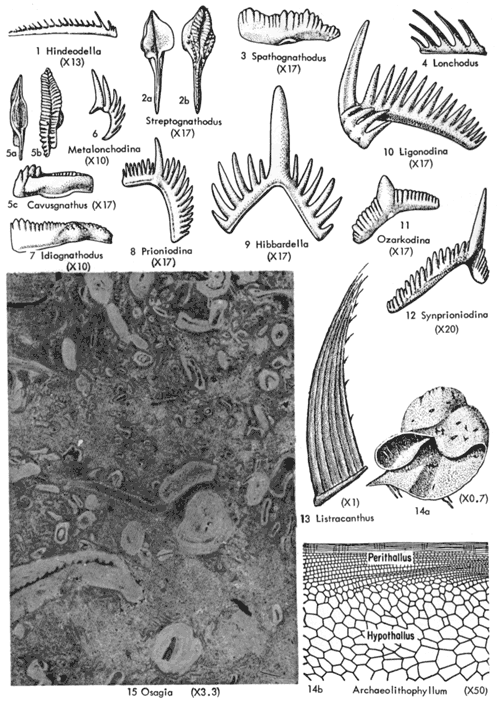
|
Original published in D.F. Merriam, ed., 1964, Symposium on cyclic sedimentation: Kansas Geological Survey, Bulletin 169, pp. 287-380 | |
University of Kansas, Lawrence, Kansas
[Please Note--An Acrobat PDF file has been created containing higher-quality versions of some of the figures in this paper (Acrobat PDF, 1.9 MB). All the large-size figures are in this file; it only needs to be downloaded once though it is linked to each figure in the article.]
The recognition of more than 100 cyclothems in the Pennsylvanian and Lower Permian rock succession of Kansas, each containing a number of distinctive types of deposits and varied assemblages of organic remains, provides opportunity for paleoecological observations and interpretations which may come to be accepted as specially trustworthy. This is because repetitive occurrences of identical or near-identical sedimentary environments and biotopes can be analyzed comparatively and because sequential relationships of these environments and biotopes within the different cyclothems aid in confirming interpretations. Traceability of thin layers for long distances along outcrops--some more than 200 miles--allows study of geographic constancy of individual biotopes or departure from constancy. Approximately 20 representative ecologic communities (ecosystems) are characterized briefly and their stratigraphic occurrence is illustrated diagrammatically, though very incompletely.
The purpose of this paper is to point out characters of Pennsylvanian and Permian cyclothems in Kansas which specially merit detailed paleoecological analysis. The characters to be discussed are not newly discovered, for most of them have been recognized since cyclic sedimentation was first described (Moore, 1929, 1930, 1931; Weller, 1930; Elias, 1934, 1937a) in parts of the Kansas Pennsylvanian-Permian rock column and a little later found to be clearly determinable in almost all parts of the succession. The dissimilar environments of deposition represented by coal beds, slabby sandstones and thin clayey limestones bearing tetrapod skeletal remains and footprints, sandy to clayey shales containing well-preserved parts of land plants, and strata characterized by assemblages of marine invertebrates were selfevident to early interpreters of the cyclothems, at least in general qualitative terms. To some extent different kinds of marine assemblages were discriminated and it was observed that structures interpreted as algal in origin seemed to be most common or essentially restricted to upper carbonate layers in very many cyclothems.
Observations during the 1930's and 1940's on several divisions of the Pennsylvanian and Permian in Kansas which were considered to display features of cyclic sedimentation were chiefly directed to discrimination of significant stratigraphic units and to determining their continuity from place to place across the state. This contributed greatly to precision in establishing correlations both within the borders of Kansas and between outcrops studied in Kansas and many geographically isolated exposures in southeastern Nebraska, northwestern Missouri, and southwestern Iowa. Areal stratigraphic work was carried in generally successful manner southward into Oklahoma, where considerable change in sedimentary facies occurs. By and large, the paleoecologic implications of different parts of the recognized cyclothems were given scant attention during this period, if not ignored. In regional perspective, eastern Kansas could be depicted reliably as a stable platform area which repeatedly was submerged shallowly by invading seas, whereas the Oklahoma-Arkansas country characterized by thick clastic deposits of Pennsylvanian and Permian age was prevailingly a wide downwarped trough.
The broad regional nature of Pennsylvanian and Permian sedimentary accumulations in the northern Midcontinent--that is, in territory north of the southern Oklahoma mountains--shows correlation with the kinds and lateral extent of biotopes which successively came to exist in different areas. In general, terrestrial environments such as typically belong to nearly flat coastal plains predominated south of the Kansas border and during much of Desmoinesian, early Missourian, and early Virgilian time within southern Kansas. Associated marine deposits tend to be local and variable, reflecting temporary submergences of parts of the plains. To the north and west, shallow seaways, each characterized by great extent of several sorts of marine environments which were developed successively, spread out again and again over the large crustal tract designated as stable platform. A remarkably even layer-cake stratigraphic record is the product of bed-on-bed sedimentation precisely balanced by regional crustal subsidence. Similar sorts of biotopes have exceptional geographic extent and one follows another in what must have been relatively very short intervals of geologic time (Moore, 1962).
Paleoecology, which may be defined as the study of all aspects of relationships between ancient organisms--essentially those preserved as fossils--and their environment, has borrowed from ecology, concerned with present-day organisms and their environments, a number of special terms. Depending on context, these may or may not warrant modification by adding the prefix "paleo-," and many of them can be dispensed with for general purposes. If it is needful to distinguish between populations of ancient organisms growing together from accumulations of ancient organic remains possibly transported from various places of origin and merely buried together, classification of them as life assemblages (not paleobiocoenoses) and death assemblages (not paleothanatocoenoses) is adequate. Divisions of paleoecology into studies devoted to individual organisms or small groups of taxonomically defined organisms preserved as fossils (paleoautecology) and those concerned with ancient organic communities (paleosynecology) are useful in textbooks (Ager, 1963) but otherwise are little required.
In the present paper only two or three special terms, possibly not familiar to everyone, are rather commonly employed. These are (1) ecosystem, defined as any sort of ecologic community considered as a unit in relation to nonliving factors of its environment, and (2) biotope or paleo biotope, used to designate a region of unspecified size which is characterized by essentially uniform environmental conditions and by a correspondingly uniform population of animals or plants or both animals and plants. The word "paleoecosystem" is thought to be superfluous, and possibly the same is true of "paleobiotope" in discussion of Pennsylvanian and Permian biotopes. Nevertheless, in various connections paleobiotope seems preferable to biotope. I find no need for biome, defined by ecologists as a major natural region characterized by certain groups of organic communities, or its paleoecological equivalent, paleobiome.
Cordial appreciation and my thanks are expressed to Daniel F. Merriam and Theodore E. Jacques for assistance in field studies of Lower Permian (Beattie) deposits in southern Kansas and northern Oklahoma, both before and during days spent with John Imbrie and Léo F. Laporte for the same purpose in early August, 1964. Also, I am indebted to Frank M. Carpenter, of Harvard University, for help in recording fossil insects which to date have been described from the Elmo area of Dickinson County, Kansas. Likewise, I need to acknowledge work done by Norman D. Newell on Pennsylvanian (Missourian) correlation diagrams, which, though prepared many years ago, are published here for the first time. I am especially grateful to John Imbrie for supplying to me parts of the data derived from his detailed investigations of Florena Shale biostratigraphy and for allowing me to publish graphs which I have prepared based on information obtained from him. Finally, I thank Roger B. Williams for careful work done by him in drafting part or all of certain figures.
Organic assemblages, or ecosystems, and the biotopes corresponding to them obviously are divisible into two main groups: (1) terrestrial or nonmarine, and (2) marine. The fact that some environments are neither strictly subaerial in the sense of being located on land (including river, lake, and marsh habitats, as well as those of hot dry deserts, frigid tundras, and many others) nor definitely marine, because situated below sea level in areas subject to flooding by ocean-connected salt water, embarrasses an oversimplified classification but presents no insuperable difficulties for an ecologist. For a paleoecologist, on the other hand, problems in dealing with many ancient assemblages and with many paleobiotopes recorded in sedimentary deposits may be extremely troublesome--so much so that no reasonably firm conclusions are possible. Therefore, a threefold classification of fossil assemblages and their equivalent paleobiotopes is desirable in paleoecologic studies: (1) definitely nonmarine, (2) definitely marine, and (3) intermediate or doubtful, the last-mentioned group including some in which paleontological and sedimentological evidence, combined with that furnished by stratigraphic relations, is deficient or possibly conflicting. Not many paleobiotopes can be treated as "cut and dried."
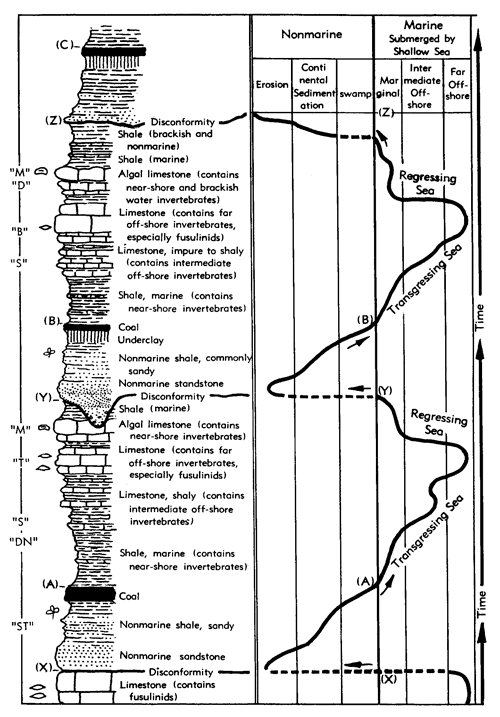
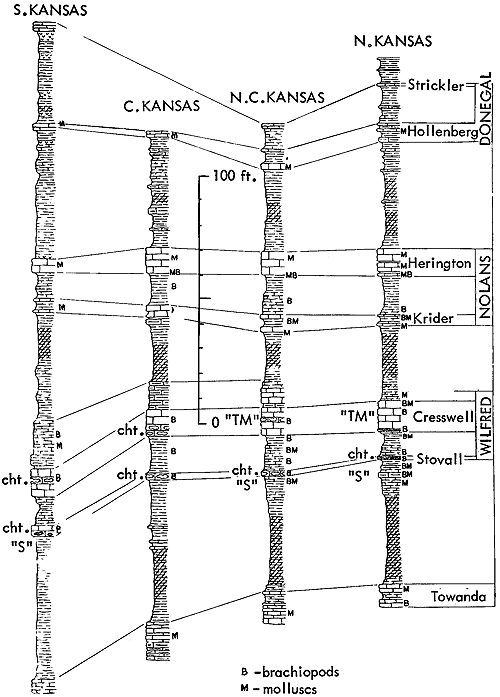
Generalized litho- and biostratigraphic attributes of Pennsylvanian and Permian cyclothems found in the Kansas region are illustrated in Figure 1, which also indicates alternative methods of drawing the boundaries of these cyclic sedimentary units. The stratigraphic section here given is accompanied by indication of roughly classified nonmarine and marine biotopes and interpretation of environments represented by the successive strata is marked by fluctuating position of a sedimentation-time line which shifts back and forth from column to column denoting different biotopes. The positions of some ecosystems, described in later parts of this paper, with explanation of symbols for them used on diagrams, are shown along the left margin of the diagram.
Figure 1--Diagrammatic section of successive cyclothems showing typical litho- and biostratigraphic features, accompanied by interpretation of changes in environments with lapse of time (graph at right). Alternative choices in defining boundaries of cyclothems are indicated by capital letters enclosed in parentheses: (A), (B), (C), boundaries drawn on top of coal beds, coinciding with change from nonmarine to marine conditions of sedimentation; (X), (Y), (Z), boundaries drawn at disconformities. Several sorts of ecologic communities (ecosystems) are indicated by letters along the left margin of the figure, which refer to subsequently described organic assemblages named from selected stratigraphic units in which they are typically developed and from some genus or genera which are distinctive or common constituents. [Explanation of symbols for ecosystems: "B," Beil-type (Pulchratia), "D," Drum-type (Euconospira), "M," Morrill-type (Osagia), "S," Speiser-type (Derbyia), "DN," Doniphan-type (Rhombopora), "ST," Stranger-type (Asterotheca), "T," Tarkio-type (Triticites).]
It is desirable at this point to make note of geographic spread of recognized organic assemblages and paleobiotopes in typical cyclothems of the Pennsylvanian and Permian succession in the Kansas region. A few of these are confined to small areas, being evidently very local, though they may be duplicated at approximately the same horizon in many other places. The great majority of distinguished biotopes, however, are extraordinarily extensive, being traceable without appreciable change for scores of miles along outcrops and some being identifiable far down dip in the subsurface. Many deposits only 10 to 20 inches in average thickness or less than 10 inches in maximum observed thickness have been identified in closely spaced outcrops extending from counties in southeastern Nebraska all of the way to northern Oklahoma, and throughout this distance essential uniformity of physical and organic features is maintained. Continuity and regularity are the rule, not the exception. This is surprising enough in a sequence of marine deposits, for in almost any seaway depth of water and the nature of bottom sediment may be expected to vary appreciably, and in company with such variation, communities of bottom-dwelling organisms should display rather easily detectable differences. Some stratigraphic observations accord with these expectations but very commonly they do not. All Kansas coal beds known to me are inferred to have been formed in a nonmarine environment, although many of them may represent plant accumulations in swamps barely beyond the reach of flooding by seemingly tideless inland seas. Some such coals have maximum known distribution of a few hundreds or thousands of square miles, and some thin beds (less than 6 inches in average thickness) are seemingly continuous along the outcrop for airline distances of 200 miles or more.
The demonstrated great areal distribution of very many, if not all paleobiotopes characteristic of Kansas Pennsylvanian and Permian cyclothems, points to exceptionlly widespread, nearly uniform environments during each episode of sedimentation, and though the nature of these environments underwent changes with lapse of time, such changes seem to have been introduced almost simultaneously everywhere. An important deduction that rests on this great geographic spread of successive paleobiotopes and their prevailing uniformity from place to place while they persisted is general stability of the underlying crust. The Kansas region thus is distinguished as having the character of a broad, relatively very stable platform.
Inherent in concepts of cyclic sedimentation is repetition of various kinds of sedimentation in constant order as recorded at any locality. The changes in environments with lapse of time eventually introduced again and again a particular kind of biotope at places where they had existed before. This is illustrated diagrammatically in numerous sections of Pennsylvanian and Permian rocks published in the present paper, in addition to being well shown in other contributions.
The purpose of writing about repetition of paleobiotopes and various kinds of organic communities in Kansas is to point out the value of classifying them in types which seem to have similar characters and then of comparing the examples of each with one another. Critical analysis which is likely to be enhanced by a multiple attack on problems of interpreting the significance of physical, chemical, and biological attributes of the deposits which comprise records of the different environments, tabulating all observable similarities or identities on one hand and dissimilarities on the other, is best suited to yield trustworthy conclusions. Also, it is a most promising method for the development of paleoecologic principles having much practical value. For example, the cyclic relationships of associated paleobiotopes, when considered in the light of their regional stratigraphic setting, may be a surer guide to correctness in understanding than can be found in seeking modern ecologic equivalents of each observed biotope. Indeed, modern equivalents of certain Pennsylvanian and Permian paleobiotopes may occur nowhere.
Absolutely indispensable for paleoecological studies of regional scope, applicable to areas larger than single localities of small extent, is a firmly established stratigraphic framework, and the value of this framework is enhanced in proportion to details of its content and its reliability. Most Pennsylvanian and Permian deposits found in Kansas so well satisfy the most rigorous requirements of stratigraphic correlations that they furnish one of the best field laboratories in the whole world for making useful paleoecological observations and for developing principles for practical application to other deposits. Numerous figures given in this paper illustrate parts of this stratigraphic framework.
The remainder of this paper is devoted to the characterization of examples of organic assemblages found recurrently in parts of Pennsylvanian and Permian cyclothems of the Kansas region. Each such assemblage, or ecosystem, is indicative of a paleobiotope, some of which differ markedly from all others and some only slightly, even though distinctions among the latter are thought to be significant. Paleontological differences that reflect evolutionary changes and probably also the effects of emigrations and immigrations are intentionally ignored, because interest is centered on the complexion of animals and plants found together as a community in various ecologic conditions.
The almost endless distinguishable organic associations found preserved as fossil groups in Pennsylvanian and Permian rocks of Kansas bring to paleoecologists difficult problems of classifying and designating them, quite apart from the still more difficult problems of interpreting them in trustworthy manner. Reference to successive assemblages in parts of typical cyclothems in such terms as "molluscan faunas," "mixed faunas," "brachiopod faunas," "fusulinid faunas," as by Moore (1936a, p. 22) and Elias (1937a, p. 410) is far from definitive, and two decades later designations such as "molluscoid," "pectinoid" (Lane, 1958, p. 158), "Osagia facies," "shelly facies," "fusuline facies" (Imbrie, Laporte, and Merriam, 1959, p. 73; Laporte, 1962, p. 526), "black shale facies," "algal limestone facies" (McCrone, 1963, p. 65-66) are little if any better. Brachiopods, bryozoans, mollusks, and algae--not mentioning other organisms--are numerous both in kinds and in their ecological associations. Any assemblage of calcareous-shelled invertebrates, including fusulinid foraminifers, is "shelly." Remembering complexities of vertical and lateral variations in organic groups preserved as fossils and changes in both generic and specific composition of seemingly homologous populations with lapse of geologic time, how may we proceed in trying to make useful paleoecological discriminations? Possibly the method of choosing specified samples as types may serve to allow more precise delineation of significant characters and to provide a better basis for recording similarities or contrasts observed when various natural assemblages are compared with the selected types. I have decided to adopt this procedure in the present paper, partly as a general test of its practicability and partly to invite others to suggest any additions and improvements which are considered desirable.
Among many ecological assemblages (surely dozens) that seem worthy of recognition in fossil-bearing strata of the Kansas Pennsylvanian-Permian section approximately 20 are described in this paper at least very briefly and examples of their occurrence are indicated on accompanying stratigraphic diagrams. They are named from a selected stratigraphic unit and one or more of its contained genera in the chosen occurrence of each assemblage considered to be typical, ignoring the facts that the adopted name-giving genera commonly are present in other ecological communities and that they may be lacking in some rock units classed as representing the assemblage. For instance, Triticites is extremely abundant in the local ecosystem (Tarkio) chosen as type of the Triticites assemblage, and very few other organisms are associated with it there. It happens, however, that Triticites occurs also in a half-dozen other, differently named assemblages, and it is unknown, for example, in the upper Cottonwood Limestone, which is cited as an excellent so-called Triticites assemblage. This serves to illustrate the point that (in usage of the present paper) the adopted name of each ecosystem is merely a nomenclatural handle, which mayor may not signify what it seems to mean concerning occurrence of the name-giving genus.
Symbols for ecologic assemblages given on stratigraphic diagrams are capital letters enclosed by quotation marks. These are initial letters of rock units selected as the type exampIes in occurrence of the assemblage, thus tying it to a particular horizon and generally a specified locality. The index letter (s) is not derived from the name-giving genus of the assemblage (e.g., "B" here refers to the populous and highly varied ecosystem called Pulchratia assemblage found typically developed in the Beil Limestone Member of the Lecompton Limestone near Lecompton, Kansas). The letter symbols adopted for some assemblages, however, may be initial letters of both the selected stratigraphic unit indicated as showing type occurrence and the name-giving genus (e.g., "T" for Tarkio Limestone containing the Triticites assemblage). Several assemblages are indicated by symbols along the left margin of Figure 1.
The following descriptions of ecologic assemblages are brief--perhaps too brief. They are aimed at pointing out main distinguishing features, with little discussion generally of their paleoecological significance.
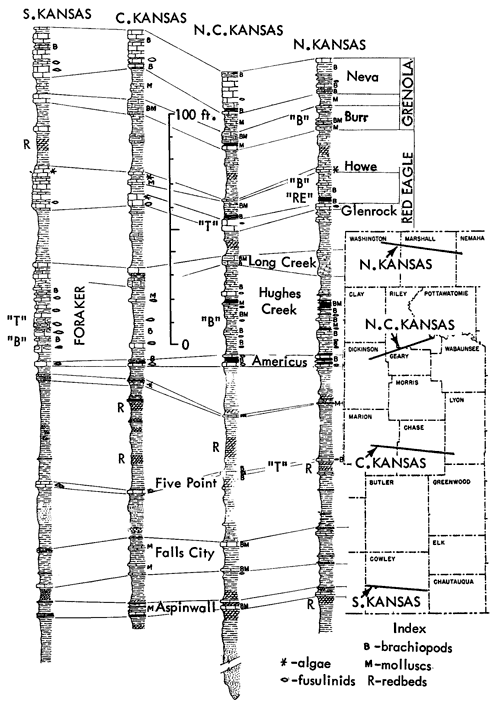
Examination of the characters of generalized simple cyclothems may be followed by calling attention to what seem to be repeated groups of more or less dissimilar cyclothems arranged in constant vertical succession. Such groups, which have been called megacyclothems (Moore, 1936a, p. 29), were first distinguished in the Shawnee part of the Virgilian Stage of the Kansas Upper Pennsylvanian and later recognized (with some undeveloped elements) in the Missourian and upper Desmoinesian successions (Fig. 2). Still later (Moore and Merriam, 1959, p. 28; Merriam, 1963; p. 105) megacyclothems have come to be delineated in the Lower Permian part of the Kansas column (Fig. 3). The distribution of various kinds of paleobiotopes in megacyclic sequences is a subject which has not yet been studied critically. This needs to be done, for seemingly it is essential to solving the origin of megacyclothems.
Figure 2--Cyclothems grouped in megacyclothems, as observed in late Missourian and early Virgilian deposits of central Kansas. Diagram emphasizes repetition of strikingly dissimilar cyclothems in constant sequence. (Moore). [Included in the Acrobat PDF file of figures for this article.]
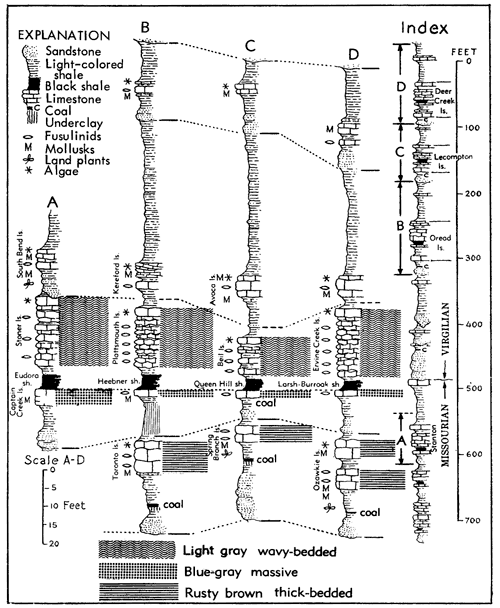
Figure 3--Cyclothems and megacyclothems in Lower Permian rock successIon of Kansas. Upper sections are accompanied by graphic representation of inferred marine transgressions (culminating phases indicated by parts of curve reaching farthest left) interrupted by sea withdrawls when sedimentation of terrestrial type prevailed (modified from Elias by notation of inferred megacyclothems, A-F). Lower part, of diagram shows correlation of cyclothem elements belonging to megacyclothems A-F (Moore in Moore and Merriam). [Included in the Acrobat PDF file of figures for this article.]
All sorts of lands plants are here grouped arbitrarily together under the designation of Asterophyllites assemblage (Suppl. Fig. 1, 1), without making effort to separate groups of swamp-dwelling hydrophytes, dry-climate xerophytes, and intermediate plants classed as mesophytes. Instead, ferns, pteridosperms, sphenophylls, calamites, lycopods, cordaites, and coniferlike plants are lumped together, because the paleoecologic significance attached to them in this consideration of cyclothems is as markers of nonmarine environments. The name Asterophyllites assemblage is taken from the comparatively rich, well-preserved land flora which has been collected from the Stranger Formation (Tonganoxie Member) at localities 3.5 miles southeast of Lawrence (Haverkampf farm) and 5 miles northeast of Baldwin (Holcomb farm). Plants from these beds, first described by Sellards (1908a), recently have been reviewed by Cridland, Morris, and Baxter (1963) for the purpose of confirming identifications and making comparisons with Carboniferous plant-bearing beds elsewhere in North America and Europe. The symbol "ST" (from Stranger-Tonganoxie) is adopted for use on diagrams. Associated with Asterophyllites are various species of Alethopteris, Aphlebia, Annularia, Asterotheca, Cordaites, Cordianthus, Cyclopteris, Mariopteris, Neuropteris, Odontopteris, Pecopteris, Ptychocarpus, Sphenophyllum, Sphenopteris, and Spiropteris (Suppl. Fig. 1, 1-7).
In aggregate a rather rich flora of land plants has been made known from the Kansas region, and counting Cherokee cyclothems, it is noteworthy that described plants come from not less than 40 horizons in Pennsylvanian and Permian deposits, with widely distributed marine strata intercalated between nearly all successive land-plant beds. The plant fossils, including many well-preserved delicate fronds of ferns and similar remains, indicate deposition in lakes, lagoons, swamps, and waters of sluggish streams.
Animals associated with the type Asterophyllites assemblage near Lawrence comprise a cockroach fauna described by Sellards (1908b) which includes 19 species referred to eight genera. Specimens have been collected subsequent to the work of Sellards but no systematic study of them has been made.
A pictorial reconstruction of the so-called Asterophyllites assemblage, representing an example belonging near the top of the Lawrence Shale at Lawrence, Kansas, is given in Figure 4. Paleobiotopes immediately following this one in the same area are illustrated in Figures 5 and 6.
Figure 4--Restoration of Stranger-type (Asterophyllites) ecosystem associated with coal-forming conditions near end of Lawrence Shale deposition, view looking west from site of Douglas County courthouse in Lawrence, Kansas (Moore).

Figure 5--Restoration of paleobiotope introduced by marine flooding of coal swamp illustrated in Figure 4, with same location and direction of view (Moore).

Figure 6--Seascape at Lawrence, Kansas, in early Oread time, defective as a paleoecologic illustration in that depth of water, nature of salinity, and type of bottom sediment cannot be shown (J. W. Koenig).

Skeletal remains and tracks of tetrapods are too uncommon in Pennsylvanian and Permian strata of Kansas to warrant more than brief notice as paleoecological indicators. At most they are highly sporadic, have only local significance, and few fossils of this sort are well enough recorded to allow determination of their relation to cyclothems. An exception is the comparatively rich find of land vertebrates in shaly upper Stanton beds near Garnett, in Anderson County, and this is discussed in a later part of the paper under the designation "Rock Lake-type (Garnettius) assemblage."
Another consists of some well-marked tracks in upper Howard layers near Osage City south of Topeka, identified (Schoewe, 1956) as belonging to the Utopia Limestone Member; these have been described as belonging to amphibians (Limnopus vagus, L. littoralis, Nanopus caudatus) and a small reptile (Dromopus agilis) with tendency toward bipedal locomotion (Marsh, 1894; Baird, 1952). The presence of raindrop impressions associated with some of the tracks is of interest.
Tracks probably made by amphibians (Crucipes parvus, Collettosaurus missouriensis) in platy sandy shale of the Kansas City Group (precise horizon and locality unknown) have been reported by Branson and Mehl (1932).
A single three-toed birdlike footprint with well-defined, relatively long heel, strikingly similar to Triassic dinosaur tracks of the Connecticut Valley, was found many years ago near the Pennsylvanian-Permian boundary at a locality 2 miles northwest of Eureka, in Greenwood County. The print, approximately 6 inches long and nearly 5 inches wide, was described and figured by Moodie (1913), who gave it no name but guessed that the existence of a Kansas Permian dinosaur is within the realm of possibility.
Skeletal remains of amphibians identified as Cricotus and Eryops have been found in Lower Permian rocks of Cowley County, near the Oklahoma boundary, and bones of the pelycosaur reptile Dimetrodon in approximately equivalent beds just south of the state line (Lane, 1946). Also, Lower Permian strata of Washington County, adjoining Nebraska, have yielded remains of another amphibian named Erpetosuchus (Lane, 1946), and a new genus of trimerorhachoid amphibians named Acroplous has been described from upper Council Grove strata (Speiser Shale) in Riley County, Kansas (Hotton, 1959).
Finally, mention may be made of presumed tracks of a giant amphibian (Martin, 1922) in the Tonganoxie Sandstone (lower Virgilian) a few miles southeast of Lawrence. The animal inferred from the tracks was named Oncychopus gigas (= Wakarusopus gigas Moodie, 1931), but the marks, real enough in themselves, now are judged to be calamite stump holes and not vertebrate tracks at all (Baird, 1963).
A paleobiotope of unusual interest is represented by closely studied deposits found in the upper part of the Stanton Formation of late Missourian age (early Late Pennsylvanian) approximately six miles northwest of Garnett, in sec. 32, T. 19 S., R. 19 E., Anderson County, Kansas. It comprises part of a cyclothem which lies disconformably on well-bedded marine limestone (Stoner Member of the Stanton) and reflects an environment interpreted as that of a somewhat brackish-water lagoon, neither typically marine nor terrestrial, for organic remains collected from its layers are a mixture of invertebrates normally confined to saline seaways and well-preserved land plants, air-breathing arthropods, riveror lagoon-dwelling coelacanth fishes, amphibians, and reptiles of cursorial and possibly semiarboreal types. This ecosystem is here termed the Garnettius assemblage, named from a distinctive scorpion originally described as Mazonia, hungerfordi Elias (in Moore, Elias, and Newell, 1936, p. 12; also, Elias, 1937b). The stratigraphic position of the unique assemblage is marked on Figure 7 by the symbol "RL" (denoting the Rock Lake Shale Member of the Stanton Limestone).
Figure 7--Correlated sections of Lansing Group (upper Missourian) in northeastern Kansas showing stratigraphic occurrence of Rock Lake (Garnettius ecosystem ("RL"), also example of Heebner-type (Listracanthus) paleobiotope (Moore, sections measured by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
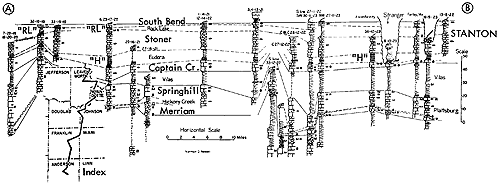
The Garnettius assemblage is comparable to the famous Burgess fauna from the Middle Cambrian of British Columbia in furnishing a glimpse of associated Paleozoic organisms unmatched by any other like community. Yet our knowledge of it obviously depends on the fortunate circumstances of exposure of a part of the deposits and the "accident" of discovery by N. D. Newell in the course of field work for the Kansas Geological Survey in 1931. If the strata and their contained fossils had been concealed by cover or if the outcrops had been unnoticed and unexplored, the Garnettius assemblage now would be as much unknown as scores of possible similar occurrences in older, age-equivalent, and younger Pennsylvanian and Permian deposits of the Kansas region.
Fossils of the South Bend Limestone Member of the Stanton at the locality northwest of Garnett are not included in the Garnettius assemblage, although they belong to the same cyclothem as that which here begins with the shaly strata (Rock Lake) containing the Garnettius community. The South Bend fauna is typically marine; it includes the fusulinid Triticites in addition to brachiopods, bryozoans, bivalves, gastropods, corals, and crinoid and echinoid remains.
Among marine invertebrates of the Garnettius assemblage are: (1) corals (?Lophophyllidium), (2) columnals of crinoids, (3) brachiopods (Lingula, Composita), (4) bryozoans (Fenestrellina, Polypora, Rhombopora), and (5) bivalves (Myalina, Yoldia, Sedgwickia) . Evidence found in ecosystems of other cyclothems suggests that all of these invertebrates except the corals are euryhaline forms, tolerant of considerable variation in salinity of waters around them, and they are interpreted to have been at home in a near-shore environment. They could have inhabited a shallow bay or lagoon having more or less restricted connection with an adjacent open shallow sea. Lingula (Suppl. Fig. 1, 8) is a burrowing inarticulate brachiopod which can survive in moderately brackish or even fresh water for brief periods of time by retreating to the deepest part of its burrow and tightly closing its shell. The genus may occur in the dark muds of tidal flats, whereas no brachiopods, including Lingula, are adapted for life in distinctly low-salinity environments.
Remains of land plants, which are a relatively abundant constituent of the Garnettius assemblage, surely were transported to the places where they became buried, as demonstrated by their association with marine organisms. A majority of the fossils are exceptionally well preserved, however. Genera (some illustrated in Suppl. Fig. 1) which have been identified are as follows, number of recognized species being indicated by accompanying numerals: Alethopteris (2), Annularia (1), Callipteridium (1), Cordaites (2), Desmopteris (2), Dichophyllum (2), ?Dicranophyllum (2), ?Lecrosia (1), ?Lepidophyllum (1), Neuropteris (2), Odontoperis (2), Palaeophycus (2), Pecopteris (1), Pteridospermostrobus (1), Samaropsis (4), Sphenopteris (3), Taeniopteris (4), ?Ulmania (1), Voltzia (1), and Walchia (4). Although hydrophytic and mesophytic types of ferns, pteridosperms, horsetails, lycopods, and sphenophylls are found in this flora, it is dominated by gymnosperms, especially the coniferlike Walchia, which denote a dry environment classifiable as xerophytic. Well-preserved specimens of coniferophyte wood from the Garnett area also have been described (Baxter and Hartman, 1954). This character and the presence of such plants as Taeniopteris, Dichophyllum, and Dicranophyllum so much resemble Permian floral assemblages that David White, a widely experienced paleobotanist of the U. S. Geological Survey, failed to see how the deposits near Garnett could be older than Permian (Moore, Elias, and Newell, 1936, p. 2, 12). In fact, they belong approximately 1,100 feet below the Pennsylvanian-Permian boundary as drawn in Kansas.
The terrestrial arthropods of the Garnettius assemblage include one genus of chelicerates, the scorpion Garnettius (Petrunkevitch, 1953, p. 34), and four genera of insects--the cockroaches Phyloblatta and Mylacris, and megasecopterans named Euchoroptera and Parabrodia (Carpenter, 1934, 1940). The megasecopterans are medium-sized to large insects with four subequal outspread wings, small head, and relatively slender, long abdomen. The cockroaches were chiefly cursorial on land but all of the Garnett insects could fly; they may have been air-borne to places where they dropped into waters of the Garnettius biotope or, more likely, their remains were transported by sluggish streams to the shallow bay or lagoon in bottom sediment of which they became buried.
Special interest relates to finds of vertebrates in the Garnettius-bearing deposits. These include coelacanth fishes (Hibbard, 1934), an amphibian named Hesperoherpeton garnettense (Peabody, 1958) which represents a new order named Plesiopoda (Eaton and Stewart, 1960) and four kinds of reptiles, the most important of which, represented by nearly complete skeletons and parts of skeletons, is a long-toed lizardlike form called Petrolacosaurus kansensis (Fig. 8), approximately 24 inches in length at maturity (Lane, 1945; Peabody, 1952). The other reptiles are all pelycosaurs, characterized by elongation of their vertebral neural spines to make a tall fin running along the middle of the back; one has been described as Edaphosarus ecordi (Peabody, 1957) and the others are an ophiacodont allied to Clepsydrops and a primitive sphenacodont. The fishes and amphibian probably lived in a stream or streams that emptied into the Garnett lagoonlike water body but the reptiles were animals of dry land. Peabody's (1952, p. 38) interpretation of the environment is worthy of quotation; with some omissions this is as follows.
Terrestrial and fluviatile organisms were rafted by a slow-running river into a marginal embayment or lagoon protected by a barrier sand bar. Calcareous mud, mainly transported by the river, was deposited in thin, possibly cyclic layers on the eroded surface of a marine limestone. The lagoon was deep enough so that waves did not disturb the bottom and the barrier bar was complete enough to exclude strong marine currents. Under these conditions, thin-bedded mud devoid of ripple marks or scour marks accumulated and formed what is now approximately 10 feet of fossiliferous shale. Deposition of mud served to impoverish the marine invertebrate fauna and exclude scavenging forms, while simultaneously preserving the remains of indigenous marine invertebrates and rafted terrestrial organisms. Rafted material became waterlogged, probably during transport, and some maceration occurred, for quantities of detached needles of conifers literally blacken some of the bedding planes. Corpses of reptiles came to rest on the mud bottom and became partly embedded. Before complete burial, differential decay weakened exposed parts. For example, the immature delicate skull rested thus dorsal side up. Most of the roof of the delicate skull then became detached and drifted away probably with the strengthened current which deposited the next layer of mud. Parts of the roof remained, because they were locked in place by the deeply embedded lower jaws and palate. . . . It is reasonable to assume that the plants, arthropods and reptiles may have been transported from one and the same terrestrial environment and at the same time by relatively quiescent but perhaps flooding waters of a Pennsylvanian river. . . . The river flowed through a relatively dry landscape dominated by conifers. Completeness of the reptilian skeletons and isolated limbs, the presence of delicate winged insects, and of fruits of conifer and pteridosperm suggest a common place of origin and one that was not far away. . . . Association of Petrolacosaurus with conifers raises the possibility of arboreal habits; elongate digits and lightened structure of the bones are suggestive of climbing ability. . . . Petrolacosaurus was an agile terrestrial reptile.
Figure 8--Restoration of part of the Garnettius ecosystem showing especially lizardlike reptile, Petrolacosaurus, approximately two feet long, and in central foreground scorpion, Garnettius. (Moore, after Peabody, 1952, painting by Victor Hogg).

Attention given to the Garnettius assemblage in this paper makes a very adequate excuse, if one is needed, for notice of another nonmarine assemblage which has been described from the famous insect-bearing deposits in the Wellington Shale near Elmo, in southern Dickinson County, Kansas, even though these are not defined as part of a recognized cyclothem. Obviously, they signify a paleobiotope which is closely comparable to that of upper Stanton beds near Garnett, though doubtless located in a prevailingly dryer climatic setting. Here, approximately 1,000 feet above the Pennsylvanian-Permian boundary, again are found marine invertebrates associated with a varied land-plant flora and a host of terrestrial arthropods, mostly insects but including aquatic chelicerates related distantly to the modern kingcrabs (Limulus). Richness of the insect fauna is indicated by published records of 7,000 or more specimens collected from the Elmo locality.
The ecologic community (ecosystem) of the Wellington in the Elmo area is designated as the Sellardsia assemblage, despite the fact that Sellardsia is a nominal genus of relatively unimportant insects and that it is considered (Carpenter, 1935, p. 129) to be a junior synonym of Lecorium Sellards (1909, p. 167). Choice of Sellardsia in this connection is dictated by the wish to honor the paleontologist who in 1902 discovered the fossil plants and insects near Elmo. After completing undergraduate and early graduate studies at The University of Kansas, Sellards was then working on his doctorate at Yale University. During the summers of 1902 and 1903 he collected some 2,000 specimens (Sellards, 1903) and subsequently described many of them (Sellards, 1906, 1907, 1909). Later, extensive collecting by Dunbar (1924, p. 173), and Carpenter (1930, p. 70; 1933, p. 411; 1939, p. 29) furnished materials for a long series of papers by Tillyard (1923-1937) and by Carpenter (1930-1950).
The invertebrates of the Sellardsia assemblage come from an evenly bedded impure limestone, 4 or 5 feet thick, mostly near the base. Locally abundant in the lowermost bed of the limestone are shells of a small bivalve identified as belonging to the marine genus Myalina, but unusually thin-shelled and interpreted, partly on this account, as abnormal, being migrants into a brackish-water habitat (Dunbar, 1924, p. 201). Nearly black shale just below the limestone contains silicified stumps and some logs of lycopods, probably Cordaites, and beneath this layer is interbedded gray shale and thin limestone. A comparatively large flora of well-preserved ferns, lycopods, horsetails, and conifers has been described from the shale. The calcareous beds have yielded minute pelecypods, conchostracans, and two kinds of merostome chelicerates. Remains of one small merostome have been described under the name Eurypterus (Anthraconectes) sellardsi (Dunbar, 1924, p. 199); these occur in association with specimens of a small limulid (Paleolimulus avitus Dunbar, 1923, Suppl. Fig. 1, 9). The plants and animals found below the insect.bearing limestone are not included in the Sellardsia assemblage.
In view of the exceptional paleontological importance of the Lower Permian insect fauna described from the Elmo, Kansas, area I have undertaken to survey its presently determined taxonomic content in terms of genera, where possible recording junior synonyms in square brackets and number of assigned species, if greater than one, by numerals enclosed in parentheses. I am grateful to Professor Frank M. Carpenter, of Harvard University, for corrections and other help in preparing the list, which is organized by orders recognized by him in the Treatise on Invertebrate Paleontology. Classification of Permian fossil insects now adopted by Carpenter differs materially from that earlier published by Brues, Melander, and Carpenter (1954) in their comprehensive survey of all living and fossil insect orders.
| Lower Permian Insect Fauna from Elmo, Kansas | |
|---|---|
| Palaeodictyoptera. Generalized four-winged insects with main veins of wings usually independent, beak commonly adapted for feeding on liquid foods, legs and abdominal segments unspecialized. U. Carb.-Perm. | |
| Calvertiella Tillyard, 1925 | |
| Permoneura Carpenter, 1931 | |
| Dunbaria Tillyard, 1924 | |
| Kansasia Tillyard, 1937 | |
| Megasecoptera. Insects with four subequal wings having all main longitudinal veins in alternating ridges and furrows, mostly with well-developed cross veins rather than irregular network (archedictyon). U. Carb.-Perm. | |
| Protohymen Tillyard, 1924 (3) [= Pseudohymen Martynov, 1932; Pseudohymenopsis Zalessky, 1956 ] | |
| Permohymen Tillyard, 1924 | |
| Diaphanopterodea. Like Megasecoptera but with wings held backward along abdomen when at rest. U. Carb.-U. Perm. | |
| Elmoa Tillyard, 1937 | |
| Martynovia Tillyard, 1932 [= Martynoviella Tillyard, 1932] | |
| Ephemeroptera. Wings with all main veins, very delicate; head with large compound eyes; abdomen slender (mayflies). U. Carb.-Rec. | |
| Protereisma Sellards, 1907 (5) [= Protechma, Prodromus, Bantiska, Recter, Pinctodia, Esca Sellards, 1907; Mecus Sellards, 1909; Loxophlebia Martynov, 1928] | |
| Misthodotes Sellards, 1909 (3) | |
| Eudoter Tillyard, 1932 | |
| Protodonata. Medium-sized to very large insects (some with 30-inch wing spread), with globose head and long slender abdomen (early dragonflies). U. Carb.-Perm. | |
| Meganeuropsis Carpenter, 1939 | |
| Megatypus Tillyard, 1925 (2) | |
| Tupus Sellards, 1906 (3) [= Meganeurula Handlirsch, 1906; Gilsonia Meunier, 1908; Typus Sellards, 1909; Meganeurina Handlirseh, 1919; Arctotypus Martynov, 1932] | |
| Oligotypus Carpenter, 1931 | |
| Odonata. Predaceous insects with four subequal wings, large compound eyes, and long slender abdomen (dragonflies). L. Perm.-Rec. | |
| Ditaxineura TiIIyard, 1926 | |
| Kennedya TiIIyard, 1925 | |
| Progoneura Carpenter, 1931 | |
| Camptotaxineura TiIIyard, 1937 | |
| Protorthoptera. Four-winged, much as in some Palaeodictyoptera but lacking complete alternations of ridges and furrows along main veins, small head with prominent antennae, legs mostly adapted for running. U. Carb.-Trias. | |
| Pursa Sellards, 1909 | |
| Sindon Sellards, 1909 | |
| Lemmatophora Sellards, 1909 | |
| Artinska Sellards, 1909 (3) [= Estadia, Lectrum, Orta Sellards, 1909] | |
| Lecorium Sellards, 1909 [= Stemma Sellards, 1909; Sellardsia Tillyard, 1928; Paralecorium, Metalecorium Handlirsch, 1937] | |
| Lisca Sellards, 1909 | |
| Paraprisca Handlirsch, 1919 (2) [= Prisca Sellards, 1909 (non Fritsch, 1899)] | |
| Liomopterum Sellards, 1909 [= Horates Sellards, 1909] | |
| Semopterum Carpenter, 1950 | |
| Tapopterum Carpenter, 1950 | |
| Phenopterum Carpenter, 1950 | |
| Probnis Sellards, 1909 [= Espira, Stoichus, Stinus Sellards, 1909] | |
| Chelopterum Carpenter, 1950 | |
| Demopterum Carpenter, 1950 | |
| Orthoptera. Includes suborder Blattaria, cockroaches. U. Carb.-Rec. | |
| Etoblattina Scudder, 1879 (3) | |
| Puknoblattina Sellards, 1908 (2) [= Pycnoblattina TiIIyard, 1937] | |
| Permoblattina Tillyard, 1937 | |
| Promartynovia TiIIyard, 1937 | |
| Caloneurodea. Like Protorthoptera except for close similarity of fore and hind wings in form, venation, and texture. U. Carb.-Perm. | |
| Anomalogramma Carpenter, 1943 | |
| Paleuthygramma Martynov, 1930 | |
| Apsidoneura Carpenter, 1943 | |
| Permobiella TiIIyard, 1937 | |
| Pleisiogramma Carpenter, 1943 (2) | |
| Protelytroptera. Small insects related to cockroaches, eyes conspicuous and antennae prominent, fore wings developed as hard cover structures (elytra). L. Perm. | |
| Elytroneura Carpenter, 1933 | |
| Archelytron Carpenter, 1933 | |
| Protelytron TiIIyard, 1931 (3) | |
| Permelytropsis Carpenter, 1933 | |
| Megelytron TiIIyard, 1931 | |
| Permelytron TiIIyard, 1931 | |
| Blattelytron TiIIyard, 1931 | |
| Parablattelytron TiIIyard, 1931 (3) [= Acosmelytron TiIIyard, 1931] | |
| Protelytropsis TiIIyard, 1931 | |
| Corrodentia. Mostly diminutive short.bodied insects with large head and prominent eyes; usually winged. Perm.-Rec. | |
| Dichentomum TiIIyard, 1926 (2) [= Psocidium, Chaetopsocidium, Metapsocidium, Pentapsocidium, Permentomum TiIIyard, 1926] | |
| Progonopsocus TiIIyard, 1926 (2) [= Ancylopsocus Tillyard, 1926] | |
| Cyphoneura Carpenter, 1932 | |
| Orthopsocus Carpenter, 1932 | |
| Lithopsocidium Carpenter, 1933 | |
| Cyphoneurodes Becker-Migdisova, 1953 | |
| Miomoptera. Small insects with similar fore and hind wings, latter lacking anal fan. Penn.-Perm. | |
| Delopterum Sellards, 1909 (3) | |
| Permembia TiIIyard, 1928 | |
| Nugonioneura TiIIyard, 1928 | |
| Hemiptera. Diverse assemblage of mostly small insects with wings usually sloping over sides of body, mouth parts modified for piercing and sucking. Perm.-Rec. | |
| Archescytina TiIIyard, 1926 | |
| Permoscytina Tillyard, 1926 | |
| Permopsylla TiIIyard, 1926 | |
| Paleoscytina Carpenter, 1931 | |
| Lithoscytina Carpenter, 1933 | |
| Neuroptera. Soft. bodied insects with relatively large wings, combed antennae, and similar legs. Perm.-Rec. | |
| Permoraphidia TiIIyard, 1932 (2) | |
| Glosselytrodea. Small insects with fore and hind wings almost alike, veins very straight, cross veins numerous, forming large cells, precostal area prominent, hairs on main veins. Perm. | |
| Permoberotha TiIIyard, 1932 [= Dictyobiella TiIIyard, 1937] | |
| Mecoptera. Small to medium-sized slender insects, nearly all with downward prolongation of head in form of beak (scorpion flies). Perm.-Rec. | |
| Permopanorpa TiIIyard, 1926 (3) | |
| Platychorista TiIIyard, 1926 [= Protomerope TiIIyard, 1926] | |
| Protochorista TiIIyard, 1926 | |
| Protopanorpa TiIIyard, 1926 | |
| Anormochorista TiIIyard, 1926 | |
| Lithopanorpa Carpenter, 1930 | |
| Agetopanorpa Carpenter, 1930 | |
| Choristosialis TiIIyard, 1932 | |
This is a surprisingly long list. Not counting synonyms, it contains 79 genera distributed among 16 insect orders, emphasizing not only the remarkable variety of the assemblage found in early Permian deposits of a single area but also the almost incredible richness of well-preserved fossils, assiduously collected by painstaking field workers and thoroughly studied by competent specialists. Even so, knowledge surely is far from complete and further paleoecological studies are warranted. Dunbar's (1924, p. 197) paleoecological inferences are summarized in the following description.
As we have seen from a study of the lithology of the insect beds and the organic remains associated with them, the stump bed of the local section indicates here a temporary emergence when the vicinity of Insect Hill was added to the lowland bordering the Kansas sea on the northeast. The swamp conditions show that it was so low as to be poorly drained and also that the rainfall was now sufficient at least to maintain a high ground-water table in such favorable low places. The swamp, after a few hundred years, was displaced by open water, at first probably a nearly fresh lagoon but slightly connected to the nearby inland sea. Soon it became entirely a fresh-water lake or lagoon, covering the site of Insect Hill, though the abundant but fragmentary leaves of land plants give evidence that timbered lands were still adjacent. Probably the swamp had widened as the deeper portions were submerged. The bordering forests furnished a home for the prolific insect fauna, which, probably driven by occasional storms, fell into the open waters in great numbers. Since there were no fish or other animals in the lagoon to devour them, the insects floated and drifted about, sometimes settling singly, but at other times drifting together in great numbers, as flotsam is wont to do, into protected places in the lee of some obstruction to the winds. Apparently the level of the water fluctuated frequently, and thus at intervals these shallow marginal places were left dry for a short time, when the insects were stranded on the soft limy mud bottom. After a short period of drying, the insects did not float when the water returned, but stuck to the bottom where they were soon buried beneath the next film of limy deposit. The distribution of the fossil insects upon the bedding planes thus finds an easy explanation. The rich "pockets" were protected places. . . . The intimate association of broken and badly preserved fragments, along with others more sharply preserved and complete, may be attributed to the fact that the former had drifted longer and been broken and partly decayed before burial, whereas the finest specimens had fallen but a short time before burial, or possibly, in some cases had alighted or been driven directly upon the exposed mud bottom.
Although the immediate home of these insects was . . . a moist locality, nevertheless they can hardly have escaped the influence of the long-enduring dryness of the larger environment. Such delicate organisms could only be recorded in places where moisture helped to conserve them, but the insect fauna may not have been restricted in its distribution to such surroundings. Granted, however, that they inhabited only the more humid portions of the semiarid landscapes, the preceding generations . . . must have experienced, time and again, the relentless encroachment of droughts upon their shrinking retreats as the climate oscillated from less to greater aridity.
The Sellardsia assemblage of the Elmo, Kansas, area is still the most remarkable Permian ecosystem of the sort yet discovered in North America but it is likely that comparable ones lie buried in nearby or distant parts of the state and that additional ones have been destroyed by erosion. In recent years rich insect-bearing Lower Permian beds have been found in southern Kansas and northern Oklahoma (Carpenter, 1947; Tasch and Zimmerman, 1959).
The several sorts of marine ecosystems selected for consideration out of the many, partly intergrading types observed in Kansas Pennsylvanian and Permian deposits are arranged in a sequence that corresponds approximately to the order of occurrence in the ideal cyclothem, that is, assemblages such as marked by inarticulate brachiopods (Lingula, Orbiculoidea) and little else or by associations of presumably nearest shore mollusks (mostly clams such as Myalina and pectinoid genera) either occurring alone or accompanied by low-salinity-tolerant brachiopods (especially Derbyia) and ramose bryozoans. Chonetoid brachiopods in great abundance may characterize the initial marine environment in a cyclothem. Algae of all kinds tend to be lacking in transgressive marine stages, but make appearance in company with normal marine assemblages of invertebrates in middle stages and are characteristically abundant in regressive parts of the marine cycles. Varied populations containing representatives of several phyla appear in middle deposits of cyclothems, with or without the presence of fusulinids. An abundance of fusulinids which seem to crowd out other organisms characterizes many limestones that mark culmination of sea advances. Accordingly, examples of these are taken up in a sequence that puts nearshore transgressive paleobiotopes first, then those found in middle parts of cyclothems, and finally those which typically mark regressive phases of the shallow-sea oscillations.
Some marine paleobiotopes are not classifiable according to this plan. For example, the environment (conceivably more than a single kind of environment) represented by thin but very extensive black platy shales was introduced in parts of the stratigraphic record immediately following coal-swamp conditions but elsewhere, especially in Missourian and Virgilian parts of the column, the black shale next overlies typical marine limestone containing invertebrates adapted to offshore environments. Such relations do not prove that these black shales belong to regressive stages of the cycle to which the underlying limestone belongs because the knife-sharp boundary between shale and limestone actually may represent a disconformity (paraconformity), the shale being in fact the initial deposit of a new cycle. Therefore, discussion of the black platy shale paleobiotopes is postponed to a last part of this paper.
Sparse to common, or even locally abundant inarticulate brachiopods characterize a marine assemblage which here is called Red Eagle type because it is found to be exceptionally persistent within the Red Eagle cyclothem. Orbiculoidea and Lingula generally are present, in some places associated with many specimens of Crurithyris (Suppl. Fig. 1, 8,10,11). The assemblage chosen as type occurs in the Bennett Shale Member of the Red Eagle Limestone, which is a formation readily traced from Oklahoma to Nebraska in the lower part of the Council Grove Group (Lower Permian) and the black to very dark-gray shale containing this assemblage occurs at the very base of the member. Outcrops at the type locality of the Bennett, 0.5 mile southeast of Bennet (sic), Nebraska, are designated as containing a representative sample of this community. Other assemblages of Bennett type are well developed at many places in the Grenola Limestone of northern Kansas and southern Nebraska, in shale of the Ozawkie Limestone (Deer Creek) of central Kansas, and elsewhere. Unlike the hard platy black shales of Heebner type, the shales containing the Orbiculoidea-Lingula assemblage are soft; some appear to be rather local in occurrence, pinching out laterally, but McCrone (1963) traced the unit at the base of the Bennett nearly 300 miles and reported the presence of Orbiculoidea at virtually all observed exposures (Fig. 9). He recorded also the finding of several genera of conodonts and ostracodes and locally small teeth of fishes identified as Palaeoniscus, Idiacanthus, and Distacodus. Macerated plant debris was found to be common but spores rare. McCrone interpreted the Red Eagle-type paleobiotope as representing a poorly oxygenated shallow sea bottom which he estimated to be less than 10 feet below sea level (p. 64). He thought that waters in which the Red Eagle assemblage lived were not freely connected with the open sea (McCrone, 1963, p. 65).
Figure 9--Generalized sections of lowermost Permian deposits in four parts of Kansas region, showing stratigraphic occurrences of some ecosystems ("B," Beil-type; "RE," Red eagle-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
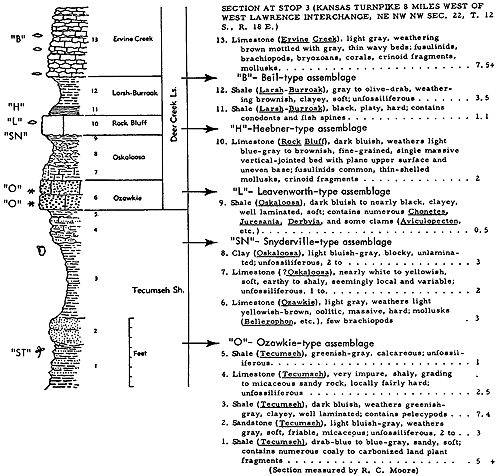
The environment of the Red Eagle-type Orbiculoidea-Lingula assemblage mentioned as occurring in beds classed as part of the Ozawkie Member of the Deer Creek limestone can hardly be interpreted as located in a marginal belt of a transgressing sea, for the abundant inarticulates (unassociated with other brachiopods) occur in shale next above a fusulinid-rich middle Ozawkie Limestone and below unfossiliferous earthy brown upper-most Ozawkie Limestone. These beds are excellently exposed 5 miles north of Lyndon, Osage County, Kansas (Fig. 10).
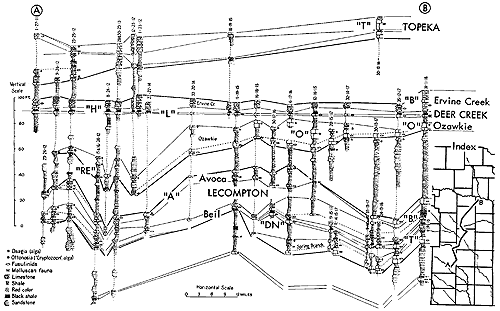
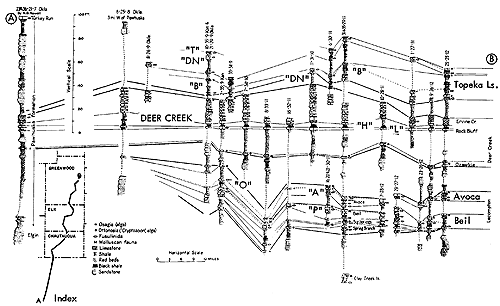
Figure 10--Correlated sections of Shawnee Group (Virgilian) units in eastern Kansas, showing stratigraphic occurrence of some specified ecosystems ("A," Avoca-type; "B," Beil-type; "DN," Doniphan-type; "H," Heebner-type; "L," Leavenworth-type; "O," Ozawkie-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
Because the black Orbiculoidea-rich basal shale of the Bennett overlies the fusulinid-bearing Glenrock Limestone and is followed above by osagite of the Howe Limestone, Elias (1937a, p. 407) interpreted the Bennett as belonging to the regressive part of the Red Eagle cyclothem. Conversely, McCrone (1963, p. 50) classed the black shale as the initial deposit of a secondary transgression within the Red Eagle cycle, because the Orbiculoidea-Lingula beds lie below gray shale of the Bennett and limestone containing a varied articulate-brachiopod and bryozoan fauna, rightly interpreted (in my judgment) as representing a farther offshore, somewhat deeper water environment than that of the black shale deposition. If this is correct, the sharp boundary between the black shale and subjacent Glenrock Limestone (with its commonly abundant contained fusulinids) marks an extremely abrupt marine retreat, and missing regressive stages that should succeed the Glenrock transgressive culmination denote what amounts to a disconforrnity (paraconformity). The question of transgressive or regressive classification of the black shale is relatively unimportant as compared with determination of the nature of the paleobiotope recorded by Red Eagle-type assemblages.
Shells of Derbyia (Suppl. Fig. 1, 12) in relatively large numbers and a few other euryhaline brachiopods, mingled in some deposits with similarly salinity-tolerant clams, comprise records in various Pennsylvanian and Permian cyclothems of the Kansas region of a ecosystem designated as the Speiser-type (Derbyia) assemblage. Excellent examples near the top of the Speiser Shale in the vicinity of Dexter (southeastern Cowley County) and elsewhere along outcrops of shale and shaly limestone next below the Wreford Limestone which forms a prominent escarpment across Kansas, suggest adoption of Speiser-type for a rock-unit-derived name of this distinctive assemblage. Examples invariably belong to initial marine phases of cyclothems, being found above nonmarine deposits which may contain land plants and in many places may be represented by a coal bed, and they are normally overlain by limestone bearing a variety of typical marine invertebratescorals, bryozoans, several genera of brachiopods, crinoid remains, gastropods, pelecypods, and possibly fusulinids-which are interpreted as belonging to an offshore population (Fig. 11, 12). Derbyia may be so abundant that shells are packed together almost as a coquina. Associated with it, especially in some Permian examples, are scattered individuals belonging to /uresania and Composita. Species of Myalina or Septimyalina and Aviculopecten are common constituents of the assemblage both in Pennsylvanian horizons and in the Permian. Rather unexpected constituents of some Derbyia assemblages (e. g., below the Haskell Limestone locally in Douglas County, and upper Doniphan Shale in Osage County) are well-preserved crinoid cups and common stem fragments associated with numerous ramose bryozoans.
Figure 11--Generalized sections of Lower Permian rocks extending upward from those shown in Figure 9, with examples of some ecosystems ("F," Florena-type; "S," Speiser-type; "T," Tarkio-type; "TM," Threemile-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
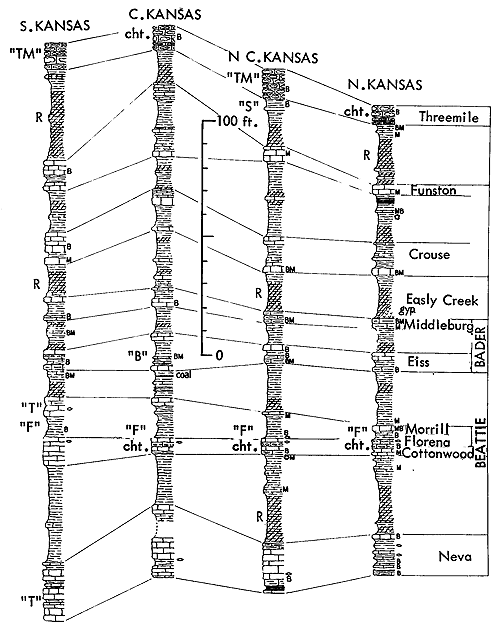
Figure 12--Generalized sections of Lower Permian rocks extending upward from those shown in Figure 11, with examples of some ecosystems ("T," Tarkio-type; "TM," Threemile-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
In southern Kansas especially, fossils of Derbyia assemblages in the upper Speiser Shale are silicified. In addition to fine specimens collected from weathered outcrops, numerous shells which show perfectly preserved internal features, as well as exterior surfaces, have been obtained by etching blocks of somewhat shaly rock in dilute hydrochloric acid. Hattin (1957) has described the fossil associations of this part of the Lower Permian section not only in southern Kansas but as observed at many outcrops in central and northern parts of the state. He distinguished lowermost fossiliferous beds dominated by mollusks (chiefly pectinoid pelecypods in most places, especially toward the north), and slightly higher beds dominated by Derbyia. Both groups are here included in the so-called Derbyia assemblage, which in the Speiser exposures contains the following forms (Hattin, 1957).
| Organisms of the Derbyia Assemblage in the Speiser Shale just below the Wreford Limestone in Kansas (Hattin, 1957) | |
|---|---|
| Algae (algal-foraminiferal intergrowths, Osagia, not common) | |
| Invertebrates (generally well-preserved, locally abundant) | |
| Foraminifers: Ammodiscus, Ammovertella, Climacammina, juvenile fusulinids (uncommon), Globivalvulina, Geinitzina, Hyperammina, Tetrataxis | |
| Corals: Aulopora, Stereostylus | |
| Bryozoans: cyclostomes (encrusting), Fenestrellina, Penniretepora, Polypora, Rhabdomeson, Rhombopora, Septopora, Streblotrypa, trepostomes (encrusting), Thamniscus, ?Batostomella, ?Leioclema | |
| Brachiopods: Chonetes [= Neochonetes], Composita, Derbyia, Dictyoclostus [Reticulatia], Enteletes, Juresania, Orbiculoidea, Petrocrania | |
| Pelecypods: Allorisma, Aviculopecten, Aviculopinna, Schizodus, Septimyalina | |
| Gastropods: Bellerophon, Euomphalus, Euphemites, ?Cymatospira | |
| Arthropods: ostracodes (Amphissites, Bairdia, Bythocypris, Cavellina, Cornigella, Ellipsella, Healdia, Hollinella, Kellettina, Kirkbya, Knightina, Knoxina, Macrocypris, Monoceratina, Paraparchites, Roundyella, Silenites, ?Haworthina), trilobite, Ditomopyge | |
| Echinoderms: crinoids (Delocrinus, stem fragments, plates), echinoids (spines, plates), holothuroids (spicules, hooks, wheels) | |
| Conodonts: unidentified genera | |
| Vertebrates (fragmentary remains of fishes, teeth, plates, bones) | |
Hattin (1957, p. 113) interpreted the environment of this so-called Derbyia assemblage as belonging to an offshore zone of nearly normal salinity in which not very strong turbulence resulted from wave activity. Conditions generally were favorable for many forms of life. Disseminated calcium carbonate he judged to have been derived from shell disintegration.
In several Pennsylvanian cyclothems deposits identified as representing initial parts of marine sequences are characterized by specially abundant ramose bryozoans, generally including Rhombopora (Suppl. Fig. 1, 15) and Batostomella. This ecosystem, which rarely contains a few other invertebrates, is termed the Rhombopora assemblage (Fig. 10) and from excellent samples in the Doniphan Shale Member of the Lecompton Limestone called Doniphan type. In one exposure of this shale west of Melvern, in southern Osage County, these bryozoans weather out in such profusion that the ground is nearly concealed by a seeming cover of straw. The stems of "straw" are a trifle short (averaging about 2 inches) and many of them show bifurcation toward one end, but their diameter, straightness, and evenly cylindrical form closely approximate sections of wheat straw; moreover, all specimens at this locality have the color of wheat straw.
The lowest marine shaly deposits of some Kansas Pennsylvanian cyclothems contain extraordinarily abundant chonetoid brachiopods which are accompanied by few other invertebrates. An assemblage of this sort is well shown in the uppermost part of the Snyderville Shale Member of the Oread Limestone as seen at many outcrops in central and northeastern Kansas and southeastern Nebraska. Nearly all of the shells belong to Neochonetes granulifer (Suppl. Fig. 2, 6) and therefore the ecosystem is named the Neochonetes assemblage of Snyderville type. Its representation in exposures at several places within a mile or two of Toronto, in southwestern Woodson County, is chosen as very typical (Fig. 13, 14). Here, as in all other localities, the lower and middle parts of the Snyderville are unfossiliferous blocky clay having physical characters of an underclay and therefore interpreted as nonmarine, but the uppermost 1 or 2 feet consist of well-laminated gray shale in which Neochonetes abounds. The shells are comparatively thin and fragile but they are well preserved, most specimens (both single valves and conjoined valves) being essentially perfect. They outnumber associated invertebrates in the ratio of approximately 5,000 to 1. Toomey (1964) observed the Snydervilletype Neochonetes assemblage in connection with detailed studies made by him on the overlying Leavenworth Limestone and has reported that northeastward from the Toronto area a very gradual increase of faunal elements other than Neochonetes can be observed, though the chonetoids remain strongly dominant. In Cass County, Nebraska, he found moderately numerous specimens of Derbyia, Crurithyris, Meekella, Juresania, Dictyoclostus [= Antiquatonia], Neospirifer, echinoid and bryozoan remains, and a pelecypod associated with abundant Neochonetes.
Figure 13--Typical section of uppermost Lawrence Shale and most of Oread Limestone near Lawrence, Kansas, showing stratigraphic occurrence of some ecosystems ("B," Beil-type; "H," Heebner-type; "K," Kereford-type; "L," Leavenworth-type; "SN," Snyderville-type; "T," Tarkio-type) (Modified from Moore and Merriam, 1959). [Included in the Acrobat PDF file of figures for this article.]
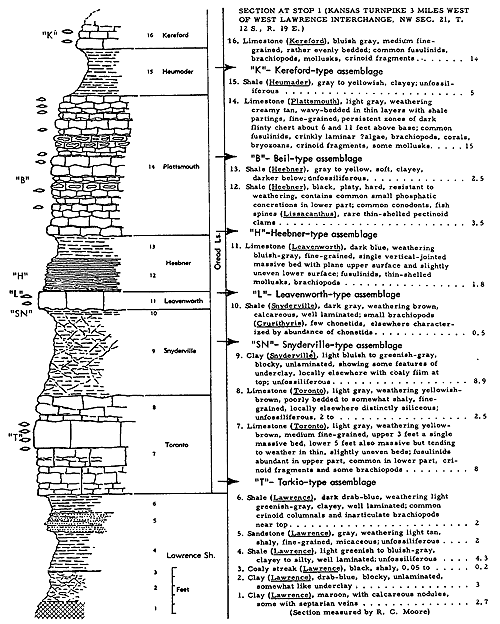
Figure 14--Typical section of uppermost Tecumseh Shale and most of Deer Creek Limestone south of Lecompton, Kansas, showing stratigraphic occurrence of some ecosystems ("B," Beil-type; "H," Heebner-type; "L," Leavenworth-type; "O," Ozawkie-type; "SN," Snyderville-type; "ST," Stranger-type) (Modified from Moore and Merriam, 1959). [Included in the Acrobat PDF file of figures for this article.]
Chonetoid assemblages which seem to correspond closely to the Neochonetes ecosystem of Snyderville type are dominated by Mesolobus and Chonetinella in various Desmoinesian cyclothems and by Chonetinella in Missourian cyclothems. Associated with these assemblages and with Neochonetes in Virgilian and Lower Permian beds at different horizons are chonetoid genera named Eolissochonetes, Lissochonetes, and Quadrochonetes; specimens of these may be common but nowhere are they found in swarms such as characterize shells of Neochonetes in the Snyderville-type assemblage.
The habitat of the chonetoid populations must have been in shallow water where a mud bottom bordered the shore, although perhaps in a belt many miles wide. Because the shale is a fine-grained deposit which is fairly well bedded and because the shells of Neochonetes are comparatively thin and fragile, disturbance of the bottom by currents and waves must have been little. Salinity may have been slightly subnormal, enough to be unfavorable for less euryhaline invertebrates, but almost surely salt content was greater than in brackish water.
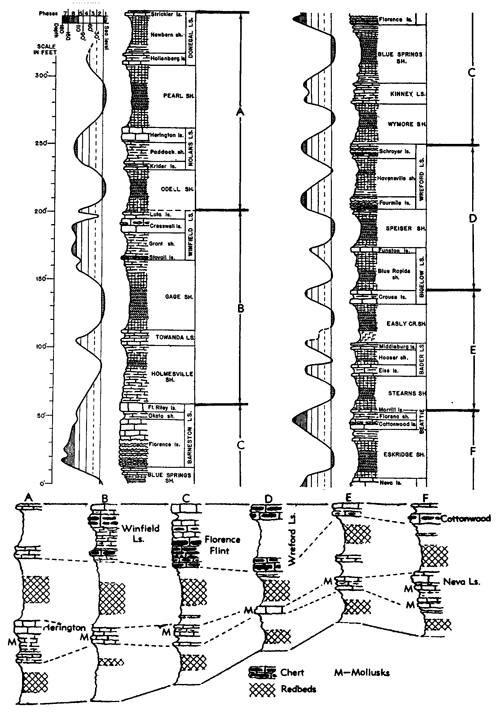
Very similar to the Snyderville-type ecosystem is the Lower Permian ecologic community of the Florena Shale Member of the Beattie Limestone as observed at most places along the outcrop. This assemblage occurs mainly in the lower few feet of the shale, next above the Cottonwood Limestone, the upper half which in northern Kansas and Nebraska is packed with slender fusulinids (chiefly Schwagerina). Therefore, the Florena assemblage, which commonly includes numerous to abundant free-weathering fusulinids in its basal few inches (and less conspicuously in higher parts of the member), is not like the Snyderville-type Neochonetes assemblage in being an initial marine community of a transgressing sea; the Florena deposits belong to the middle or regressive part of the Beattie cyclothem. At any rate, I judge it desirable to refer separately to a Florena-type Neochonetes-Derbyia assemblage (Suppl. Fig. 1, 2) and cite the very accessible and typical development of it on Kansas Highway 18 at the west edge of Manhattan as selected reference example (Fig. 15).
Figure 15--Typical section of Grenola, Eskridge, and Beattie strata just west of Manhattan, Kansas, showing stratigraphic occurrence of some ecosystems ("F," Florena-type; "M," Morrill-type; "RE," Red Eagle-type; "T," Tarkio-type; "TM," Threemile-type) (Modified from Moore and Merriam, 1959). [Included in the Acrobat PDF file of figures for this article.]
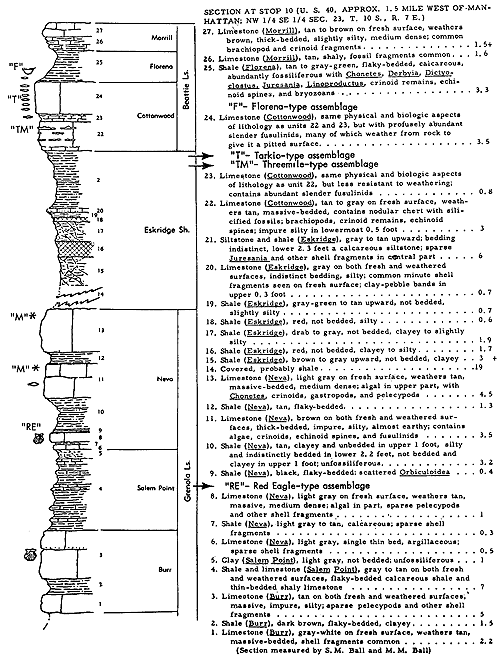
Information concerning both the FIorena fauna and varying composition of the enclosing sediment is unusually complete and detailed owing to quantitative studies by Imbrie (1955), who collected foot-by-foot samples, each weighing approximately 11 Ibs. (dry), from 24 measured sections distributed along the outcrop from Nebraska to Oklahoma. At each locality a representative collection of larger fossils was made and in the laboratory other fossils were obtained by washing and screening the samples. Also, the physical and chemical composition of each sample was determined analytically. Main results of the study have been reported and interpreted in terms of paleoecology and paleogeography (Imbrie, 1955; Imbrie, Laporte, and Merriam, 1959).
According to Imbrie, the invertebrate assemblage of the FIorena is dominated by brachiopods, among which he records Chonetes [= Neochonetes], Dictyoclostus [= Reticulatia], Juresania, Composita, Meekella, and Derbyia. My observations at many more places than reported on by Imbrie confirm this but Neochonetes is far more abundant than one may conclude from his data, partly because he measured abundance by weight in grams instead of specimen counts and partly because fossils surveyed by him were confined to those washed from his samples. Accumulations of fossils weathered from a shale ought to be representative of the fauna as a whole (unless differential transportation or solution operates to remove some kinds) but weathered-out fossils are ill-suited for bed-by-bed study of faunas. At most localities the FIorena yields a few fragments of crinoids (mostly single columnals or short stem fragments), echinoid spines and plates, parts of bryozoan zoaria (both fenestrate and ramose), and uncommonly a gastropod or two; Ditomopyge has been reported but except locally (e. g., near Grand Summit in Cowley County) trilobite remains are very rare. The Florena of southern Kansas exhibits a facies unlike that of central and northern parts of the state. This is discussed in a later part of the present paper devoted to a review of Beattie cyclothem biotopes.
A marine paleobiotope which is a characteristic and prominent one in many Pennsylvanian cyclothems but not recognized in others is marked by an unusually large fauna of varied invertebrates, which here is designated as the Pulchratia assemblage (Suppl. Fig. 2, 3). Its occurrences are indicated on stratigraphic diagrams by the symbol "B," which refers to its typical representation in the Beil Limestone Member of the Lecompton Limestone near Lecompton, Kansas, especially at outcrops in the south bluffs of Kansas River at Grover Station, approximately 3 miles west of Lecompton (Perkins, Perry, and Hattin, 1962), where large collections of fossils have been made (Fig. 13, 14, 16, 17). Specimens are exceptionally well preserved, few if any forms showing abrasions such as might be caused by being moved about by currents. Fragile bryozoans and delicate small brachiopods are unbroken; fine surface ornamentation, including granules, cancellate or parallel lineation, minute spines and lamellae, generally is intact. Collections commonly contain perfect specimens best suited for museum display.
Figure 16--Typical section of uppermost Kanwaka Shale and most of Lecompton Limestone southeast of Lecompton, Kansas, showing stratigraphic occurrence of some ecosystems ("A," Avoca-type; "B," Beil-type; "H," Heebner-type; "S," Speiser-type; "ST," Stranger-type; "T," Tarkio-type) (Modified from Moore and Merriam, 1959). [Included in the Acrobat PDF file of figures for this article.]
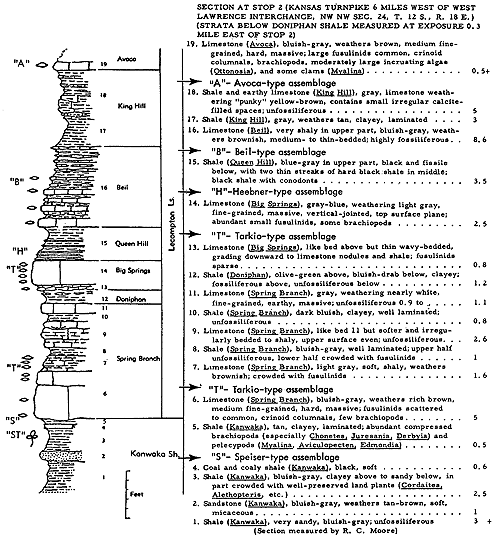
Figure 17--Correlated sections of Shawnee Group (Virgilian) units in southeastern Kansas and northern Oklahoma, showing stratigraphic occurrence of some ecosystems ("A," Avoca-type; "B," Beil-type; "DN," Doniphan-type; "H," Heebner-type; "L," Leavenworth-type; "O," Ozawkie-type; "P," Plattsmouth-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
Ecosystems of Beil type are relatively common and widely distributed in the Pennsylvanian part of the Kansas column above the Cherokee but are not well developed in most of the Permian section. They do occur, however, in the Council Grove Group (e.g., Hughes Creek-Foraker beds, Beattie Limestone of southern Kansas).
Naturally, the specific and generic composition of so-called Pulchratia assemblages differs both stratigraphically and geographcally and conditions favorable for collecting by no means are found at all outcrops of cyclothem units which are classifiable as representing this paleobiotope. Average good collections consist preponderantly of brachiopods; bryozoans, corals, and crinoid remains (cups and crowns generally rare) are commonly numerous and mollusks (small gastropods and pelecypods, chiefly) tend to be varied but not prominent; fusulinids may occur in profusion; pygidia and other remains of trilobites are almost invariably findable by searching. Most briefly described, abundance and varied composition of well-preserved invertebrates distinguish Pulchratia assemblages. Not to be overlooked is rather common occurrence of certain lamellose lettucelike types of algae.
Typical Beil-type (Pulchratia) assemblages are likely to contain a good representation of invertebrates and other organisms belonging to genera listed in the following tabulation (also partly illustrated in Suppl. Fig. 1-7).
| Organisms Found Commonly to Abundantly in Beil-type (Pulchratia) Assemblages | |
|---|---|
| Algae (relatively few as a rule but may include some Anchicodium or Archaeolithophyllum, incrustations of Ottonosia type, and scattered inconspicuous Osagia colonies) | |
| Invertebrates (strongly dominant, highly varied, exceptionally well preserved) | |
| Foraminifers: Fusulina, Fusulinella, Triticites, Wedekindellina, Waeringella, Dunbarinella, Pseudoschwagerina | |
| Corals: Aulophyllum, Axophyllum, Caninia, Cladochonus, Dibunophyllum, Lophophyllidium, Michelinia, Syringopora | |
| Bryozoans: Batostomella, Cystodictya, Fenestrellina, Fistulipora, Goniocladia, Meekopora, Meekoporella, Penniretepora, Polypora, Rhabdomeson, Rhombopora, Septopora, Streblotrypa, Tabulipora, Thamniscus | |
| Brachiopods: (Orthidina) Enteletes, Rhipidomella, Schizophoria, (Strophomenidina) Derbyia, Derbyoides, Meekella, Orthotetina, (Chonetidina) Chonetinella, Eolissochonetes, Lissochonetes, Mesolobus, Neochonetes, Quadrochonetes, (Productidina) Antiquatonia, Cancrinella, Desmoinesia, Echinaria, Hystriculina, Juresania, Kozlowskia, Linoproductus, Pulchratia, Retaria, Reticulatia (Suppl. Figs. 2, 3, 5), (Oldhaminidina) Poikilosakos, (Rhynchonellidina) Wellerella, (Retziidina) Hustedia, (Athyrididina) Cleiothyridina, Composita, (Spiriferidina) Crurithyris, Neospirifer, Phricodothyris, Punctospirifer, (Terebratulidina) Cryptacanthia, Dielasma | |
| Mollusks: (Gastropods) Bellerophon, Euphemites, Glabrocingulum, Ianthinopsis, Meekospira, Naticopsis, Phymatopleura, Soleniscus, Straparolus, Trachydomia, Trepospira, Worthenia (Suppl. Fig. 6, 7), (Bivalves) Acanthopecten, Allorisma, Aviculopecten, Edmondia, Leda, Myalina, Nucula, Nuculopsis, Parallelodon, Pinna, Pleurophorus, Pseudomonotis, Schizodus, Yoldia (Suppl. Fig. 4), (Cephalopods) Metacoceras, Pseudorthoceras | |
| Arthropods: (Trilobites) Ameura, Ditomopyge, Paladin (Suppl. Fig. 2), (Ostracodes) inconspicuous but common | |
| Echinoderms: (Crinoids) locally numerous dorsal cups, rare crowns, abundant fragmental remains, mostly columnals blit some plates, (Echinoids) Archaeocidaris spines and plates | |
| Vertebrates (generally rare fish teeth) | |
Brachiopods named in the foregoing list are grouped in suborders which are recognized in the Treatise on Invertebrate Paleontology; all have terminations in -idina. Paleontologists generally familiar with Pennsylvanian and Permian invertebrates of the Midcontinent region are likely to miss familiar generic names (e.g., Ambocoelia, Chonetes, Chonetina, Dictyoclostus, Echinoconchus, Marginifera, Productus, Pustula, Spiriferina, Squamularia--all now recognized as restricted to older rocks or to Permian outside of North America or replaced by senior synonyms) and they are called on to learn that many well-known species presently masquerade in new binominal combinations. This is illustrated by names of brachiopods selected from Condra and Dunbar (1932) accompanied by their modified equivalents as given by Muir-Wood and Cooper (1960), Muir-Wood (1962) and Treatise authors (Brachiopoda, Part H, 1964).
| Revised Nomenclature of Some Pennsylvanian and Permian Brachiopoda | |
|---|---|
| Dunbar and Condra (1932) | Various Authors (1960-1964) |
| Ambocoelia expansa D. & C. | Crurithyris expansa (D. & C.) |
| Ambocoelia lobata Girty | Crurithyris lobata (Girty) |
| Ambocoelia planoconvexa (Shumard) | Crurithyris planoconvexa (Shumard) |
| Chonetes granulifer Owen | Neochonetes granulifer (Owen) |
| Chonetes granulifer meekanus Girty | Neochonetes meekanus (Girty) |
| Chonetes granulifer transversalis, D. & C. | Neochonetes transversalis (D. & C.) |
| Chonetes granulifer armatus Girty | Neochonetes armatus (Girty) |
| Chonetina flemingi (Norwood & Pratten) | Chonetinella flemingi (N. & P.) |
| Chonetina flemingi alata D. & C. | Chonetinella alata (D. & C.) |
| Chonetina flemingi plebeia D. & C. | Chonetinella plebeia (D. & C.) |
| Chonetina verneuiliana (N. & P.) | Chonetinella verneuiliana (N. & P.) |
| Dictyoclostus portlockianus (N. & P.) | Antiquatonia portlockiana (N. & P.) |
| Dictyoclostus portlockianus crassicostata D. & C. | Antiquatonia crassicostata (D. & C.) |
| Dictyoclostus americanus D. & C. | Reticulatia americana (D. & C.) |
| Echinoconchus moorei D. & C. | Echinaria moorei (D. & C.) |
| Echinoconchus semipunctatus (Shepard) | Echinaria semipunctata (Shepard) |
| Echinoconchus semipunctatus knighti D. & C. | Echinaria knighti (D. & C.) |
| Juresania ovalis D. & C. | Pulchratia ovalis (D. & C.) |
| Juresania symmetrica (McChesney) | Pulchratia symmetrica (McChesney) |
| Krotovia meeki D. & C. | Pulchratia meeki (D. & C.) |
| Lissochonetes geinitzianus geronticus D. & C. | Quadrochonetes geronticus (D. & C.) |
| Lissochonetes geinitzianus plattsmouthensis D. & C. | Quadrochonetes plattsmouthensis (D. & C.) |
| Marginifera fragilis D. & C. | Hystriculina fragilis (D. & C.) |
| Marginifera haydensis Girty | Kozlowskia haydensis (Girty) |
| Marginifera hystricula D. & C. | Hystriculina hystricula (D. & C.) |
| Marginifera lasallensis (Worthen) | Retaria lasallensis (Worthen) |
| Marginifera missouriensis Girty | Desmoinesia missouriensis (Girty) |
| Marginifera muricatina D. & C. | Desmoinesia muricatina (D. & C.) |
| Marginifera splendens (N. & P.) | Kozlowskia splendens (N. & P.) |
| Marginifera wabashensis (N. & P.) | Hystriculina wabashensis (N. & P.) |
| Marginifera wabashensis armata D. & C. | Hystriculina armata (D. & C.) |
The paleobiotope represented by the Beil-type (Pulchratia) assemblage is thought to be one that existed in clear sunlit shallow waters (estimated less than 20 m. on the average) far from nearest shores (probably 50 to 100 miles distant). The environment is interpreted to belong in the culminating marine part of the cyclothem in which it is recorded by presence of the assemblage and the nature of the sediments (generally thin-bedded carbonate mud, now limestone) surrounding the fossils. I can suggest no diagnostic characters as basis for distinguishing Beil-type paleobiotopes from such others as are called Tarkio type (with Triticites assemblage), Wakarusa type (with Ottonosia-Reticulatia assemblage), and Avoca type (with Amblysiphonella assemblage) , even though each seems to possess persistent features of its own. At the present stage of paleoecological inquiry, it is judged desirable to treat the various types separately, rather than to lump them on the ground that all of those mentioned are characterized by common occurrence of fusulinids in the organic com. munity. Why the Beil-type ecosystems are marked by special abundance and variety of invertebrates in contrast with random distribution of a much smaller number and taxonomically restricted sea-bottom population of invertebrates is a problem for study, first by comparing as analytically as possible all recognized Beil-type assemblages among themselves. The same sort of comparisons of somewhat similar types with one another may ultimately lead to discrimination of sought-for paleoecological criteria.
An ecologic community which may be construed as merely a variant of the Beil-type (Pulchratia) assemblage is observed at several places in Kansas and Nebraska in such rock units as the Beil and Plattsmouth Limestones (Fig. 17, 18). It is characterized by abundance of corals, especially Caninia (Suppl. Fig. 4, 24) and Syringopora, and by a less varied fauna of brachiopods, bryozoans, and mollusks, all represented by fewer individuals than in the typical Pulchratia assemblage. Fusulinids generally are common, however. The coral-dominated ecosystem is named the Plattsmouth-type (Caninia) assemblage, mainly in order to avoid duplication of Beil-type as applied to both the coral-rich and Pulchratia assemblages. Species belonging to the mentioned coral genera are minor elements of most Pulchratia assemblages.
The most common marine ecologic assemblage of organisms found in Pennsylvanian and Permian cyclothems of Kansas consists predominantly, in many places almost exclusively, of fusulinid foraminifers. Genera represented, differing according to stratigraphic horizon, include Triticites (Suppl. Fig. 5, 7) most commonly and therefore the assemblage associated with this paleobiotope is named the Tarkio-type (Triticites) assemblage and the excellent sample of it seen in exposures of the Tarkio Limestone on US Highway 40 near the Shawnee-Wabaunsee county line is indicated as typical. Genera in other so-called Triticites assemblages include Kansanella, Fusulina, Fusulinella, Dunbarinella, Waeringella, Wedekindellina, Schwagerina, Paraschwagerina, and Pseudoschwagerina, as well as inconspicuous Schubertella, Millerella, Staffella, and Oketaella (Fig. 9-11, 13, 15-20).
Figure 18--Correlated sections of lower Shawnee (Virgilian) units in eastern Kansas, showing stratigraphic occurrences of some ecosystems ("H," Heebner-type; "L," Leavenworth-type; "P," Plattsmouth-type; "SN," Snyderville-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
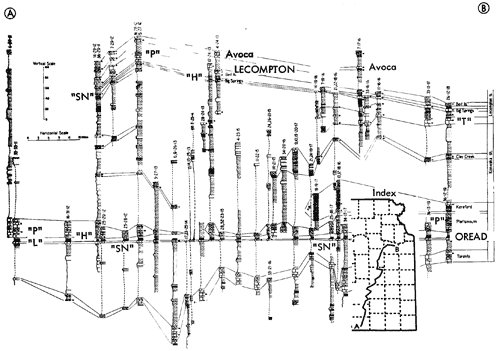
Figure 19--Correlated sections of lower Shawnee (Virgilian) units in northeastern Kansas and adjacent parts of Missouri and Nebraska, showing stratigraphic occurrences of some specified ecosystems ("A," Avoca-type; "B," Beil-type; "H," Heebner-type; "K," Kereford-type; "L," Leavenworth-type; "SN," Snyderville-type; "ST," Stranger-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
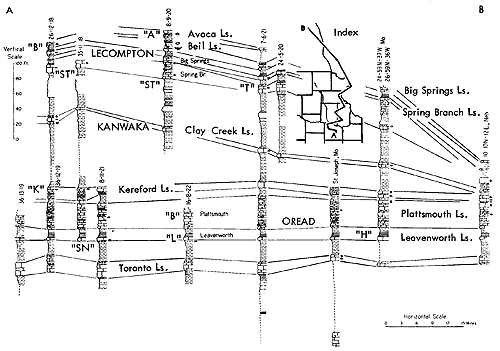
Figure 20--Correlated sections of Shawnee (Virgilian) units in northeastern Kansas and adjacent parts of Missouri, Iowa, and Nebraska, showing stratigraphic occurrences of some specified ecosystems ("A," Avoca-type; "B," Beil-type; "H," Heebner-type; "L," Leavenworth-type; "O," Ozawkie-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
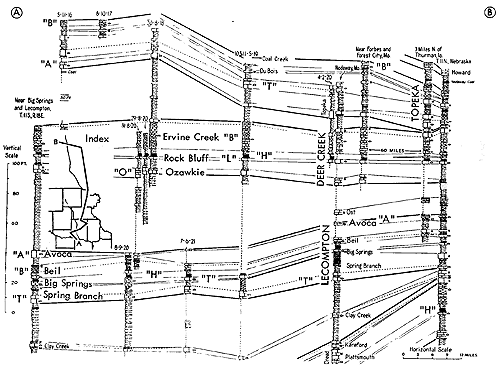
Some beds are mostly composed of fusulinids and in these the shells may be crowded so closely together that not only is room lacking for other invertebrates but not much space is occupied by rock matrix. Soft or shaly limestones of this type weather readily in manner that yields literally bushels of free specimens (e.g., Triticites ventricosus, Dunbarinella hughesensis) at some Hughes Creek Shale outcrops in central and southern Kansas; Dunbarinella glenensis at some Glenrock Limestone exposures in the same region (Fig. 9). The upper part of the Spring Branch Limestone and entire thickness of the Big Spring Limestone (both Lecompton) at many localities are largely formed by the shells of Triticites cullomensis (Fig. 10, 16-23). Large white specimens of T. ventricosus spread throughout massive brown Tarkio Limestone several feet thick give a porphyrylike appearance of groundmass and phenocrysts to the rock which is very striking, and virtually no other fossils are seen. Across nearly all of Kansas the upper half of the Cottonwood Limestone is a uniform massive layer of even-textured rock in which almost the only fossils are extremely abundant specimens of elongate, slender fusulinids (Schwagerina emaciata, S. jewetti). These are examples of Triticites assemblages and it is obvious that they must represent a distinctive sort of paleobiotope which recurred many times and generally was extremely uniform and widespread.
Figure 21--Correlated sections of upper Wabaunsee (Virgilian) units in eastern Kansas, showing stratigraphic occurrences of some specified ecosystems (location in Greenwood and Lyon Counties shown on index map, Figure 22) ("M," Morrill-type; "ST," Stranger-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
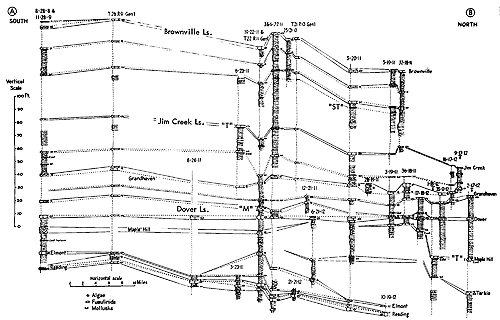
Figure 22--Correlated sections of upper Wabaunsee (Virgilian) units in eastern Kansas, showing stratigraphic occurrences of some specified ecosystems ("ST," Stranger-type; "T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
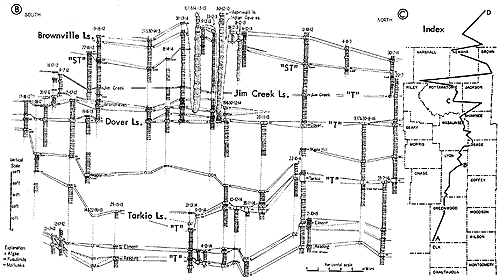
Figure 23--Correlated sections of upper Wabaunsee (Virgilian) units in northeastern Kansas and southeastern Nebraska, showing stratigraphic occurrences of some specified ecosystems (location shown on index map, Figure 22) ("T," Tarkio-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
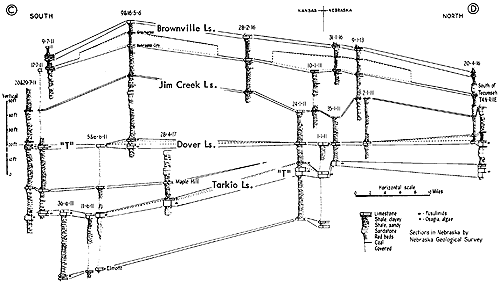
The fact that Triticites and other fusulinids are by no means restricted to so-called Triticites assemblages certainly indicates that paleobiotopes characterized by profuse occurrence of these foraminifers, so great in number that they seem to crowd out other invertebrates, probably were not radically different from shallow-sea environments in which the fusulinids were proportionally and actually much fewer. Because fusulinid-bearing beds almost invariably overlie and underlie marine deposits which lack them, these latter being discriminated on the basis of their cyclic relationships as belonging to initial or intermediate transgressive phases of inundation by sea waters on one hand or to retreatal phases on the other; the strata containing fusulinids are in the middle of the marine parts of cyclothems. At least this is true ideally. Triticites is not a marker of deepest parts of invading Pennsylvanian and Permian seas but presumably of intermediate to greatest distances from sea margins.
Both in Pennsylvanian and Permian parts of the Kansas column one may observe stratigraphically adjacent but obviously dissimilar ecosystems classifiable as Triticites assemblages (e.g., abundant T. ventricosus, a very robust form, as almost exclusive constituent of the Tarkio Limestone assemblage, followed a few feet higher by the Maple Hill Limestone assemblage containing exclusively small and slender Triticites). Clearly, the Maple Hill fusulinid population is not composed of lineal descendants of Tarkio stocks (which presumably do appear in T. ventricosus of the Hughes Creek, Grenola, and some other Lower Permian units). Question arises whether Maple Hill paleoecology was different enough from that of the Tarkio sea to explain the faunal dissimilarity and one may ask where the post-Tarkio populations of T. ventricosus came from, since this species was not a continuous inhabitant of the Kansas region in late Pennsylvanian and early Permian time. Such problems have been considered previously by me, arriving at the conclusion that fusulinids of successive cyclothems mostly do not show stages in the evolution of identifiable lineages which contributed to the paleontological record in Kansas by their presence in repeated marine invasions (Moore, 1954).
It is also noteworthy that in some cyclothems (e.g., Foraker, Beattie, discussed by Elias, 1962, p. 108-110; Red Eagle, reported by McCrone, 1963, p. 64, 96-105) fusulinid-rich beds alternate with layers containing more or less abundant fossils of other sorts, mainly brachiopods, but lacking fusulinids. Oscillatory changes in local sea-bottom environments should not be considered exceptional and surely they do not demand explanation in terms of significant movements of nearest strand lines or of appreciable changes in water depths. The successive fusulinid populations in examples of this sort generally are similar in generic and specific composition.
The Cottonwood Limestone was cited in the first part of the discussion of Triticites assemblages as an excellent example, although fusulinids other than Triticites make up the Cottonwood community. This is given attention in a later part of the present paper in a section devoted to review of biotopes of the Beattie cyclothem.
An ecologic community of organisms which seems clearly to differ from those here designated as Beil-type (Pulchratia) assemblage and Tarkio-type (Triticites) assemblage but nevertheless corresponding to them as regards position in the marine part of cyclothems is considered next. It is named the Wakarusa-type (Ottonosia-Reticulatia) assemblage and among several examples of it which might be chosen, its occurrence as seen in outcrops of the Wakarusa Limestone on US Highway 54 approximately 3 miles west of Eureka, in Greenwood County, is cited for reference (Fig. 24, 25). Main observed features are moderately large specimens of the productidine genus Reticulatia (Suppl. Fig. 5, 10), partly to almost entirely incrusted by laminated algal-foraminiferal colonies typically representative of Ottonosia. Such specimens are well scattered at random in the fine granular matrix of the rock and they seem to display no consistent orientation. They are not crowded together in any way. The growths of Ottonosia are quite variable in thickness, in different colonies or parts of the same colony, ranging from approximately 2 to 30 mm. A tendency toward maximum coverage and thickness of the colonies on upper surfaces of their support indicates growth on shells fairly well stabilized in position on the sea floor and consequently an absence of strong currents or wave-induced turbulence. Associated fossils include several other genera of brachiopods, a few mollusks (both snails and bivalves), some bryozoans and rare corals, and sparse to common fusulinids and crinoid remains (chiefly stem fragments). Ottonosia may be found on various shells and incrusting the crinoid stems on all sides, covering even the articular surface at one or both ends. Of course, this proves that such stems were disarticulated when the Ottonosia colonies were developing around them, as they might on occasionally rolled loose pebbles. Specimens of Osagia have been observed but are not common constituents of the Wakarusa-type assemblage.
Figure 24--Correlated sections of Wabaunsee (Virgilian) units in northeastern Kansas, showing stratigraphic occurrences of some specified ecosystems (location indicated on inset map, Figure 25) ("T," Tarkio-type; "W," Wakarusa-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
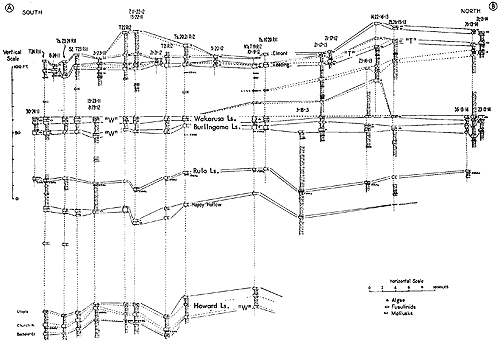
Figure 25--Correlated sections of Wabaunsee (Virgilian) units in northeastern Kansas, showing stratigraphic occurrences of some specified ecosystems ("T," Tarkio-type; "W," Wakarusa-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
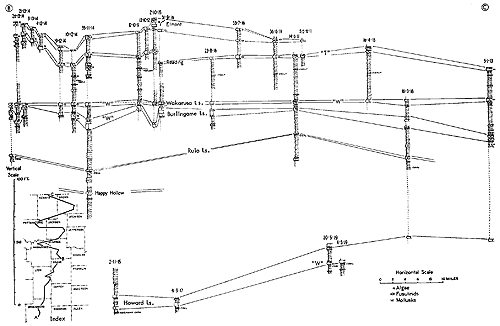
Assemblages which closely resemble that widely distributed in the Wakarusa cyclothem, duplicating it in fact, are found in the Church cyclothem of the Howard Limestone and the Avoca cyclothem of the Lecompton Limestone, to mention only two examples. Paleoecological research obviously calls for detailed comparisons of these stratigraphically well-separated occurrences, and it needs also to explore resemblances and differences to be found in comparing Wakarusa-type assemblages with such Ottonosia- and Reticulatia- or Antiguatonia-containing communities as Beil- and Plattsmouth-type assemblages and others. Large algal "biscuits" of Ottonosia or Somphospongia with initial growths on very inconspicuous supports are seen, for example, on the upper surface of the Burlingame Limestone, Howe Limestone, etc., at least locally associated with abundant Osagia (McCrone, 1963, p. 30). What environmental conditions do these signify? Johnson (1946, 1963) has described and illustrated Pennsylvanian and Permian algae extensively, including numerous forms from Kansas, but unhappily has written virtually nothing (less than 6 lines) on paleoecological interpretation of them.
Observations on geographic variations in composition of the Wakarusa Limestone assemblage previously have been noted' by Moore (1949, p. 10). In northern Kansas this organic group is dominated by robust fusulinids in association with common large productidine brachiopods (Reticulatia), fistuliporoid and other bryozoan colonies, crinoid remains, and the like, Ottonosia being present but not very common. In central Kansas fusulinids are less numerous and other elements of the assemblage, including Ottonosia, are more prominent. In southern Kansas, the Wakarusa, readily and almost continuously traceable along the outcrop, is found to have still fewer fusulinids, but more numerous colonies of Ottonosia. This trend continues into Oklahoma where fusulinids and most other invertebrates vanish from the Wakarusa, whereas Ottonosia is the most conspicuous fossil, so much so that this rock unit came to be known in early stratigraphic classification of northern Oklahoma Pennsylvanian formations as the Cryptozoon.bearing Limestone, a member of the so-called Buck Creek Formation (Beckwith, 1928, p. 12). The Cryptozoon referred to is the same as Ottonosia. Its prominence and usefulness as a horizon-marker were first reported by Heald (1919, p. 215) . The geographic variations reported must have paleoecological significance, in my opinion indicating that the Wakarusa-type assemblage of northern Kansas, with abundant Triticites and relatively few Ottonosia, represents a paleobiotope in a far offshore locacation, whereas the presumably contemporaneous Ottonosia-dominated, fusulinid-Iacking assemblage in the Wakarusa Limestone of Osage County, Oklahoma, denotes a paleobiotope located comparatively nearshore, possibly less than 30 miles or so from a strand line to the south.
The sponge named Amblysiphonella (Suppl. Fig. 1, 21) is a marker of marine units in some Kansas Pennsylvanian cyclothems, occurring within or rather commonly on the underside of limestone beds (e. g., Avoca Limestone, Fig. 10, 16-20; lowermost part of Topeka Limestone, Fig. 20). A so-called Avoca-type (Amblysiphonella) assemblage is distinguished by the presence of this genus and a reference example of it is cited from good exposures of the Avoca Limestone in a small east-flowing creek bed and banks 2 miles north of Big Springs. The containing rock is almost invariably a massive limestone ranging in thickness from 1 to 5 or 6 feet. The fact that moderately large fusulinids, robust Antiquatonia, crinoid columnals, and prominent algal-foraminiferal colonies of Ottonosia are characteristically associated with the occurrence of Amblysiphonella indicates that the paleobiotope represented by the assemblage is closely comparable to some others which here have been classed as marked by the Wakarusa-type (Ottonosia-Reticulatia) assemblage (e.g., Church Limestone Member of the Howard Limestone and Wakarusa Limestone (Fig. 21, 22). Possibly no significant paleoecological distinctions in the examples cited warrant separation of one from another. At any rate, the Avoca-type assemblage is very typically developed in central and southern Kansas, as well as in northern Kansas and Nebraska.
Cherty and chalky limestone units which are very persistent characterize the Chase Group of the Kansas Lower Permian (e.g., Wreford, Barneston, Winfield Limestones). They are mainly composed of comminuted invertebrate remains and because of their very low content of clastic sediment are judged to represent culminating marine phases of cyclothems. In this respect they are interpreted as homologous with Pennsylvanian deposits such as the Plattsmouth Limestone (also commonly chert-bearing) characterized by its content of the Pulchratia assemblage. The Permian limestones referred to are only sparsely fossiliferous at most outcrops, however, and the fauna found in them is neither varied nor even locally marked by abundance of individuals, though fragmentary remains of benthonic organisms may be extremely common. The brachiopod Composita (Suppl. Fig. 5, 1) can be seen at virtually every exposure and fenestrate bryozoans are ubiquitous. The Threemile Limestone Member of the Wreford Limestone is selected for reference and the name Threemile-type (Composita-Fenestrellina) assemblage is applied to the whole community of organisms, including both relatively complete and very fragmentary remains (Fig. 26). Fusulinids (chiefly Triticites and Dunbarinella) are abundant in parts of the Florence Limestone (lower Barneston) but generally this group of fossils is absent. Scattered spines and plates of echinoids are characteristic.
Figure 26--Generalized sections of Lower Permian rocks extending upward from those given in Figure 11, showing stratigraphic occurrence of some ecosystems ("S," Speiser-type; "TM," Threemile-type) (Moore). [Included in the Acrobat PDF file of figures for this article.]
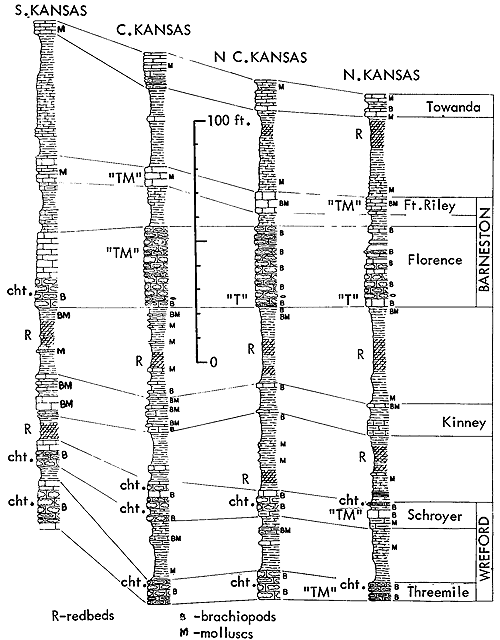
The most comprehensively studied Composita-Fenestrellina assemblages are those of the Wreford Limestone (Hattin, 1957) and these are reported to contain the following organisms.
| Composition of Threemile (Composita-Fenestrellina) Assemblages (Hattin, 1957) | |
|---|---|
| Algae (Girvanella, sparse algal-foraminiferal Osagia) | |
| Invertebrates (generally sparse, locally common, many fragmentary) | |
| Foraminifers: Ammodiscus, Ammovertella, Globivalvulina, Tetrataxis, Schwagerina (single small area in central Chase County) | |
| Sponges: siliceous spicules | |
| Corals: Aulopora, Dibunophyllum, Stereostylus (very locally in reef-like masses) | |
| Bryozoans: Fenestrellina, Penniretepora, Polypora, Rhombopora, Septopora, Streblotrypa, Tabulipora, Thamniscus, encrusting cyclostomes | |
| Brachiopods: Chonetes [= Neochonetes], Composita, Derbyia, Dictyoclostus [= Reticulatia], Enteletes, Orbiculoidea, Wellerella | |
| Mollusks: Aviculopecten, Aviculopinna, Septimyalina, small gastropods (unidentified) | |
| Arthropods: Bairdia, other unidentified ostracodes, trilobite (Ditomopyge) | |
| Echinoderms: Delocrinus, crinoid stem fragments, echinoid spines and plates | |
| Vertebrates (fragmentary remains, ?fish teeth) | |
According to Hattin (1957, p. 69, 92) the coral component of the Threemile assemblage deserves emphasis because at least a few specimens can be found at almost every exposure of the Wreford Limestone and they are abundant in the uppermost 12 inches or so of the Threemile Member at a few places. The prevailingly fine granular texture of the limestone indicates a high degree of disintegration of sea-bottom organisms, yet a considerable part of the material is identifiable to the extent of recognizing derivation from bryozoans such as Fenestrellina, echinoderms (both crinoids and echinoids), brachiopods, and mollusks. The sediment closely corresponds to the lower part of the Cottonwood Limestone called bioclastic by Laporte (1962), for example, which also is thought to represent an accumulation of organic remains far from shore. The agency or agencies of fragmentation are undetermined.
Isogramma (Suppl. Fig. 5, 4) is a distinctive but rare brachiopod and therefore its use for designating numerous puzzling organic assemblages and their paleobiotopes in the Pennsylvanian part of the Kansas rock column is decidely inappropriate if widespread occurrence and even local abundance are considered to be criteria for choosing a name. Actually, the only known specimens of Isogramma from Kansas come from the basal part of the Rock Bluff Limestone Member of the Deer Creek Limestone at localities in Osage County south of Topeka. It is the assemblage and biotope represented by the limestone containing this fossil that I wish to discriminate and it happens that generic names applied to other organisms in the Rock Bluff, Leavenworth, and homologous units are too general in occurrence elsewhere and too nondiagnostic of these assemblages to offer a good alternative name. Isogramma is adopted because it is unique and because it calls attention to neglect in paleontological study of this and closely similar stratigraphic units (e.g., Middle Creek. DuBois, etc., see, Fig. 10, 13, 14, 17-20). The Rock Bluff Limestone is an excellent example of deposits containing this assemblage for which reference exposures on US Highway 75 just north and south of Dragoon Creek, 5 miles north of Lyndon, in Osage County, are cited. There Isogramma has been collected. The excellent exposures of the Leavenworth Limestone on the Kansas Turnpike northwest of Lawrence are indicated as typical of this unit.
Physical characters of rock units classed as representing the biotope of the Isogramma assemblage are specially important, seemingly more so than the nature of organic remains contained in them, as evidence of the environmental conditions which need to be understood. All examples of these units are dense, brittle, rather dark bluish, only slightly ferruginous limestones which lack perceptible bedding and which very rarely exceed 2 feet in thickness anywhere. They crop out as single massive layers with knife-sharp lower and upper boundaries, that at the base uneven but that at the top so smoothly even as to give the appearance of having been expertly planed. Another feature is uniformity along the outcrop throughout air-line distances of 200 or more miles and some examples (e.g., Leavenworth Limestone) have been identified as equally persistent in down-dip direction approximately normal to the trend of outcrops (Fig. 27, 28). Truly remarkable as stratigraphic entities, they are equally remarkable as paleobiotopes, providing unusual demands on paleoecologists who try to interpret them accurately.
Figure 27--Correlated members of Oread Limestone penetrated in 16 northwestern Kansas wells, based on gamma-ray logs furnished by Lane-Wells Company, Wichita, Kansas. Especially noteworthy is the subsurface persistence of thing units such as Heebner Shale and Leavenworth Limestone (after Moore, 1950). [Included in the Acrobat PDF file of figures for this article.]
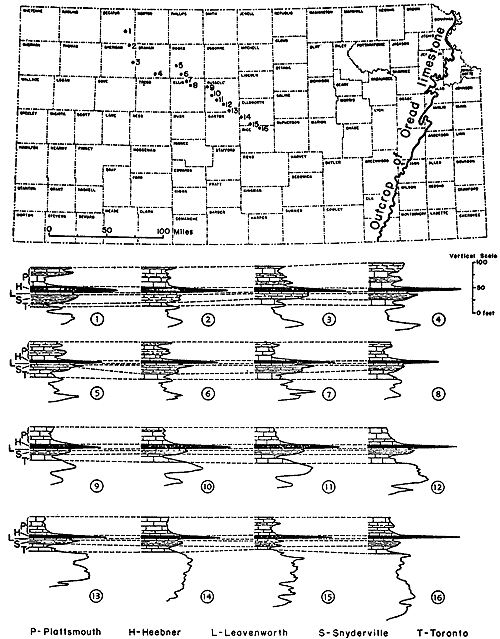
Figure 28--Surface and subsurface sections of major part of Oread megacyclothem in Kansas, Missouri, Nebraska, and Iowa, showing persistence of cyclothem elements (after Moore, 1950). [Included in the Acrobat PDF file of figures for this article.]
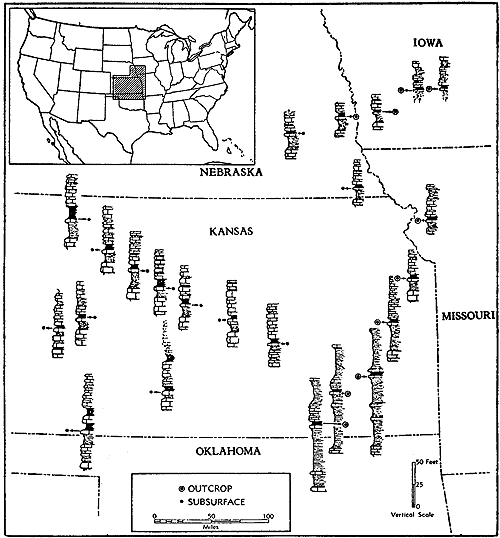
Invertebrates commonly present in Leavenworth-type assemblages are sparsely distributed like frugal admixtures of currants and raisins well stirred into a cake. They consist of rather small fusulinids, small gastropods, and scattered small brachiopods. Algal-foraminiferal incrustations of Osagia type occur but are nowhere abundant. On the whole, fossils are difficult to collect, for they neither weather out on exposed surfaces of the limestone nor break out cleanly from the rock matrix, tending almost invariably to fracture through the shells.
A recently completed study of the Leavenworth Limestone, which is a very typical representative of paleobiotopes characterized by the so-called Isogramma assemblage, is reported in an as yet unpublished doctoral dissertation by Toomey (1964). This contains abundant detailed information which confirms the general descriptive statements made in preceding paragraphs. Toomey worked on samples of the Leavenworth collected from 33 localities distributed along the outcrop from northern Oklahoma to northeastern Kansas and beyond in Missouri, Nebraska, and Iowa. Aided by computer analysis of lithologic and paleotologic data he was able to determine objectively that the Leavenworth unit has exceptional lateral homogeneity, consisting everywhere (other than 6 localities near southern and northern extremities of the outcrop) of comparatively pure calcareous skeletal mudstone and exhibiting extreme uniformity of organic content. In addition to study of fossils observed in 68 selected 2- by 3-inch thin sections and numerous smaller ones and on insoluble microfossils in residues, approximately 80 hours of searching for macrofossils at 24 localities, both by scanning weathered surfaces and by breaking up large quantities of rock, fewer than 600 specimens of mostly fragmentary invertebrates were collected. Assuredly, fossils are not very numerous and they are difficult to obtain. Though most of Toomey's identifications were not carried beyond the generic level, and though many of them, owing to incompleteness or poor preservation of the material and small number of available specimens, were uncertain even as to genera, his paleontological observations are valuable. The variety of reported organisms is summarized in the following tabulation.
| Organic Remains Reported by Toomey (1964) from Leavenworth Limestone of Kansas and Adjoining States | |
|---|---|
| ALGAE (quantitatively unimportant as rock-builders) | |
| Osagia (all localities), Ottonosia, Archaeolithophyllum, Epimastopora, Eugonophyllum, Girvanella, Tubiphytes | |
| INVERTEBRATES (generally sporadic in distribution, not abundant) | |
| Foraminifers: Triticites (most common fusulinid), Millerella, Staffella, Waeringella, Kansanella, Hedraites, Tetrataxis, Climacammina, Bradyina, Globovalvulina, Hemigordius, Syzrania, Endothyra, Tuberitina, Apterrinella, arenaceous forms (Textularia, Minammodytes, Bigenerina, Hyperammina, Thurammina, Ammovertella, Rheophax, Psammosphaera) | |
| Sponges: Coelocladia? (several locally) | |
| Corals: Lophophyllidium (rare, Iowa and Oklahoma only), Dibunophyllum (single specimen, Iowa) | |
| Bryozoans: encrusting cyclostomes (undetermined), ramose and fenestrate fragments (undetermined) | |
| Brachiopods: represented at all localities, none common; Cancrinella, Chonetinella?, Composita, Condrathyris [= Phricodothyris], Derbyia, Dictyoclostus [= Antiquatonia], Echinoconchus [= Echinaria], Enteletes, Linoproductus, Marginifera [= Desmoinesia (?misidentification), Kozlowskia], Meekella, Neospirifer, Wellerella | |
| Gastropods: Euphemites?, Naticopsis, Palaeostylus (Pseudozygopleura), Straparolus (Euomphalus), Worthenia | |
| Pelecypods: unidentified fragments | |
| Cephalopods: fragments of 2 large nautiloids | |
| Arthropods: ostracodes uncommon; trilobite (Ditomopyge) rare | |
| Worms: scolecodonts (unidentified) | |
| Conodonts: all rare, Streptognathodus, Spathognathodus, Hindeodella, Cavusgnathus, Prioniodina, Ozarkodina (Suppl. Fig. 8) | |
| Vertebrates (rare fish teeth, generically unidentified) | |
Toomey's discussion of this assemblage emphasizes its general uniformity in most of the study region and suggests that the calcareous mud substrate probably was not well suited for a majority of bottom-dwelling invertebrates which otherwise might be expected to occur.
The presence of fusulinids as moderately common constituents throughout the entire known geographic extent of several (but not all) Leavenworth-type assemblages raises question concerning homology within respective cyclothems of the units here discussed and several others, such as the Tarkio-type (Triticites) assemblage, Beil-type (Pulchratia) assemblage, and Wakarusa-type (Ottonosia-Reticulatia) assemblage. If all of those mentioned represent culminating epochs of marine transgression, their obvious differences need explanation in paleoecological terms. I cannot offer this.
An ecosystem termed Captain Creek-type (Enteletes) assemblage is discriminated on the basis of especially common occurrence of this brachiopod, in many places associated with Hystriculina hystricula and a few other forms (Suppl. Fig. 5, 2, 3). It is most characteristic of the Captain Creek Limestone Member of the Stanton Limestone (upper Missourian; Fig. 9, 29) and excellent examples are furnished by outcrops along Kansas Highway 10 at Captain Creek, east of Eudora, in Douglas County, and north of Eudora. Fusulinids generally are lacking but they are moderately abundant constituents of the assemblage in some places, especially in southern Kansas. A peculiarity is the manner in which perfect or nearly perfect specimens of Enteletes, Hystriculina, and other brachiopods can be broken out of the brittle limestone matrix, like shelling out the kernels of nuts. Paleoecological inquiry needs comparison of the Enteletes assemblage with those classed as Isogramma assemblages, for both occur next below fissile black platy shale of Heebner type. Also, comparison with Leavenworthtype paleobiotopes represented by such beds as the Merriam Limestone (Fig. 7, 29) and the Paola Limestone (Fig. 30, 31) is desirable because these units also underlie Heebner-type black shales. All of the limestones mentioned except the Captain Creek are uniformly thin but very persistent, whereas the Captain Creek is variably and locally thick.
Figure 29--Correlated sections of Lansing (Missourian) strata in southeastern Kansas, showing occurrence of some ecosystems ("C," Captain Creek-type; "L," Leavenworth-type) (Moore, sections mostly measured by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
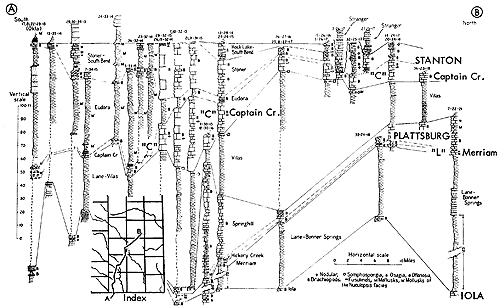
Figure 30--Correlated sections of Kansas City Group (Missourian) in eastern Kansas, showing occurrence of specified ecosystems (index map serves also for Figure 31) ("D," Drum-type; "L," Leavenworth-type; "ST," Stranger-type) (Moore, sections measured by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
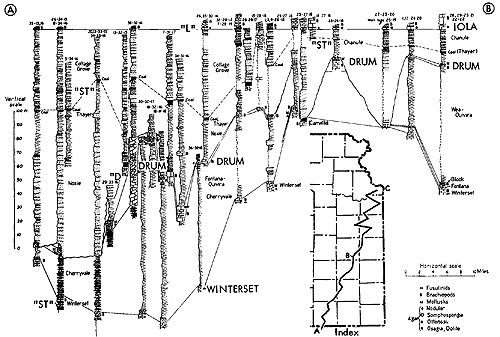
Figure 31--Correlated sections of upper Kansas City and Lower Lansing units in eastern Kansas, showing occurrence of specified ecosystems (index map included in Figure 30) ("B," Beil-type; "D," Drum-type; "L," Leavenworth-type; "ST," Stranger-type) (Moore, sections measured by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
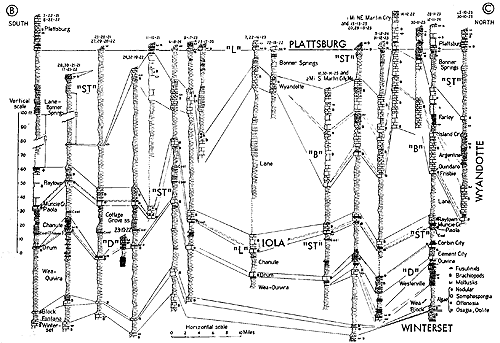
A community of marine invertebrates in which the lacelike fronds of fenestrate bryozoans, including Fenestrellina (Suppl. Fig. 1, 22; 5, 9), are very common is designated as the Kereford-type (Fenestrellina) assemblage. Examples of its occurrence in Pennsylvanian deposits of Kansas are indicated in accompanying correlated sections (Fig. 13, 19). A type reference section of Kereford beds containing the assemblage can be seen in a quarry at the northeast edge of Lecompton, in Douglas County, but well-preserved zoaria of Fenestrellina and other bryozoans are more numerous on the surfaces of slabby limestone of the Kereford near Waverly ("Waverly flags"), in northeastern Coffey County. Fusulinids occur locally in some Kereford-type assemblages but generally they are lacking.
Most common invertebrate associates of the bryozoans are varied kinds of brachiopods and mollusks, especially pelecypods and gastropods. Fossils are found to be better preserved than on the average, so that many very perfect specimens can be collected. The environment seems generally most like that marked by the Pulchratia assemblage, but waters are judged to have been somewhat nearer shore and a muddy bottom is indicated by greater proportion of interbedded shale and a common clayey to silty nature of slabby limestone beds. However this may be, conditions favorable for growth of large populations of bottom-dwelling invertebrates are denoted by the average abundance of individuals.
Early in studies of the biological associations found to characterize different parts of Kansas Pennsylvanian cyclothems, deposits interpreted as locally marking a late stage in the retreat of shallow seas were discriminated as "algal-molluscan" (Moore, 1936a, p. 25-32). Such beds overlie strata containing a varied assemblage of brachiopods, corals, crinoids, bryozoans ("mixed fauna"), some with associated fusulinids, or the algal-molluscan deposits were observed to occur next above fusulinid-packed limestone like that here named Tarkio-type. Similar sequences were less clearly recognized in Permian cyclothems (Elias, 1937a, p. 411). An assemblage here designated as Drum-type (Euconospira) assemblage illustrates a moderately late retreatal stage in Missourian cyclothems (Euconospira, Suppl. Fig. 6, 26). The fossils referred to are in the upper member (Corbin City) of the formation, which is a cross-bedded oolite of varying thickness (as much as 50 feet near Independence, Montgomery County); the underlying Dewey Limestone Member (= Cement City) of the Drum is dissimilar to the Corbin City Limestone lithologically, faunally, and in its much more uniform thickness (Fig. 30-32). Outcrops near Corbin City, 2 miles south of Cherryvale, contain a good representation of the Drum-type assemblage in which the fossils generally are near-perfect specimens. Places where they occur abundantly have been favorite localities for collectors.
Figure 32--Correlated sections of Missourian strata in eastern Kansas, showing some occurrences of specified ecosystems (assemblages) ("B," Beil-type; "D," Drum-type; "H," Heenner-type; "ST," Stranger-type) (Moore, sections mostly measured by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
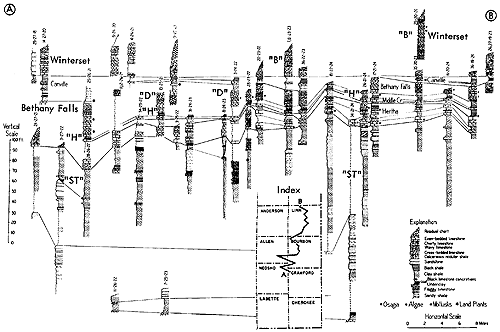
The oolite grains of the Corbin City Limestone consist of finely laminated, soft calcium carbonate deposited around a minute organic or inorganic particle, and partly because similar coatings cover the outside (but not inside) of many shells, algae of some sort, or possibly bacteria, are thought to have secreted the carbonate. The very common occurrence of cross bedding indicates both shallow water and current action. Cross-bedded to relatively massive oolitic limestone containing the Euconospira assemblage is especially well developed in the Drum Limestone of southeastern Kansas but is found also elsewhere (e.g., Westerville, upper Bethany Falls, upper Drum of the Kansas City area) (Fig. 31). Actually, many fossils described and illustrated by Sayre (1930) as constituents of the Drum fauna were obtained from outcrops now known to represent units beneath the Drum. Even so, all may be included in the Euconospira assemblage (many illustrated in Suppl. Fig. 1-7).
A composite list of Euconospira-assemblage invertebrates by genera is inadequate for characterization of the ecosystem because relative abundance of the different components is not indicated. Sayre (1930, p. 80) has reported that in northern areas diminutive ("dwarfed") mollusks predominate, whereas normal-sized to moderately robust specimens, including more numerous brachiopods, are found in: the south.
| Invertebrates Common in Drum-type (Euconospira) Assemblages (mainly compiled from Sayre, 1930, with nomenclatural revisions) | |
|---|---|
| Corals: Lophophyllidium, Michelinia | |
| Bryozoans: Fenestrellina, Fistulipora, Polypora, Rhabdomeson, Rhombopora, Septopora, Tabulipora | |
| Brachiopods: Chonetinella, Composita, Derbyia, Dielasma, Echinaria, Hustedia, Juresania, Linoproductus, Neospirifer, Phricodothyris, Pulchratia, Punctospirifer, Wellerella | |
| Mollusks: (gastropods) Anematina, Anomphalus, Araeonema, Baylea, Bellerophon (Bellerophon), B. (Pharkidonotus), Ceraunocochlis, Cinclidonema, Cyclozyga, Donaldina, Eucochlis, Euconospira, Euphemites, Glabrocingulum (Glabrocingulum), G. (Ananias), Glyphodeta, Goniasma, Gosseletina, Hemizyga (Hemizyga), H. (Hyphantozyga), H. (Plocezyga), Hypselentoma, Ianthinopsis, Knightites (Cymatospira), K. (Retispira), Leptoptygma, Meekospira, Microdoma, Microptychia, Murchisonia (Murchisonia), Naticopsis (Naticopsis), N. (Jedria), Orthonema, Palaeostylus (Palaeostylus), P. (Leptozyga), Patellilabia, P. (Pseudozygopleura), P. (Stephanozyga), Patellilabia, Soleniscus, Spiroscala, Stegocoelia, Trachydomia, Vernelia, Worthenia; (bivalves) Acanthopecten, Allorisma, Astartella, Aviculopecten, Aviculopinna, Clavicosta, Conocardium, Cypricardinia, Deltopecten, Edmondia, Euchondria, Fasciculiconcha, Leda, Limatula, Lithophaga, Monopteria, Myalina (Myalina), M. (Myalinella), M. (Orthomyalina), M. (Selenimyalina) , Nucula, Nuculopsis, Parallelodon, Pernopecten, Pleurophorus, Promytilus, Pseudomonotis, Schizodus, Solenomya, Streblochondria, Streblopteria, Yoldia; (cephalopods) Discites, Ephippioceras, Gonioloboceras, Metacoceras, Mooreoceras, Pseudorthoceras, Schistoceras, Tainoceras | |
| Arthropods: (trilobites) Ameura | |
| Echinoderms: (crinoids) fragments, columnals and plates; (echinoids) Archaeocidaris spines and plates | |
An ecologic community which is somewhat comparable to the Drum-type assemblage just discussed has been observed in the upper part of the Ozawkie Limestone Member of the Deer Creek Limestone, especially in outcrops adjacent to the Kansas River valley. It is designated as the Ozawkie-type (Knightites) assemblage (Fig. 14, 17, 20). Greatly predominant in numbers are small-to medium-sized colonies of Osagia, which compose a major part of the rock, 2 to 4 feet thick. Distributed in random manner but locally common are moderately robust gastropods and a few large bivalves, which, like fossils of the Drum-type algal-molluscan assemblage, are unusually well preserved (Moore, 1941), not only showing surface features in minute detail but allowing very useful studies of shell structure. The gastropods include species of Bellerophon (Bellerophon), Euphemites, Warthia, and Knightites (Knightites) (Suppl. Fig. 7, 15), which have been described and illustrated (Moore, 1941), but the bivalves have not been studied as a group. It is noteworthy that brachiopods and bryozoans seem to be lacking.
The paleobiotope represented by the Ozawkie-type (Knightites) assemblage was at least moderately extensive, for the Osagia-rich upper part of the Ozawkie member can be followed for a distance of 60 miles or more along the outcrop. So far as known, Knightites, and mollusks associated with it, is more local in distribution. The lower Ozawkie is a fusulinid-rich rock of Tarkio-type and is considered to mark culminating marine conditions within the Ozawkie cycle. The Knightites assemblage, with associated Osagia, doubtless lived in marginal parts of the retreating Ozawkie sea, though at undeterminable distance from the nearest strand line.
Ecologic communities of organisms which here are termed Spring Hill-type (Archaeolithophyllum) assemblages are especially interesting and seemingly they are not unusual in cyclothems of Pennsylvanian and Permian age in Kansas. Further, they are important as local rockbuilders [e.g., in the Lansing Group, upper Missourian, of southeastern Kansas and northeastern Oklahoma (Merriam, 1962; Wilson, 1962) and in the Council Grove Group, Lower Permian, of central Kansas (Fig. 29)]. Harbaugh (1959, 1960, 1962) has described deposits associated with and largely formed by these organisms as marine banks, which are well displayed by exposures in Montgomery and Wilson Counties northwest of Independence and west of Neodesha and Altoona. More recently Wray (1964) has described and illustrated species of the algal genus Archaeolithophyllum (Suppl. Fig. 8, 14) collected from the Lansing marine banks in the area mentioned. Localities selected as type examples of Archaeolithophyllum assemblage occurrences are (1) on Kansas Highway 96 in sec. 23, T. 30, S., R. 15 E., 3 miles west of Neodesha, and (2) on Kansas Highway 47 in sec. 18, T. 29 S., R. 16 E., 2 miles west of Altoona; at both places the assemblage is developed in the middle and upper part of the Spring Hill Limestone Member of the Plattsburg Limestone. Inasmuch as detailed information is readily available in the cited papers by Harbaugh and Wray, it is sufficient here to note that lettucelike (or according to Wray, potato-chip-like) fronds of algal thallus are abundant, some packed together in loose layers and some irregularly distributed, but nearly all with their surfaces roughly parallel to bedding. Organisms associated with the algae include scattered brachiopods, bryozoans, crinoid remains, and a few mollusks.
Wray (1964, p. 12) has briefly summarized paleoecological aspects of the Archaeolithophyllum assemblage as follows: "(1) Species [of the algae] developed as either encrusting, locally attached, or free forms. (2) Thalli were semirigid crusts capable of providing a self-supporting skeletal framework and a sediment-binding function in the depositional environment. ( 3) Thalli are believed to have grown on both carbonate mud and coarser grained sediment substrata. (4) Based on lithologic evidence and on analogy with Recent coralline algae, Archaeolithophyllum most probably inhabited the inner sublittoral marine environment, being able to tolerate appreciable wave agitation, but it also extended to depths of about 100 feet."
Thin, irregularly crenulated leaflike sheets, which doubtless are algal thalli, though altered by recrystallization to such extent that internal structures have been obliterated, are common in various Pennsylvanian and Permian limestones (e.g., Plattsmouth Limestone containing a Beil-type Pulchratia assemblage, Leavenworth Limestone containing an Isogramma assemblage). Plainly, these algae do not in themselves indicate paleobiotopes comparable to that called Spring Hill-type with abundant Archaeolithophyllum, and likewise the presence of more or less common Osagia or Ottonosia algal-foraminiferal colonies does not alone serve to identify paleobiotopes represented by so-called Osagia-type and Ottonosia-type organic assemblages. Possibly such observations are unnecessary, but it is well to bear them in mind.
An early Permian ecologic assemblage which with little or no hesitation I class as Spring Hill (Archaeolithophyllum) type is very well displayed in the Cottonwood Limestone of Greenwood County, Kansas. It is represented by what others (Imbrie, 1955; Imbrie, Laporte, and Merriam, 1959; Laporte, 1962) have designated as Anchicodium facies or platy algal facies of the Cottonwood. In my opinion, the leaflike (phylloid) algal remains of the Cottonwood, abundant enough locally to form marine banks, are misidentified as Anchicodium, for Johnson's (1946, 1963) descriptions and illustrations of this genus emphasize the "cylindrical, long, slender, and ropelike" nature of the thallus and common branching of the cylinders as main attributes. Among the half-dozen species distinguished by Johnson a platelike growth is mentioned only in one, for concerning A. gracile (Stanton and Oread Limestones) he reported (Johnson, 1946, p. 1098) "one specimen seems to have thick crustlike base from which cylindrical thalli arise." Internal structure of the Cottonwood algae is too poorly preserved to allow satisfactory generic identification but on the basis of all observed features they belong, in my judgment, to Archaeolithophyllum, not at all to Anchicodium nor to the codiacean Eugonophyllum, with vertically growing blade-like thalli. If so, this strengthens classification of the Cottonwood "platy algal facies" with the Spring Hill-type (Archaeolithophyllum) facies.
Very numerous Pennsylvanian and Permian limestones in Kansas are available for selection to illustrate marine organic assemblages which consist predominantly or, so far as observable in the field, exclusively of Osagia colonies (Suppl. Fig. 8, 15). Typically, limestones containing abundant Osagia occur at or near the top of marine parts of Pennsylvanian and Permian cyclothems and accordingly they have been interpreted as marginal and nearshore deposits laid down by retreating shallow seas. So much is a sound inference based on stratigraphy. Whereas geographically equivalent parts of transgressing seas may have been somewhat subnormal in salinity owing to fresh water from the land being invaded, marginal waters of retreating seas, including those temporarily trapped in lagoons, are likely to have been hypersaline. Many modern algae are adapted to such an environment, as witnessed by algal deposits along shores of Great Salt Lake.
Upper layers of the Morrill Member of the Beattie Limestone, especially in southern Kansas and farther south in Oklahoma are so largely composed of Osagia that they are exceptionally good examples of limestone called "osagite." The whole mass of the rock consists of closely packed ellipsoidal, spheroidal, or irregularly tabular bodies which range in longest dimension from approximately 3 to 15 mm. Concentric laminated structure commonly is observable without resorting to examination with a hand lens, and this structure characterizes all of the bodies except for bits of foreign substance such as shell fragments and calcareous or siliceous grains at centers of growth. Specimens studied by Twenhofel (1919), who first described and named them, were nearly opaque limonitized objects which he originally thought might be inorganically formed ooliths and pisoliths but later, finding that they lacked radial structure, concluded were produced by iron-fixing algae. He introduced for them the generic name Osagia, with O. incrustata as type-species, based on specimens collected from the Foraker Limestone (approximately 175 feet below the Morrill) in southern Kansas. Osagia is now recognized as consisting mostly of intergrowths of Girvanella-like algae and various genera of ammodiscid or cornuspirid foraminifers (Johnson and Konishi, 1956; Henbest, 1963). The foraminiferal participants in the colonies are chiefly Hedraites, Apterrinella, Minylecytheca, and Minytripetes; according to Henbest (1963, p. 35) the genus identified as Nubecularia by Johnson (1946, 1947) is not the same as true Nubecularia, known only from Jurassic to Recent (Loeblich and Tappan, 1964, p. 444) and accordingly should be rejected as a supposed associate of Pennsylvanian and Permian algae of Osagia type.
Among reasons for attaching paleoecological importance to Osagia and similar algal-foraminiferal intergrowths is phylogenetic kinship of the algaloid colonies with chlorophyll plants. Their occurrence in association with marine invertebrates indicates an environment within the photic zone of Pennsylvanian-Permian shallow seas. The algal laminae of Osagia commonly surround completely the nuclei or supports on which they grow. Inasmuch as they and foraminifers intergrown with them must have had unimpeded access to open water, yet lacked any means for moving about or turning over, their supports had to be shifted and turned frequently by such agencies as waves and currents. Bottom-feeding animals may have contributed to moving the colonies during growth so as to build them somewhat evenly. It is reasonably certain that most specimens lived and developed where they became buried and thus they indicate ecological features pertaining to the algae-bearing beds. If, as suggested in the introductory paragraph on Osagia-type assemblages, these belong to late retreatal stages of cyclic marine invasions in which marginal parts of shallow seas tended to have excess salinity, the foraminiferal constituents of the Osagia colonies would have needed to be as tolerant of hypersaline environments as the algae. Gypsum deposits several feet thick were laid down in northern Kansas at the close of the Bader cycle, which next followed the Beattie cycle when Morrill osagites accumulated shortly before the Beattie sea vanished.
Last of the Pennsylvanian-Permian ecologic assemblages and biotopes chosen for consideration in this paper are those which I find most puzzling of all. They are designated as Heebner-type because the Heebner Shale Member of the Oread Limestone is selected as a very representative example. Excellent outcrops of this member on the Kansas Turnpike, 3 miles west of the West Lawrence Interchange (Fig. 13) are cited for reference. Here, as everywhere else, the black platy Heebner shale deposits rest with knife-sharp contact on the Leavenworth Limestone, previously discussed. We are primarily concerned with the lower half of the Heebner, all of which consists of very evenly laminated layers that generally weather in papery flakes or thin sheets resembling tough black cardboard. The total thickness of such deposits ranges from a few inches to approximately 3 feet, rarely more than this anywhere. In horizontal distribution these shales are even more extensive than the remarkable persistent limestone next beneath them. A character which makes them readily identifiable in well borings is high radioactive content, shown by a strong "kick" in gamma-ray well logs (Fig. 27).
Fossils are reasonably common but extremely inconspicuous in Heebner-type assemblages. They consist of conodonts, among which the following genera (some represented by several species) have been identified (Ellison, 1941, 1946, 1947) (Suppl. Fig. 8, 1-12) .
| Conodonts of Heebner-type Assemblages | |
|---|---|
| Cavusgnathus | Metalonchodina |
| Euprioniodina | Ozarkodina |
| Gondolella | Prioniodina |
| Hibbardella | Prioniodus |
| Hindeodella | Spathognathodus |
| Idiognathodus | Streptognathodus |
| Ligonodina | Synprioniodina |
| Lonchodina | Trichognathodus |
| Lonchodus | |
Fairly common at some localities and seemingly characteristic of the Heebner-type assemblage are small longitudinally striated fish spines (Listracanthus), which occur as very thin impressions with delicate ray extensions along one margin and at the tip, giving a frayed appearance to these edges (Suppl. Fig. 8, 13). Extremely thin-shelled, fragile specimens of Aviculopecten, a marine bivalve, have been collected from some outcrops. The environment indicated by these organic remains and by physical characters of the shale is very problematical. Assuredly the setting was not a normal open-sea with shallow bottom and with equal assurance one may conclude that the black shale is not a deep-water deposit (that is, in terms of the Pennsylvanian seas which repeatedly spread over the Kansas stable platform, with estimated maximum depths of less than 100 m.). A reason for inferring that depths of only a few meters existed even at great distances from shore when marine waters extended most widely is the necessity of eXplaining fluctuations of water depth either by warping of the sea bottom or by changing the sea level. Inasmuch as emergent conditions in large areas are indicated by deposits located a few feet below and a few feet above the marine strata, it is unreasonable to postulate up-and-down movements of the earth crust or of sea level measured in tens of meters. Rather, the stratigraphic record indicates that sedimentation and subsidence were almost in equilibrium throughout the Kansas region during Pennsylvanian and Permian time, and it follows that nowhere at any time were seaways describable as other than very shallow, or in terms of modern coastal shelves as extremely shallow.
In connection with paleoecological study of Heebner-type black shales in Kansas and adjoining states, attention should be called to an extraordinarily thorough and painstaking investigation by Zangerl and Richardson (1963) of lithologically similar black platy shales and associated beds of Pennsylvanian age in western Indiana. Detailed stratigraphic field studies and extensive collecting of fossils, mainly at two sites, were supplemented by investigations of modern black mud accumulations in the Mississippi delta region. Then a "sample" of the Pennsylvanian shale from a selected quarry was taken out and shipped en bloc forbe laid out on the floor exactly as observed when the shale was in place. Dimensions of the collected part of the beds are reported as approximately 12 by 15 feet (180 sq. ft.) in area and 9 inches in thickness. Investigation proceeded by concentrating attention first on the uppermost 0.25 inch, analyzing all of its physical and chemical characters and mapping the occurrence of all observable kinds of organic remains. Ultimately, 35 additional divisions, each approximately 0.25-inch thick, were similarly studied. The density of fossil materials and their variety showed considerable difference from layer to layer; one of the thin units displayed 68,000 specimens on its surface, not all of which could be charted. No remotely comparable study of even a small block of Heebner-type black shale in the Kansas region has been made, but it is safe to predict that results of such an investigation would be extremely dissimilar from those described by Zangerl and Richardson, especially in the paleontological features discovered. Information given here on their observations is condensed into mere listing of the kinds of fossil remains discovered, supplemented by brief statement of a few of their conclusions.
| Organic Remains Reported by Zangerl and Richardson (1963) from Pennsylvanian Black Platy Shale in Western Indiana | |
|---|---|
| Plants | |
| Terrestrial ferns (e. g., Neuropteris, Sphenopteris, Pecopteris), rare, undoubtedly transported from adjacent land | |
| Driftwood, indeterminate, locally abundant | |
| Plant debris, including supposed seaweeds, fusain, abundant degraded products, few spores | |
| Invertebrates | |
| Sponges: small patches of spicules | |
| Corals: *Lophophyllidium, locally common | |
| Bryozoans: *Fenestrate zoaria, several species common; rare ctenostomes | |
| Brachiopods: Lingula, orbiculoids, *chonetids (Mesolobus), *productids (Desmoinesia, Antiquatonia), *spiriferoids (Neospirifer, Composita), rhynchonelloids (Wellerella) rare | |
| Gastropods: chiefly varied small pseudozygopleurids, juvenile bellerophontids, Phanerotrema | |
| Cephalopods: straight (Pseudorthoceras) and coiled nautiloids, goniatites (Pronorites, Paralegoceras), moderately common locally | |
| Pelecypods: myalinids (Myalinella), locally common, pectinids (Dunbarella, locally very abundant, Euchondria, Streblochondria), pteriids (Pteria, locally very common), nuculids (Nucula, rare), cardiomorphid (Edmondia, rare), pholadellid (Allorisma, rare) | |
| Worms: polychaetes (Microconchus, fairly common), oligochaete, rare | |
| Trilobites: *"Brachymetopus," single specimen | |
| Crustaceans: ostracodes (several species, fairly common), phyllocarids (Concavicaris, locally abundant), percarids (common) | |
| Echinoderms: crinoid columnals and plates (rare), echinoid spine (Archaeocidaris, rare) | |
| Conodonts: several kinds (esp. Hindeodella) common | |
| Vertebrates | |
| Fishes: acanthodian sharks (Acanthodes?, common; "placoderms," rare), elasmobranchs (Petrodus, Listracanthus, at least 10 more genera, fairly abundant), palaeoniscids (locally abundant); crossopterygians (fairly common) | |
| *Fossils mostly confined to lenticular masses of black mudstone locally interbedded in the shale are marked by an asterisk. | |
This is an amazing list, even in the summary form here presented, and the possibility of reporting it is entirely dependent on the exhausitive researches of Zangerl and Richardson. Paleoecologists generally and Pennsylvanian stratigraphers and paleontologists especially owe a large debt to them. A few of their conclusions warrant statement in the present paper. (1) The black platy shale studied in Indiana accumulated at the rate of approximately 3 inches in a year (if true, very surprising). (2) Compaction of the shale was synchronous with deposition, air-filled cavities in spongy bone remaining uncrushed. (3) The black shale comprises shoal-water deposits of slightly variable depth in nearshore environments during initial phases of a marine transgression. (4) Concentration of autochthonous burial communities was effected in residual ponds on the coastal plain during dry seasons. (5) Owing to widespread growth of a dense floating mat of vegetation (flotant), water beneath the mat was quite undisturbed by waves and currents and the bottom was extremly quiet. (6) Whereas water above the bottom was inhabitable, the bottom itself was toxic, lacking scavengers and having neither an infauna or epifauna of benthonic invertebrates. (7) The fauna invaded the burial site from its natural habitat farther seaward and individuals died mostly from effects of predation, all vertebrates discovered showing marks of bite injury and study in a laboratory at the Chicago Natural History Museum where it could many invertebrates being found in gastric residues and coprolites.
Work on samples of one or more Kansas black platy shales, collected so as to retain all features intact and selected to consist of only a few cubic feet, would undoubtedly add a good deal to knowledge if the intensiveness of study even halfway matched that given to the Indiana sample, yet in my opinion the examination of the Kansas black shale would be monotonous at least in organisms discovered and paleontological results would not call for lengthy description. The Kansas shales are too much of a single type, each closely resembling others, and none showing indication of numerous and varied kinds of organisms. Even so, we need to learn more about them if headway in dealing with paleoecological interpretation is to be made. Whatever the findings, these should relate in essentially uniform manner to sea-covered areas of 40,000 to 50,000 square miles in the Kansas region (Fig. 27, 28), instead of the small tract, say 500 square miles (estimated 5-mile wide zone along margin of basin multiplied by arbitrary 100 miles of shore line; Zangerl and Richardson, 1963, p. 214), which the Indiana black-shale sample was used to interpret paleoecologically.
It seems to me worth while to examine at least part of the observations made by Imbrie (1955) and his protege Laporte (1962) on the characters of ecologic assemblages and corresponding biotopes based on unusually exhaustive studies of organisms and sediments collected from numerous places along the outcrop of the Beattie cyclothem in Kansas (Imbrie, Laporte, and Merriam, 1959, p. 69; called Cottonwood cyclothem by Merriam, 1963, p. 105). Most of the very widespread marine deposits of this cyclothem are contained in the three members (Cottonwood Limestone, FIorena Shale, Morrill Limestone) of the Beattie Limestone, but some of them occur in the Eskridge Shale and Stearns Shale which mainly are nonmarine (Fig. 33).
Figure 33--Diagrammatic section of the Beattie cyclothem (after Merriam, 1963).
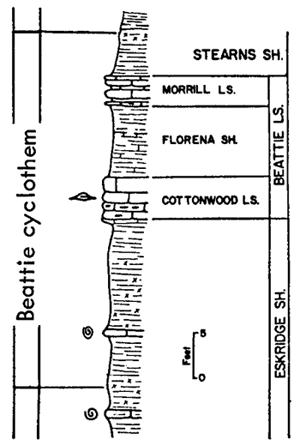
Both qualitative and quantitative investigations of Beattie units made by Imbrie and Laporte have been very detailed, although most of Imbrie's work has not yet been published. Dependence by them on methods making large use of computers is of interest. It is plain, however, that objectivity of results and reliability of conclusions are not at all guaranteed by investigative techniques alone. Data processing by means of computers certainly involves subjective factors in programming, and human brains still are needed for tasks of synthesis and interpretation. Effort to fit paleoecological conclusions into acceptable representation of paleogeography corresponding to them puts heaviest of all demands on control of geological imagination. Some aspects of these matters I propose to discuss.
To begin, reference is made to a diagrammatic representation of Beattie Limestone facies observed along the outcrop from southern Nebraska to northern Oklahoma as interpreted by Imbrie (Imbrie, Laporte, and Merriam, 1959) (Fig. 34A) and to a very similar diagram drawn at larger scale showing facies of the Cottonwood Limestone Member of the Beattie (Laporte, 1962) (Fig. 34B, C). These depict several very dissimilar types of deposits which can be distinguished by both physical and paleontological characters. Each marks a certain kind of paleobiotope which needs to be studied in all possible ways for the purpose of interpreting it accurately as a unit and then interrelationships of each with others found bordering it on all sides must be examined in order to identify changing environmental patterns in their correct regional and temporal settings. Problems are numerous and some of them difficult, but when most are reasonably solved, geologists can undertake to describe the nature and distribution of Beattie sedimentary environments and with appropriate qualifications to translate them into paleogeography of the Kansas region in Beattie time. The studies by Imbrie and associates have been carried far enough to justify in their opinion not only expression of the extent of the Beattie sea, with northwesterly trending strand lines on northeast and southwest sides of a 400-mile-wide strait, but also indication of the topography of the shallow sea bottom (Fig. 35). Thus we are introduced to a so-called "Arkansas Embayment," a "Greenwood Shoal," a "Kansas Shelf Lagoon," and other environmental provinces or subprovinces. I shall return to these imagined features later.
Figure 34--Facies of Beattie Limestone differentiated by studies along outcrop from Oklahoma to Nebraska. A, Diagrammatic section of entire Beattie (after Imbrie, Laporte, and Merriam, 1959). B, C, Diagrammatic section of Cottonwood as determined from sampled exposures (after Laporte, 1962). [Included in the Acrobat PDF file of figures for this article.]
Figure 35--Paleogeography of Beattie time and diagrammatic north-south profile across "Kansas Strait" as interpreted by Imbrie, Laporte, and Merriam (1959).
One should take for granted that firm determination of the stratigraphic framework of any given paleoecologic investigation is indispensable, because misidentification of sedimentary units, including regional or local miscorrelation of them, inevitably leads to faulty conclusions and possibly to completely erroneous ones. Rock samples and fossil collections from incorrectly recorded stratigraphic sources are worse than valueless since work done on them not only is wasted but it serves to provide built-in errors.
Fortunately, most parts of the Pennsylvanian and Permian rock succession in Kansas have been differentiated, defined, and traced along outcrops (to some extent also beneath the surface) in such detail and with such precision that a highly reliable stratigraphic framework is available for almost any chosen paleoecologic study. By and large, this reflects the evenly constructed layer-cake arrangement of deposits which were spread out over the Kansas stable-platform area in late Paleozoic time. Strata of the Beattie Limestone conform to the prevailing regularity of pattern and particularly throughout central and northern Kansas and southern Nebraska the three members are easily identified without question. In addition, the Cottonwood is divisible everywhere in this area into a lower "bioclastic" half lacking fusulinids and an upper fusulinid-packed half (Imbrie, Laporte, and Merriam, 1959; Laporte, 1962) (Fig. 34). Southward from Greenwood County, Kansas, the stratigraphy of the Beattie Limestone is not so simple, though it has been supposed to present no problems.
Imbrie (1955) published the locations of four southern Kansas sections of the FIorena Shale from which he collected 11-pound (dry weight) samples at 1-foot intervals but described none of them. Later, two Osage County, Oklahoma, sections and two Cowley County, Kansas, sections measured, described, and drawn up graphically by Imbrie (1959, p. 45-47, 51; though without indicated authorship), they purport to show lithologic, paleontologic, and thickness characters of Beattie Limestone divisions. Also given are Beattie sections in Greenwood, Chase, and Morris Counties, Kansas (Imbrie, 1959, p. 56, 61, 64). Laporte (1962) utilized virtually all of Imbrie's Nebraska-to-Oklahoma sections and seemingly additional ones of his own in a detailed paleoecological study of the Cottonwood Limestone, but the stratigraphic content of his observations must be gleaned from a single restored regional cross section intended to show areal distribution of facies in the Cottonwood (Fig. 34B,C). A similar regional diagram of the entire Beattie, previously cited (Fig. 34A), shows that all parts of the formation change markedly in facies from Greenwood County southward and eventually almost pinch out.
I have long been interested in Imbrie's paleoecological research on Beattie deposits, being responsible, indeed, for initiation of his work (Imbrie, 1955, p. 649). Also, I have measured and described in field notebooks many Beattie sections and I have made large collections of fossils at many Beattie outcrops. The background of such acquaintance facilitates understanding and acceptance of the various facies which have been described by Imbrie and Laporte. Their indicated distribution and thickness of these in southern Kansas and northern Oklahoma, however, has seemed to me open to question. For example, the upper Cottonwood "fusuline facies," persistent and well developed in the north, is indicated by them to disappear near the Chase-Greenwood County line, whereas I have recognized that Beattie fusulinids persist across Greenwood and Elk Counties into Cowley County and Osage County, Oklahoma, some 20 miles south of the Kansas state line. Actually, Imbrie (1959, p. 46, 47, 51, 61) has recorded correctly the fusulinid zone of the Cottonwood in three Greenwood and Cowley County sections and one in Osage County, Oklahoma, but in another section farther south he miscorrelated it with fusulinid-bearing beds just beneath the Morrill Limestone (Imbrie, 1959, p. 45). Laporte (1962, p. 530) also recorded the presence of a few inches of fusulinid-rich rock "which occurs at the top of the Cottonwood from central Kansas to northern Oklahoma" but because of its thinness he did not consider it to be a continuation of the "fusuline facies" of the Cottonwood (Fig. 34B, C). Seemingly, statistical analysis of facies attributes minimizes quantitatively small, though possibly very significant stratigraphic elements to the point of obliterating them from notice. Thus, in successive tracts the entire Cottonwood is classed as belonging to a platy algal facies, a "shelly facies," or a silty Osagia facies (Fig. 34C). It is evident that stratigraphic definition of Cottonwood in this region constitutes a prerequisite for facies analysis of the unit and if this definition is untrustworthy or erroneous, the facies analysis is likewise untrustworthy or erroneous. The first problem to settle is what strata are correctly classifiable as Cottonwood, FIorena, and Morrill.
My decision to review the work of Imbrie and Laporte on Beattie cyclothem ecologic assemblages and biotopes for purposes of the present paper initially was aimed at considering some of their general conclusions, such as determination that maximum water depths in the Beattie sea are recorded by "shelly facies" and "Chonetes facies" deposits, rather than by "fusuline facies" sediments, and decision that Beattie deposition was essentially unrelated to distance from a postulated geographically stable, nonfluctuating shore line (Imbrie, Laporte, and Merriam, 1959, p. 78; Laporte, 1962, p. 540). Attempts to reconstruct the environmental settings of each distinguished Beattie facies, based on interpretation of all discoverable physical and biologic characters in each, are laudable even if highly speculative, as is most likely when any given facies is studied independently. Efforts along this line are best guided in right directions by focusing attention on resemblances and differences of contiguous facies and on stratigraphic relationships of each to others. An accurate stratigraphic framework is an indispensable starting point.
As already stated, from central Kansas northward, Beattie stratigraphy is clearly defined and it provides a straightforward approach for investigation of ecosystems and biotopes belonging to the formation. In southern Kansas and northern Oklahoma conditions are sufficiently otherwise to call for field re-examination, and accordingly, aided by D.F. Merriam and T.E. Jacques, of the Kansas Geological Survey, I devoted some days (13-16 June, 12-13 July, 1964) to renewed study of Beattie outcrops in the southern Kansas and adjacent Oklahoma area. Stratigraphic observations made, including remeasurements of some sections for more accurate determination of thicknesses and placement of various thin beds, are shown graphically in Figure 36, but as explained subsequently, these have been modified from their original form with respect to delineation of the Cottonwood and indication of fusulinid occurrences in the Florena. Because my review had shown that information published by Imbrie and Laporte on Beattie deposits south of Greenwood County was inadequate and partly open to question, these geologists were invited to come back to Kansas for the purpose of examining field evidence and discussing its significance. They consented readily, with result that on 5-7 August 1964 a trip to the area was made by Imbrie, Laporte, Merriam, Jacques, and me. This led to agreements which are incorporated in the graphic sections (Fig. 36) now presented. Additionally, through Imbrie's kindness, previously unpublished records of Florena fusulinids are given; with minor exceptions, for me this was new information.
Problems of paleoecologic and paleogeographic interpretation of Beattie deposits are encountered in observing contrasted features of stratigraphic development along the outcrop (1) in northern Kansas and southern Nebraska, (2) in the Greenwood County area of central Kansas, and (3) in southern Kansas and northern Oklahoma. From the southern boundary of Greenwood County northward, members of the Beattie are clearly delineated and the same is true for the Morrill and most of the FIorena southward, but definition of the Cottonwood Limestone becomes difficult owing to change of facies and lithologic-paleontologic similarities with lower Florena and upper Eskridge strata. Therefore, some description of the rock succession in the Beattie part of the column in southern Kansas and northern Oklahoma is needed.
In west-central Greenwood County (Fig. 36, Loc. 12) divisions of the Beattie are clearly defined, the Morrill consisting of massive osagite limestone with numerous fairly large algal "biscuits," the Florena being mostly soft shale with Neochonetes, Meekopora and a few other fossils sparsely distributed in the upper part and abundant fusulinids at the base, and the Cottonwood comprising a prominent scarp-making limestone with abundant fusulinids in the topmost few inches and the remainder predominantly built by platy algae. A sharp boundary separates the Cottonwood from underlying poorly fossiliferous (pectinoid clams, fenestrate bryozoans), tan-weathering Eskridge Shale.
Members of the Beattie are also differentiated unequivocally in southwestern Greenwood County (Fig. 36, Loc. 11), although here irregularly bedded fusulinid-bearing rock, 2 feet thick, occurs at the base of the Morrill. The Florena consists mainly of shale but contains thin discontinuous limestone layers; fossils are moderately abundant, with Neochonetes predominant in the upper half and fusulinids near the base. The Cottonwood is mainly a platy algal limestone, but the upper 1 foot lacks algae and contains common fusulinids at the top. The upper Eskridge, with knife-sharp separation from the Cottonwood, contains abundant marine fossils (chiefly bivalves and brachiopods).
Figure 36--Correlated sections of Beattie and upper Eskridge strata in southgern Kansas and northern Oklahoma based on Moore's 1964 field work, supplemented by observations made during field conferences with Imbrie, Laporte, Merriam, and Jacques in August, 1964, and data on Florena fusulinids furnished by Imbrie. [Included in the Acrobat PDF file of figures for this article.]
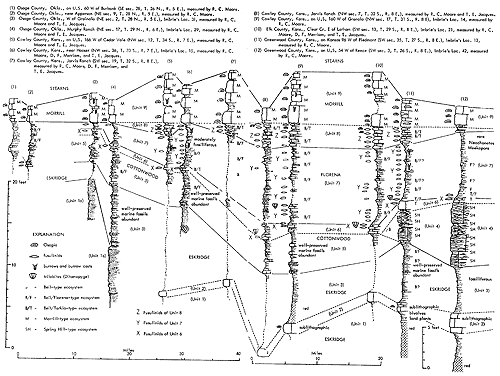
Ten miles farther south, in northwestern Elk County (Fig. 37, Loc. 10), the Morrill and Florena resemble counterparts in the last-described section but are thicker. Imbrie (Fig. 34A) here recorded 12 feet of Florena beds (instead of 9 feet measured by me), the lower half being classed as "shelly facies," underlain by 5 feet of "shelly" Cottonwood. The platy algal facies of the Cottonwood is gone. At this horizon I found fusulinid-bearing and bioclastic limestone 1 foot thick resting with sharp boundary on shale containing some marine fossils, only 1.5 feet approximately being exposed. Unhappily, the Elk County section was not visited during the joint conference of early August and therefore questions concerning members of the Beattie at the locality are unsettled.
The Cowley County sections at Localities 9, 8, 6, and 5 (Fig. 36), in southward order, were examined by the members of our conference group and possibly as a result of working under the pressure of unusually hot weather (temperatures to 111° F.), agreement concerning definition of Beattie members was reached. The same can be said of sections in Osage County, Oklahoma (Fig. 36, Loc. 3, 4). It is unnecessary to describe these separately but desirable to summarize some features of the stratigraphic succession in the area as now understood. For this purpose, various units independent of stratigraphic classification are discriminated and numbered in upward order. They are described briefly in following paragraphs.
Unit 1 (Eskridge)--Lowest strata shown in sections plotted in Figure 36 consist of unfossiliferous gray and red shale (Loc. 12) , brownish shale containing land-plant remains and marine bivalves underlain by red shale (Loc. 11), and shale of undetermined nature, mostly covered (Loc. 5, 7-10).
Unit 1a (Eskridge)--Fine-grained crossbedded sandstone, 10 to 12 feet or more thick, occurs at the base of some Osage County sections (Loc. 3, 4). The unit contains no observed fossils and its relation to Unit 1 is unknown.
Unit 2 (Eskridge)--A distinctive, seemingly persistent limestone which is easily found at outcrops is a useful marker designated as Unit 2. It is a distinctly blue, light blue-gray-weathering sublithographic limestone which is hard, massive, and generally devoid of fossils other than a few small Osagia pellets locally. No other stratum comparable to it is found higher in the section at least to the top of the Morrill Limestone. In some exposures the bed displays a faintly mottled appearance and when broken with a hammer, conchoidally shaped fragments reveal rounded structures resembling pebbles, but not true pebbles. The bed commonly is little more than 1 foot thick, but at Locality 12 (Fig. 36) it is 2.3 feet in thickness and appears cross laminated. At Locality 11, the top 0.2 foot contains Osagia and abundant shells of Aviculopecten associated with a few other clams and the basal 0.3 foot and subjacent shale also bear numerous clams.
Unit 3 (Eskridge)--A lithologically and paleontologically variable division next above Unit 2 or Unit 1a is differentiated as Unit 3. At Localities 4 and 11 shale alternating with thin, partly nodular and discontinuous limestone layers contain numerous to abundant well-preserved marine fossils, among which Schizodus, Aviculopecten, Clavicosta, Pseudomonotis, and Septimyalina are most common bivalves and Juresania, Derbyia, Neochonetes, Composita, and Meekella predominant brachiopods; associated with these shells are gastropods (Amphiscapha, ?Warthia) and bryozoans (ramose and fenestrate forms). The upper boundary of the unit is sharply marked at Locality 11 by the base of massive Cottonwood Limestone, but at Locality 4 hardly any difference between Unit 3 and overlying fossiliferous shale and limestone placed in Unit 5 can be discerned. At this place the basal bed of Unit 3, resting on Unit 1a, is a thin limestone conglomerate containing pebbles of sublithographic limestone up to 1 inch or more in diameter. At Locality 12 the unit consists entirely of shale which bears a few ill-preserved marine fossils (bivalves, bryozoans); the lower and upper boundaries are sharp. Elsewhere Unit 3 consists of gray and red shale without observed fossils.
Unit 4 (Cottonwood)--Massive light-gray limestone attaining a maximum observed thickness of 8 feet (Fig. 36, Loc. 12) is differentiated as Unit 4. The rock is crowded with the predominantly horizontal thin leaves of platy algae, which in my opinion belong to Archaeolithophyllum rather than Anchicodium, as recorded by Imbrie, Laporte, and Merriam (1959) and Laporte (1962). The unit is not known to extend southward beyond the limits of Greenwood County, Kansas.
Unit 5 (Cottonwood)--Discontinuous, uneven, partly nodular thin blue-gray limestones interbedded with gray, tan-weathering calcareous shale characterize Unit 5. Well-preserved invertebrates of varied kinds are numerous in all exposures and very abundant in some. This division, 5 feet thick at Locality 9 (Fig. 36) and 3 or 4 feet in sections farther south, constitutes most of the "shelly" Cottonwood of Imbrie and Laporte, who include under this term the overlying fusulinid-bearing limestone differentiated here as Unit 6. At Locality 4 (Fig. 36) the lower boundary of Unit 5 is so ill-defined that separation from the lithologically similar and almost equally fossiliferous upper Eskridge (Unit 3) is almost arbitrary, but elsewhere difficulty in defining the boundary is not encountered. Because the fossiliferous beds of Unit 3 at Locality 11, unquestionably assigned to the Eskridge Shale, rather closely resemble Unit 5, I was inclined, before the August field conference with Imbrie and Laporte, to consider them as equivalent to one another, restricting the Cottonwood to Unit 6. Now I agree that Unit 5, which contains more limestone than Unit 3 at Localities 4 and 11, is classifiable as a facies of the Cottonwood very unlike the normal expression of this member and rather closely similar to Florena and locally developed facies of the upper Eskridge in the southern Kansas and northern Oklahoma region. Fossils found in Unit 5 include the following (most common and widely distributed forms marked by an asterisk): *Neochonetes, *Reticulatia, *Juresania, *Composita, *Derbyia, *Meekella, Pulchratia, Crurithyris, Dielasma, Isogramma, *Aviculopecten, *Clavicosta, *Pseudomonotis, *Schizodus, *Allorisma, *Septimyalina, Aviculopinna, *Septopora, Fenestrellina, Polypora, Tabulipora, Rhombopora, Bellerophon, Omphalotrochus, high-spired gastropods, crinoid plates and columnals, echinoid spines and plates, small ophiuroids, Metacoceras. Possibly overlooked as components of this varied assemblage are fusulinids; at any rate it is noteworthy that they have not been found.
Unit 6 (Cottonwood)--Light bluish-gray, tan-weathering, mostly unevenly bedded limestone which everywhere contains common to abundant fusulinids is differentiated as Unit 6. The division is thin, generally less than 1 foot in thickness, but very persistent. It forms the topmost part of the Cottonwood, overlying platy algal limestone, at Localities 11 and 12 (Fig. 36) and similarly comprises the uppermost Cottonwood, overlying "shelly" beds of Unit 5 elsewhere south of Greenwood County. This fusulinid zone, marked "X" on Figure 36, is characterized by Schwagerina jewetti, S. emaciata, and Schubertella kingi. The species of Schwagerina may occur at higher horizons, but the diminutive S. kingi seems to be confined to Unit 6. Associated with the fusulinids are a few brachiopods and search at most outcrops is rewarded by finding numerous remains of trilobites. They consist of Ditomopyge pygidia (most common), parts of cephalons or whole cephalons, and in some places very perfect whole enrolled carapaces. The fusulinids and trilobites are an excellent marker faunal assemblage, observed by D. F. Merriam at the top of the Cottonwood in central and northern Kansas, as well as in the south.
Unit 7 (Florena)--Gray to tan, generally calcareous shale interbedded with thin nodular limestone layers characterizes Unit 7, which is 7 to 9 feet thick at Localities 8-12 but only 1 to 5 feet thick at Localities 3-6 (Fig. 36). In most places fossils, including well-preserved brachiopods and bivalves, are numerous. Fusulinids (Schwagerina jewetti, S. vervillei) were found by me in a thin limestone 2 feet above the base at Locality 9 and in topmost soft shale at Localities 3-6. Actually, fusulinids are much more widely distributed than this in Unit 7, although I did not know this until Imbrie kindly furnished information derived from his study of washed residues from systematically collected bulk samples of the Florena. The occurrences are marked by "Y" on Figure 36; the uppermost fusulinids of Unit 7 at Locality 6 were reported by Imbrie but curiously were not recorded at Localities 3 and 4 where I obtained numerous specimens. It is interesting to learn that the "Y" fusulinids are found from bottom to top of the Florena at Localities 6, 9, and 11, and with small gaps at Locality 10. A distinctive feature of Unit 7 is the common occurrence of burrow casts approximately 1 inch in diameter and several inches in length, some packed with small fossils. They are composed of tightly cemented material and thus weather out readily from inclosing shaly beds. Laporte has recorded their presence also in the "shelly" Cottonwood and Imbrie has found them widely distributed in the Morrill Limestone. Thus, the burrow casts are not a diagnostic feature of the southern Florena deposits.
Unit 8 (Morrill)--Fusulinid-bearing, unevenly bedded gray limestone which commonly weathers to splotchy tan-brown and gray characterizes Unit 8, ranging in thickness from a featheredge to nearly 3 feet. The limestone generally is harder and its layers slightly thicker than beds in Unit 6 of southern Kansas, and hence it is more prominent along the outcrop. A characteristic feature is the almost universal presence of moderately thick, light-gray to nearly white colonies of Osagia, especially in upper beds of the unit. The Osagia specimens very commonly surround fusulinids but also grow on shell fragments (Suppl. Fig. 8, 15). The fusulinids are prevailingly larger than those of Unit 6 and although many of them are similar in shape, some are distinctly more swollen medially. Nearly all specimens which have been studied in section are identified as Schwagerina vervillei, though small forms considered to be immature individuals of this species may belong to S. jewetti or S. emaciata. No Schubertella has been found in numerous samples of the Morrill fusulinid zone studied in thin section.
At Localities 5 and 7 to 10 (Fig. 36, 37), Unit 6 is directly overlain by thickbedded limestone with abundant Osagia, which here is defined as Unit 9, the upper division of the Morrill Limestone. At Locality 12 limestone lithologically extremely similar to the mottled gray and brown uneven beds of Unit 8 occurs next above Florena Shale containing Neochonetes and other sparse fossils, but no fusulinids were found in this limestone. Hence, it is doubtfully classed as belonging to Unit 8.
Unit 9 (Morrill)--The uppermost beds of the Morrill Limestone Member of the Beattie, 2 to 8 feet thick in most exposures, are distinguished as Unit 9. The layers are characterized by their massive, rough-pitted surfaces, and an abundance of small to moderately large Osagia colonies. Burrows and burrow casts approximately 1 inch in diameter and several inches in length are common in some localities but not found everywhere. Invertebrate fossils, consisting chiefly of such clams as Allorisma and Septimyalina, are rare. Unit 9 is a distinctive, readily traced part of the Beattie in southern Kansas and northern Oklahoma. The outcrop at Locality 1 (Fig. 36) is typical but no fossiliferous beds beneath the Morrill are found. The section at Locality 2, measured several years ago, is doubtful because it differs materially from sections farther north and because the exposure could not be relocated in the course of recent field work.
A qualitative and quantitative view of the assemblages of fossil organisms found in sections of the FIorena Shale distributed along the outcrop across Kansas has been made possible by John Imbrie in providing me with a partial summary of studies made by him and in generously allowing me to shape data which I have selected into graphic form as illustrations (Fig. 37, 38) in this paper. Details and much supplementary information will be published by Imbrie. In my opinion, the diagrams speak for themselves in showing clearly the diversity in composition and distribution of faunal assemblages within the FIorena, including several relationships which to me, at least, were quite unexpected. For example, field observations have led me to judge that Neochonetes is an overwhelming constituent of the Florena fauna generally and especially that representatives of this genus predominate in lower parts of the member. Imbrie's data serve more to refute than to support such judgment.
Figure 37--Biofacies of Florena Shale (composite by localities). [Localities and explanation of letters and patterns as in Figure 38.]
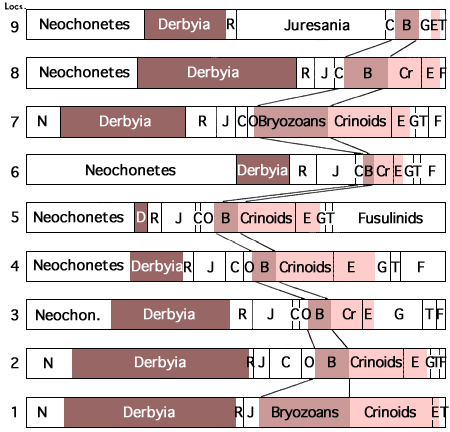
Not available at this time.
I refrain trom any effort to explain the composition of Florena fossil assemblages at different levels and from place to place, for under the circumstances this would be highly inappropriate. Obviously, measurements of discriminated faunal components in terms of skeletal weight furnish only a rough indication of the makeup of fossil assemblages in terms of total numbers of individual specimens classified as belonging to designated taxa. The census of a given local population of Florena invertebrates might be approximated from Imbrie's data if we knew that the weight of one clean empty shell of adult Reticulatia matched four Derbyia shells, 18 Neochonetes shells, 20 Ditomopyge carapaces, 24 Schwagerina tests, and so on. Lack of information of this sort and inability to take account of juvenile individuals lead us to be content with weight fractions of skeletal residues as measure of the competitive withdrawal of calcium carbonate from sea water by the differentiated groups of organisms.
Leaving paleoecological interpretation of the Florena data to Imbrie, I merely call attention to dominance of brachiopods throughout most of the sediments (best shown in Fig. 37), at the same time noting the diversities in proportions of different genera shown by the diagrams. Although some Pennsylvanian and Permian ecosystems seem to be measurably uniform throughout very large areas, the Florena diagrams indicate that the nature and distribution of the biota are complexly varied--so much so as to suggest that the designation Neochonetes-Derbyia assemblage is an oversimplification. Also, I readily agree with Imbrie, Laporte, and Merriam in concluding that the paleoecology of Beattie deposits must be interpreted in terms of more factors than depth of seas, distance from nearest shore lines, and progressive changes at selected stations incident to transgression and recession of shallow marine waters.
My purpose in discussing the paleogeography of Beattie time is to stress two or three general points which in my judgment have importance. It is not to translate the classification of sedimentary facies displayed by outcropping beds into plotted locations of Cottonwood, Florena, and Morrill strand lines or to outline within the area of the inferred Beattie sea such features as shoals, shelf lagoons, shelves, and embayments. Imbrie, Laporte, and Merriam (1959, p. 71) undertook to do these things (Fig. 35), assuming that their map was good for essentially the entire span of Beattie time, whatever its duration may have been, for they wrote (p. 78) "during Beattie deposition shoreline positions . . . did not fluctuate significantly" and "it is clear that no major advances and retreats of the shoreline could have occurred during Beattie time."
The first point worthy of consideration, I think, is the utter inadequacy of basic data available for paleogeographical interpretation. Suppose we assume that paleoecological interpretations are penetrating and trustworthy. Then it is important to remember that the source observations come almost exclusively from a few dozen exposures along a narrow outcrop belt which ends northward and probably also southward far short of Beattie shore lines. It is true that subsurface information is available, but this consists of little more than determination of the presence and thickness of some rock units, providing data of small value for paleoecological and paleogeographical studies. Naturally, quite unknowable are the nature and areal extent of Beattie sediments destroyed by post-Permian erosion. That they originally were spread over many hundreds and probably some thousands of square miles east of present-day outcrops is undeniable, but only so much can be inferred safely. In my opinion, extrapolations of the sort given by Imbrie, Laporte, and Merriam (1959, p. 74) showing thickness of the Florena Shale throughout eastern Kansas counties in which hypothetically once-present deposits have been obliterated are both baseless and possibly quite misleading even if one recognizes the element of pure guesswork (Fig. 39). For example, the depicted tract of southeastwardly thickening Florena deposits is an extrapolation from a single very small area of exposure in northeastern Cowley County, Kansas, in which the plotted thickness of the Florena is too great (12 feet instead of actual 8.5 to 9 feet). This, then, is a faulty, as well as extremely tenuous, basis for the mapped east-of-outcrop thick Florena, though it accords possibly with the unexplained and unsupported paleogeographic concept of a so-called "Arkansas Embayment." "Waters of the Kansas Strait communicated with the open ocean principally to the southeast by way of an arm of the sea here called the Arkansas Embayment" (Imbrie, Laporte, and Merriam, 1959, p. 70).
Figure 39--Thickness of Florena Shale (feet) in eastern Kansas showing extrapolated isopach contours east of the outcrop (simplified from Imbrie, Laporte, and Merriam, 1959).
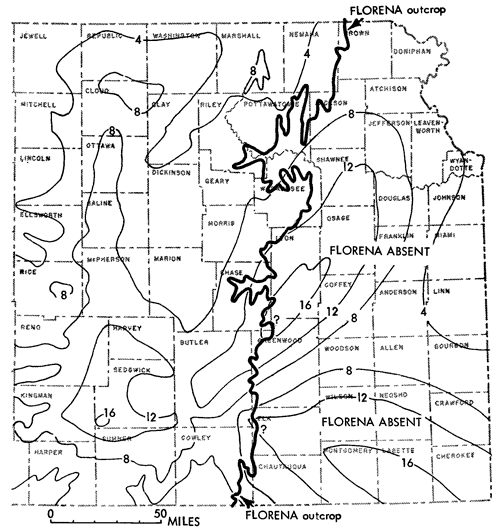
A second point which has important bearing on efforts to define paleogeographic features of Beattie time is relationship of the Beattie sediments to all other Lower Permian deposits in Kansas and bordering regions. The Beattie Limestone is a comparatively thin stratigraphic unit which is merely one of many evenly layered divisions of the rock column. It closely resembles lower and higher marine units containing more or less abundant bottom-dwelling invertebrates, calcareous algae, and scattered remains of fishes. It differs from relatively unfossiliferous red and greenish to drab-gray shales and associated thin fresh- or brackish-water and shallow lagoonal types of marine limestones, which occur between the limestone-and-interbedded-shale deposits of normal salinity. All are approximately equal in horizontal persistence. The similarities of Beattie, Foraker, Red Eagle, Grenola, Bader, Crouse, Funston, Wreford and other predominantly limestone units strongly imply similarities of their paleogeographic patterns. Likewise, but differently, the similarities of such predominantly shale units as the Johnson, Roca, Eskridge, Stearns, Easley Creek, Blue Rapids, Speiser, and others strongly imply similarities of their paleogeographic patterns. This simply means that the whole group of Lower Permian units needs to be studied collectively, with prospect that two related but mutually very dissimilar sets of paleogeographies will emerge. Beattie-type paleogeography should correspond closely to that of the Foraker, Grenola, Red Eagle, and Wreford, etc., though the locations of successive strand lines belonging within and among them might differ appreciably in various quarters. Unpredictably large fluctuations of sea borders must have occurred in passing from Eskridge-type to Beattie-type paleogeog raphy, and from Beattie-type to Stearns-type, etc., not to mention changes of sea outlines within each.
Collective considerations of the Lower Permian marine and associated semimarine and nonmarine deposits in the northern Midcontinent brings to notice recurrent seaway connections extending southwestward from the Hugoton and Oklahoma Panhandle regions leading to eastern New Mexico, western Texas, and south of the Amarillo-Wichita-Arbuckle chain to north-central Texas. With small doubt shallow early Permian seas of the Kansas stable platform extended intermittently northwestward to Wyoming, the western Dakotas (as indicated by Imbrie, Laporte, and Merriam, 1959, p. 71) and beyond (Fig. 35), but this feature of paleogeography which may have included seaways of Beattie time is thought to be much less important than connections southwestward. For the purpose of illustrating this concept of Beattie paleogeography and to demonstrate the highly speculative nature of Beattie-sea outlines plotted on a map, postulated paleogeography very unlike that shown by Imbrie, Laporte, and Merriam is given in Figures 40 and 4l.
Figure 4O--Paleogeography of early part of Beattie cyclothem (late Eskridge) as interpreted by Moore, details entirely subjective and diagrammatic with intent to depict "semimarine" environment of most of Kansas region and presumed shallow but more continuous seaway toward southwest. Sedimentation comparable to parts of Mississippi delta country of present day in southern Louisiana is inferred, with fresh. water lakes, brackish-water lagoons, and broad marshy tracts bordering barely submerged territory. [Included in the Acrobat PDF file of figures for this article.]
Figure 41--Paleogeography of intermediate and late stages in development of Beattie cyclothem in Kansas region. Map actually depicts postulated distribution of submerged and emergent territory in early Stearns time, when dominantly nonmarine deposits in upper part of Beattie cyclothem were formed, with guessed locations of Cottonwood, Florena, and Morrill shore lines left uncovered by retreating shallow sea. Great fluctuations of sea border are inferred to have accompanied very minor recurrent subsidences in the area of sedimentation, possibly combined with effects of small eustatic changes of sea level. [Included in the Acrobat PDF file of figures for this article.]
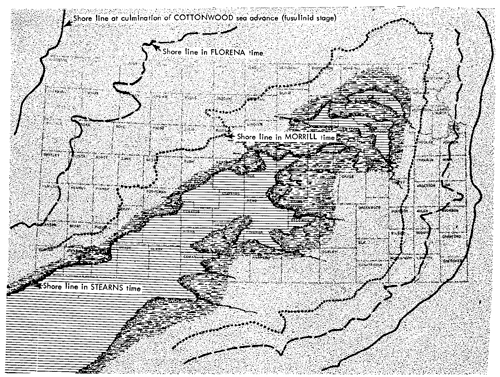
A third point is worthy of comment, though it may not apply universally to Pennsylvanian and Permian cyclothems of the Kansas region or have great importance. This relates to paleoecologic interpretation of fusulinid-bearing deposits in comparison with neighboring ones which lack these foraminifers. Despite incompleteness of development in many cyclothems, the consistency of sequences of fossil assemblages having obviously dissimilar characters led very early (Moore, 1936a; Elias, 1937a) to recognition of paleontologically defined stages or phases in the transgressive and regressive parts of Pennsylvanian and Permian shallow-sea oscillations. At individual localities complete cyclothems may show distinguishable zones which in upward order consist of beds characterized by (1) inarticulate brachiopods or a molluscan assemblage generally dominated by clams, (2) a varied fauna of brachiopods, bryozoans, corals, crinoids, and other marine invertebrates, (3) common to very abundant fusulinids, with or without associated other organisms in large numbers, (4) an assemblage of mainly euryhaline brachiopods (especially Composita), ramose bryozoans, mollusks (including both clams and moderately abundant gastropods), and calcareous algae, and (5) limestone largely composed of such algalforaminiferal colonies as Osagia. Nonmarine strata containing coal beds and land-plant remains or unfossiliferous red and green shales intervene between such sequences of marine deposits. Hattin (1957) and McCrone (1963) have published more detailed ecologic zonation of ideal Lower Permian cyclothems and agree with Elias in classing fusulinid assemblages as markers of deepest water conditions of deposition, correlated with culminations of marine submergences. In my view the occurrence of fusulinid swarms signifies maximum offshore environments, rather than maximum water depths. McCrone (1963, p. 66) agreed with Laporte (1962, p. 541) in estimating that fusulinids are typical of "water depths closer to 50 feet than 160-180 as suggested by Elias."
With respect to Beattie sedimentation, Imbrie, Laporte, and Merriam (1959, p. 78) concluded that "maximum water depths were attained in basin areas during FIorena deposition; and the lower FIorena shelly and Chonetes facies, rather than the fusuline limestone, should be taken as representing the climax of marine transgression. The fusuline and bioclastic facies reflect deposition in turbulent water probably less than 30 feet deep." Laporte (1962, p. 540, 541) added statements that "the Cottonwood shelly facies . . . was certainly formed in an environment as deep as, if not deeper, than that of the fusulinid phase. . . . Fusulines occur relatively near the northern shoreline of the Cottonwood sea and are absent in the offshore position occupied by the shelly facies." On the contrary, fusulinids are in fact associated with the so-called "shelly facies" of the Beattie in southern Kansas and northern Oklahoma (Fig. 36) and no basis is seen for interpreting this area as farther offshore than Beattie locations studied in Nebraska. I see little value, even of hair-splitting academic sort, in debating these points, because it is hardly conceivable that materially different environments are represented by the Tarkio-type (fusulinid) and Threemile-type (bioclastic) of the normal central and northern Kansas Cottonwood and the "shelly" Cottonwood and FIorena with associated fusulinids (Fig. 36-38) and the fusulinid'bearing lower Morrill of southern Kansas. The influx of terrigenous clay and silt seems mainly to account for dissimilarity of the Florena (Neochonetes-Derbyia) biotope and those of associated Tarkio-type fusulinid limestones.
Main conclusions arrived at in this paper can be briefly and simply stated.
(1) Eastern Kansas is a superlative laboratory for studies directed to discriminating and testing criteria needed to guide paleoecological interpretation of cyclically arranged late Paleozoic sedimentary deposits.
Effort to understand the diverse paleobiotopes and ecosystems recorded by these cyclic deposits and to translate this understanding into paleogeographic patterns should begin with the all-important deduction that surfaces of sedimentation throughout enormous areas in the northern Midcontinent region were constantly maintained very close to sea level during late Paleozoic history. It follows that remarkably even subsidence of the earth crust must have kept pace with the accumulations of cyclothems, greater in Oklahoma and southern Kansas than farther north (Fig. 42) but almost identical across Kansas in early Permian time, for example (Fig. 43).
Figure 42--Diagrammatic section of Missorian strata in eastern Kansas and northeastern Oklahoma illustrating southward disappearance of limestones accompanied by thickening of clastic deposits (Moore, based on sections by N.D. Newell). [Included in the Acrobat PDF file of figures for this article.]
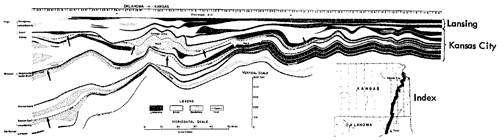
Figure 43--Diagrammatic section of Lower Permian strata extending from northern to southern boundary of Kansas, showing great regularity of "layer-cake" stratigraphy and denoting pulsatory subsidence of almost incredible evenness aggregating approximately 700 feet in north as compared with 800 feet in the south (Moore in Moore and Merriam, 1959). [Included in the Acrobat PDF file of figures for this article.]

An indispensable requirement for paleoecological researches having more than extremely local application is a firmly established stratigraphic framework. This need is satisfied by the large amount of very detailed field work on Pennsylvanian and Permian rocks in Kansas. Also, the excellence of outcrops at many places and the varied kinds of organic remains obtainable from them are valuable contributing factors. Correlated stratigraphic sections given in this paper illustrate a small sample of data available in files of the Kansas Geological Survey.
(2) Many kinds of paleobiotopes and ecosystems can be discriminated in Pennsylvanian and Permian cyclothems of the Kansas region.
In general, each kind exhibits nearly constant characters throughout a very large area, introducing questions as to the precise contemporaneity in development of the very widespread, seemingly continuous biotopes. Similarities of various homologous ecosystems, including both inorganic and organic components, are offset by puzzling dissimilarities. Another feature is the essential identity of ecological communities which are found to recur many times in the stratigraphic succession, some in particular parts of cyclothem groups (megacyclothems). This affords valuable opportunity for comparative analyses.
(3) The selection of a specified type for each discriminated kind of ecosystem facilitates designation of it.
The method of adopting selected paleoecological "yardsticks" simplifies description and aids classification of numerous ecosystems, for all significant attributes of each are tied together as in a labeled package. It is appropriate for paleoecology to borrow from standard practice of paleontology in designating types of species, genera, and (if wanted) families.
(4) The seas which repeatedly transgressed the Kansas region in Pennsylvanian and early Permian time were very shallow, having greatest depth measured in a few tens of feet. Between times of marine submergence, widespread nonmarine and semimarine deposits accumulated virtually at sea level.
The extensive sea invasions can be explained by intermittent eustatic rise of sea level or by crustal subsidence without change of sea level or by both rise of sea level and contemporaneous subsidence (Moore, 1936b) . Of course, water depths at any place during stages of submergence are measured by the magnitude of the causal movements, offset by thickness of sediments accumulating on the sea floor. Diagrammatic representation of repeated fluctuations between water depths of 0 and 200 feet or more (Fig. 44A, B) serves to emphasize their improbability, and a similar graph based on the postulate of subsidence alone as cause of marine invasions (Fig. 44C) is quite unconvincing. Eustatic fluctuations of sea level correlated with recurrent glaciation (Wanless and Shepard, 1936) must be admitted, but for reasons given by Weller (1956, p. 21-22; 1957, p. 362), not to mention other objections, Pennsylvanian and Permian cyclothems and water depths of their marine phases from place to place almost certainly do not reflect such fluctuations alone. To a large extent, the hypothesis of diastrophic control of cyclothems advocated by Weller (1957) is plausible but I can find no good reasons for postulated upward movements of the crust in such areas as the Illinois Basin and Kansas platform alternating with recurrent subsidences (Fig. 44B). Weller (1957, p. 45) has estimated that average uplift of the Illinois Basin surface amounted to 150 feet during the time of making a cyclothem and the average down warping in this interval was 200 feet, but because average thickness of a cyclothem is given as 50 feet, no hint of water depth during marine submergence is found.
Modest eustatic rise and fall of sea level in late Paleozoic time from diastrophic causes (Moore, 1936a) may be assumed reasonably as a corollary of intermittent upthrusting of continental tracts and enlargement of oceanic capacity. Extensive marine inundations of flatland areas of graded sedimentation, as in the northern Midcontinent, would ensue (Fig. 44D).
Figure 44--Diagrammatic representation of fluctuating depth of water in Grenola-to-Funston marine incursions of Kansas region, based on postulates of A, eustatic rise and fall of sea level accompanied by gradual subsidence of accumulated sediments, B, crustal downwarping and upwarping, C (see p. 368), gradual subsidence without reverse upward movements, and D, modest eustatic rise and fall of sea level. [Each line relates to condition at a single station (Wichita, Kansas) in successive moments of geologic time, but simultaneous changes of like sort are to be inferred throughout 100,000 square miles or more in the northern Midcontinent.j
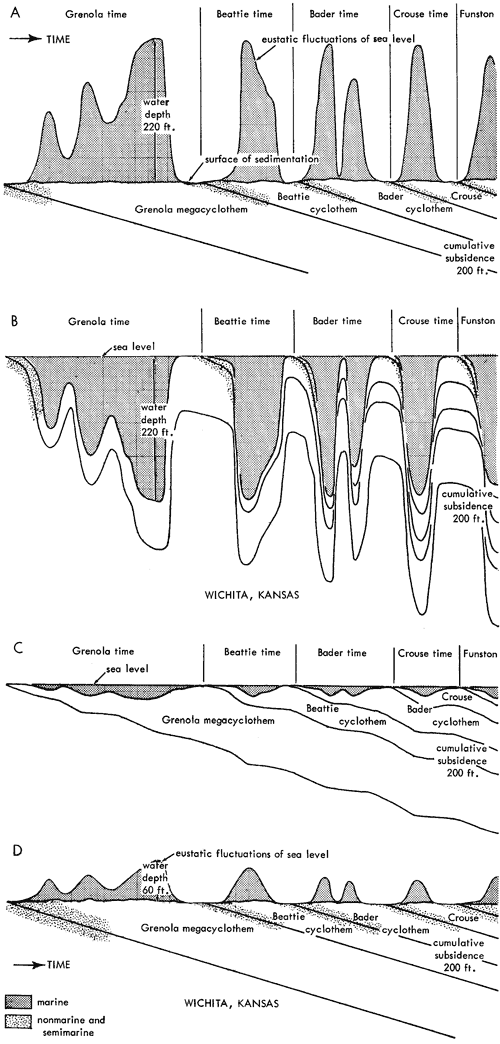
Acer, D. V., 1963, Principles of paleoecology: McGraw-Hill, New York, 371 p.
Baird, Donald, 1952, Revision of the Pennsylvanian and Permian footprints Limnopus, Allopus and Baropus: Jour. Paleontology, v. 26, p. 832-840.
Baird, Donald, 1963, Fossil footprints or stump holes?: Kansas Acad. Sci., Trans., v. 66, p. 397-400.
Baxter, R.W., and Hartman, E.L., 1954, A fossil coniferophyte wood [from Garnett, Kansas]: Phytomorphology, v. 4, p. 316-325.
Beckwith, H.T., 1928, Oil and gas in Oklahoma; geology of Osage County: Oklahoma Geol. Survey Bull. 40-T, 63 p.
Branson, E.B., and Mehl, M.G., 1932, Footprint records from the Paleozoic and Mesozoic of Missouri, Kansas and Wyoming: Geol. Soc. America Bull., v. 43, p. 383-398.
Brues, C.T., Melander, A.L., and Carpenter, F.M., 1954 Classification of insects: Harvard ColI. Mus. Compar. Zool. Bull., v. 108, 917 p.
Carpenter, F.M., 1930-50, The Lower Permian insects of Kansas: Pt. 1, Introduction and the order Mecoptera: Harvard ColI. Mus. Compar. Zool. Bull., v. 70, p. 69-101 (1930); Pt. 2, The orders Palaeodictyoptera, Protodonata, and Odonata: Am. Jour. Sci., ser. 5, v. 21, p. 97-139 (1931); Pt. 3, The Protohymenoptera: Psyche, v. 37, p. 343-374 (1930); Pt. 4, The order Hemiptera and additions to the Palaeodictyoptera and Protohymenoptera: Am. Jour. Sci., ser. 5, v. 22, p. 113-130 (1931); Pt. 5, Psocoptera and additions to the Homoptera: Same, ser. 5, v. 24, p. 1-22 (1932); Pt. 6, Delopteridae, Protelytroptera, Plectroptera, and a new collection of Protodonata, Odonata, Megasecoptera, Homoptera, and Psocoptera: Am. Acad. Arts and Sci. Proc., v. 68, p. 411-504 (1933); Pt. 7, The order Protoperlaria: Same, v. 70, p. 101-146 (1935); Pt. 8, Additional Megasecoptera, Proto donata, Odonata, Homoptera, Psocoptera, Protelytroptera, Plecoptera and Protoperlaria: Same, v. 73, pt. 3, p. 29-70 (1939); Pt. 9, The orders Neuroptera Raphidiodea, Caloneurodea and Protorthoptera (Probnisidae) with additional Protodonata and Megasecoptera: Same, v. 75, p. 55-84 (1943); Pt. 10, The order Protorthoptera, the family Limopteridae and its relatives: Same, v. 78, p. 185-219 (1950).
Carpenter, F.M., 1934, A new megasecopteran from the Carboniferous of Kansas: Univ. Kansas Sci. Bull., v. 21, p. 365-367.
Carpenter, F.M., 1940, Carboniferous insects from the Stanton formation, Kansas: Am. Jour. Sci., v. 238, p. 636-642.
Carpenter, F.M., 1947, Lower Permian insects from Oklahoma. Part 1. Introduction and the orders Megasecoptera, Protodonata, and Odonata: Am. Acad. Arts and Sciences Proc., v. 76, no. 2, p. 25-54.
Condra, G.E., and Dunbar, C.O., 1932, Brachiopoda of the Pennsylvanian System in Nebraska: Nebraska Geol. Survey Bull. 5, ser. 2, 377 p.
Cridland, A.A., Morris, J.E., and Baxter, R.W., 1963, The Pennsylvanian plants of Kansas and their stratigraphic significance: Palaeontographica, v. 112, pt. B, p. 58-92.
Dunbar, C.O., 1923, Kansas Permian insects. Part 2. Paleolimulus, a new genus of Paleozoic Xiphosura, with notes on other genera: Am. Jour. Sci., ser. 5, v. 5, p. 443-454.
Dunbar, C.O., 1924, Kansas Permian insects. Part 1. The geologic occurrence and the environment of the insects: Am. Jour. Sci., ser. 5, v. 7, p. 171-209.
Eaton, T.H., Jr., and Stewart, P.L., 1960, A new order of fishlike Amphibia from the Pennsylvanian of Kansas: Univ. Kansas PubI., Mus. Nat. History Bull., v. 12, p. 217-240.
Elias, M.K., 1934, Cycles of sedimentation in the Big BIue Series of Kansas (abs.): Geol. Soc. America Proc. 1933, p. 235.
Elias, M.K., 1937a, Depth of deposition of the Big BIue (Late Paleozoic) sediments in Kansas: Geol. Soc. America Bull., v. 48, p. 403-432.
Elias, M.K., 1937b, A new scorpion from the Pennsylvanian Walchia beds near Garnett, Kansas: Jour. Paleontology, v. 11, p. 335-336.
Elias, M.K., 1962, Comments on recent paleoecological studies of late Paleozoic rocks in Kansas: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 106-115.
Ellison, S.P., Jr., 1941, Revision of the Pennsylvanian conodonts: Jour. Paleontology, v. 15, p. 105-143.
Ellison, S.P., Jr., 1946-47, Conodonts as Paleozoic guide fossils: Am. Assoc. Petroleum Geologists Bull., v. 30 (1946), p. 93-110; v. 31 (1947), p. 778.
Harbaugh, J.W., 1959, Marine bank development in Plattsburg Limestone (Pennsylvanian), Neodesha-Fredonia area, Kansas: Kansas GeoI. Survey Bull. 134, pt. 8, p. 289-331. [available online]
Harbaugh, J.W., 1960, Petrology of marine bank limestones of Lansing Group (Pennsylvanian), southeast Kansas: Kansas GeoI. Survey Bull. 142, pt. 5, p. 189-234. [available online]
Harbaugh, J.W., 1962, Geologic guide to Pennsylvanian marine banks, southeast Kansas: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 13-67.
Hattin, D.E., 1957, Depositional environment of the Wreford megacyclothem (Lower Permian) of Kansas: Kansas Geol. Survey Bull. 124, 150 p. [available online]
Heald, K.C., 1919, Structure and oil and gas resources of the Osage Reservation, Oklahoma: U. S. GeoI. Survey Bull. 686, p. 213-222.
Henbest, L.G., 1963, Biology, mineralogy, and diagenesis of some typical late Paleozoic sedentary Foraminifera and algal-foraminiferal colonies: Cushman Foundation for Foram. Research Spec. Pub. 6, 44 p.
Hibbard, C.W., 1934, Two new species of Coelacanthus from the Middle Pennsylvanian of Anderson County, Kansas: Univ. Kansas Sci. Bull., v. 21, p. 279-287.
Hotton, N., 1959, Acroplous vorax, a new and unusual labyrinthodont amphibian from the Kansas Permian: Jour. Paleontology, v. 33, p. 161-178. [Speiser Sh.]
Imbrie, John, 1955, Quantitative lithofacies and biofacies study of Florena Shale (Permian) of Kansas: Am. Assoc. Petroleum Geologists Bull., v. 39, p. 649-670.
Imbrie, John, 1959, [Stratigraphic sections of Beattie Limestone, in Road Log, Field Trip No.2]: Kansas Geol. Soc. 24th Field Conf. Guidebook, p. 43-68.
Imbrie, John, Laporte, Léo, and Merriam, D.F., 1959, Beattie Limestone facies and their bearing on cyclical sedimentation theory: Kansas Geol. Soc. 24th Field Conf. Guidebook, p. 69-78.
Johnson, J.H., 1946, Lime-secreting algae from the Pennsylvanian and Permian of Kansas: GeoI. Soc. America Bull., v. 57, p. 1087-1120.
Johnson, J.H., 1963, Pennsylvanian and Permian algae: Colorado School Mines Quart., v. 58, no. 3, 211 p.
Johnson, J.H., and Konishi, K., 1956, Studies of Mississippian algae: Colorado School Mines Quart., v. 51, no. 4, 133 p.
Lane, H.H., 1945, New Mid-Pennsylvanian reptiles from Kansas: Kansas Acad. Sci. Trans., v. 47, p. 381. 390.
Lane, H.H., 1946-47, A survey of the fossil vertebrates of Kansas. Pt. 2, Amphibia; Pt. 3, The reptiles: Kansas Acad. Sci. Trans., v. 48, p. 286-316, v. 49, p. 289-332.
Lane, N.G., 1958, Environment of deposition of the Grenola Limestone (Lower Permian) in southern Kansas: Kansas Geol. Survey Bull. 130, pt. 3, p. 117-164.
Laporte, L.F., 1962, Paleoecology of the Cottonwood Limestone (Permian), northern Mid-continent: Geol. Soc. America Bull., v. 73, p. 521.544.
Loeblich, A.R., Jr., and Tappen, Helen, and others, 1964, Sarcodina, chiefly "Thecamoebians" and Foraminiferida: GeoI. Soc. America and Univ. Kansas Press, Treatise on Invertebrate Paleontology (R. C. Moore, ed.), Part C, Protista, 900 p.
McCrone, A.W., 1963, Paleoecology and biostratigraphy of the Red Eagle cyclothem (Lower Permian) in Kansas: Kansas Geol. Survey Bull. 164, 114 p. [available online]
Marsh, O.C., 1894, Footprints of vertebrates in the Coal Measures of Kansas: Am. Jour. Sci., ser. 3, v. 48, p. 81-84.
Martin, H.T., 1922, Indications of a giant amphibian in the Coal Measures of Kansas: Kansas Univ. Sci. Bull., v. 13, p. 103-114.
Merriam, D.F., 1962, Late Paleozoic limestone "buildups" in Kansas: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 73.81.
Merriam, D.F., 1963, The geologic history of Kansas: Kansas Geol. Survey Bull. 162,317 p.
Moodie, R.L., 1913, Vertebrate footprints in the Lower Permian of Kansas: Am. Jour. Sci., ser. 4, v. 35, p. 31-33.
Moodie, R.L., 1931, The Pennsylvanian vertebrate fauna of Kentucky: Kentucky Geol. Survey, ser. 6, v. 36, p. 351-385.
Moore, R.C., 1929, Environment of Pennsylvanian life in North America: Am. Assoc. Petroleum Geologists Bull., v. 13, p. 459-487.
Moore, R.C., 1930, Sedimentation cycles in the Pennsylvanian of the northern Mid.Continent region: Geol. Soc. America Bull., v. 41, p. 51-52.
Moore, R.C., 1931, Pennsylvanian cycles in the northern Mid-Continent region: Illinois Geol. Survey Bull. 60, p. 247-257.
Moore, R.C., 1936a, Stratigraphic classification of the Pennsylvanian rocks of Kansas: Kansas Geol. Survey Bull. 22, 256 p.
Moore, R.C., 1936b, Stratigraphic evidence bearing on problems of continental tectonics: Geol. Soc. America Bull., v. 47, p. 1785-1808.
Moore, R.C., 1941, Upper Pennsylvanian gastropods from Kansas: Kansas Geol. Survey Bull. 38, pt. 4, p. 121-163.
Moore, R.C., 1949, Meaning of facies: Geol. Soc. America Mem. 39, p. 1-34.
Moore, R.C., 1950, Late Paleozoic cyclic sedimentation in central United States: 18th Internat. Geol. Congress, London, 1948, pt. 4, p. 5-16.
Moore, R.C., 1954, Evolution of late Paleozoic invertebrates in response to major oscillations of shallow seas: Harvard Coll. Mus. Compar. ZooI. Bull., v. 112, no. 3, p. 259-286.
Moore, R.C., 1962, Geological understanding of cyclic sedimentation represented by Pennsylvanian and Permian rocks of northern Midcontinent region: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 91-100.
Moore, R.C., Elias, M.K., and Newell, N.D., 1936, A "Permian" flora from the Pennsylvanian rocks of Kansas: Jour. Geology, v. 44, p. 1-31.
Moore, R.C., and Merriam, D.F., 1959, Kansas field conference, 1959: Kansas Geol. Survey Guidebook, 55 p.
Muir-Wood, H.M., 1962, On the morphology and classification of the brachiopod suborder Chonetoidea: British Mus. (Nat. History), 132 p.
Muir-Wood, H.M., and Cooper, G.A., 1960, Morphology, classification and life habits of the Productoidea (Brachiopoda): Geol. Soc. America Mem. 81, 447 p.
Peabody, F.E., 1952, Petrolacosaurus kansensis Lane, a Pennsylvanian reptile from Kansas: Univ. Kansas Paleont. Contrib., Vertebrata, art. 1, 41 p.
Peabody, F.E., 1957, Pennsylvanian reptiles of Garnett, Kansas: Edaphosaurs: Jour. Paleontology, v. 31, p. 947949.
Peabody, F.E., 1958, An embolomerous amphibian in the Garnett fauna (Pennsylvanian) of Kansas: Jour. Paleontology, v. 32, p. 571-573.
Petrunkevitch, Alexander, 1953, Paleozoic and Mesozoic Arachnida of Europe: Geol. Soc. America Mem. 53, 128 p.
Perkins, R.D., Perry, T.G., and Hattin, D.E., 1962, Some bryozoans from the Beil Limestone Member of the Lecompton Limestone (Virgilian) of Kansas: Kansas Geol. Survey Bull. 157, pt. 5, p. 1-25.
Sayre, A.N., 1930, The fauna of the Drum Limestone of Kansas and western Missouri: Kansas Univ. Sci. Bull., v. 19, p. 75-203. [available online]
Schoewe, W.H., 1956, Geographic locations and stratigraphic horizon of Pennsylvanian fossil footprints discovered in Osage County, Kansas, by B.F. Mudge in 1873: Jour. Paleontology, v. 30, p. 389-393.
Sellards, E.H., 1903, Discovery of fossil insects in the Permian of Kansas: Am. Jour. Sci., ser. 4, v. 16, p. 323-324.
Sellards, E.H., 1906-09, Types of Permian insects: Am. Jour. Sci., ser. 4, v. 22 (1906), p. 249-258; v. 23 (1907), p. 345-355; v.24 (1909), p. 151-173.
Sellards, E.H., 1908a, Fossil plants of the upper Paleozoic of Kansas: Kansas Univ. Geol. Survey, v. 9, p. 386-480.
Sellards, E.H., 1908b, Cockroaches of the Kansas Coal Measures and of the Kansas Permian: Kansas Univ. Geol. Survey, v. 9, p. 501-541.
Tasch, Paul, and Zimmerman, J.R., 1959, New Permian insects discovered in Kansas and Oklahoma: Science, v. 130, p. 1656.
Tillyard, R.J., 1923-37, Kansas Permian insects: Pt. 1, Preliminary announcement: Entom. News, v. 34, p. 292-295 (1923); Pt. 2, Description of a new paleodictyopterid: Am. Jour. Sci., ser. 5, v. 7, p. 203-207 (1924); Pt. 3, The new order Protohymenoptera: Same, ser. 5, v. 8, p. 111-122 (1924); Pt. 4, The order Palaeodictyoptera: Same, ser. 5, v. 9, p. 328-335 (1925); Pt. 5, The orders Protodonata and Odonata: Same, ser. 5, v. 10, p. 41-73 (1925); Pt. 6, Additions to the orders Protohymenoptera and Odonata: Same, ser. 5, v. 11, p. 58-73 (1926); Pt. 7, The order Mecoptera: Same, ser. 5, v. 11, p. 133-164 (1926); Pt. 8, The order Copeognatha: Same, ser. 5, v. 11, p. 315-349, (1926) ; Pt. 9, The order Hemiptera: Same, ser. 5, v. 11, p. 381-395 (1926); Pt. 10, The new order Protoperlaria, a study of the typical genus Lemmatophora Sellards: Same, ser. 5, v. 16, p. 185-220 (1928) ; Pt. 11, Order Protoperlaria, family Lemmatophoridae (continued); Same, ser. 5, v. 16, p. 313-348 (1928); Pt. 12, The family Delopteridae, with a discussion of its ordinal position: Same, ser. 5, v. 16, p. 469-484 (1928) ; Pt. 13, The new order Protelytroptera, with a discussion of its relationships: Same, ser. 5, v. 21, p. 232-266 (1931); Pt. 14, The order Neuroptera: Same, ser. 5, v. 23, p. 1-30 (1932); Pt. 15, The order Plectoptera: Same, ser. 5, v. 23, p. 98-134, 237-272 (1932); Pt. 16, The order Plectoptera (continued), the family Doteridae, with a note on the affinities of the order Protohymenoptera: Same, ser. 5, v. 32, p. 435.453 (1936); Pt. 17, The order Megasecoptera and additions to the Palaeodictyoptera, Odonata, Protoperlaria, Copeognatha and Neuroptera: Same, ser. 5, v. 33, p. 81-110; Pt. 18, The order Embiaria: Same, ser. 5, v. 33, p. 241-251 (1937); Pt. 19, The order Protoperlaria (continued), the family Probnisidae: Same, ser. 5, v. 33, p. 401-425 (1937); Pt. 20, The cockroaches or order Blattaria: Same, ser. 5, v. 34, p. 169-202, 249-276 (1937).
Toomey, D.F., 1964, Lateral homogeneity in a "middle limestone member" (Leavenworth) of a Kansas Pennsylvanian megacyclothem: Rice Univ., Unpub. doctoral thesis, 184 p.
Twenhofel, W.H., 1919, Pre-Cambrian and Carboniferous algal deposits: Am. Jour. Sci., ser. 4, v. 48, p. 339-352.
Wanless, H.R., and Shepard, F.P., 1936, Sea level and climatic changes related to late Paleozoic cycles: Geol. Soc. America Bull., v. 47, p. 1177-1206.
Weller, J.M., 1930, Cyclical sedimentation of the Pennsylvanian Period and its significance: Jour. Geology, v. 38, p. 97-135.
Weller, J.M., 1956, Argument for diastrophic control of late Paleozoic cyclothems: Am. Assoc. Petroleum Geologists Bull., v.40, p. 17-50.
Weller, J.M., 1957, Paleoecology of the Pennsylvanian Period in Illinois and adjacent states: Geol. Soc. America Mem. 67, v. 2, p. 325-364.
Wilson, F.W., 1962, A discussion of the origin of the reeflike limestone lenses of the Lansing Group (Upper Pennsylvanian) of southeast Kansas: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 101-105.
Wray, J.L., 1964, Archaeolithophyllum, an abundant calcareous alga in limestones of the Lansing Group (Pennsylvanian), southeastern Kansas: Kansas Geol. Survey Bull. 170, pt. 1, p. 1-13. [available online]
Zanterl, Rainer, and Richardson, E.S., Jr., 1963, The paleoecological history of two Pennsylvanian black shales: Fieldiana Geol. Mem., v. 4, 352 p.
In response to recommendations made to me by friendly critics after a draft of this paper had been completed, I have prepared a supplement consisting of eight figure assemblages which at least partially illustrate generic characters of fossil organisms found in Pennsylvanian and Permian ecosystems of the Kansas region. It is hoped that this addition will be helpful to readers who are unfamiliar with the named fossils or who have forgotten what they once knew about them. Mostly, the drawings are diagrammatic and show only some external features. Many forms are omitted entirely.
[Please Note--An Acrobat PDF file has been created of the supplementary figures so that they can be presented in higher quality (Acrobat PDF, 1.2 MB).]
Figure 1--Organisms of Asterophyllites, Orbiculoidea-Lingula, Derbyia, Rhombopora, Amblysiphonella and some other ecologic assemblages. [Mags., X 0.3 (Figs. 5, 7), X 0.5 (Figs. 4, 12), X 0.7 (Figs. 1, 2a, 3, 6, 8, 21), X 1 (Figs. 11, 17a, b), X 1.3 (Figs. 17c, 18b), X 2 (Figs. 2b, 10, 18a) , X 3 (Figs. 9, 23), X 7 (Figs. 13-16, 19, 20, 22, 24). Sources: modified from British Museum (Natural History), Figs. 1-8; from "Invertebrate Fossils" by Moore, Lalicker, and Fischer, McGraw-Hill, Figs. 12-24.]

Figure 2--Organisms of Neochonetes, Neochonetes-Derbyia, Pulchratia, and some other ecologic assemblages. [Original, except Fig. 7, from "Invertebrate Fossils" by Moore, Lalicker, and Fischer, McGraw-Hill.]
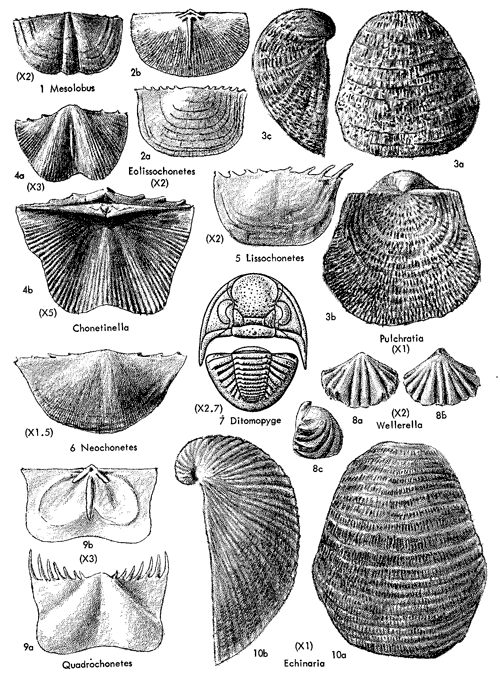
Figure 3--Organisms of Pulchratia and some other ecologic assemblages. [Original.]

Figure 4--Organisms of Caninia and some other ecologic assemblages. [Mags.: X 0.3 (Figs. 6, 8a, 8c, 8d, 9, 13, 19, 21, 23), X 0.7 (Figs. 2-5, 7, 8b, 12, 14, 16, 17, 20, 24), X 1.3 (Figs. 1, 10, 11, 18, 22, 25), X 2 (Fig. 15). Source: "Invertebrate Fossils" by Moore, Lalicker, and Fischer, McGraw-Hill.]

Figure 5--Organisms of Triticites, Ottonosia-Reticulatia, Fenestrellina-Composita, Isogramma, Enteletes, Fenestrellina, and some other ecologic assemblages. [Original, except Figs. 5, 6.9, from "Invertebrate Fossils" by Moore, Lalicker, and Fisher, McGraw-Hill.]

Figure 6--Organisms of Euconospira and some other ecologic assemblages. [Mags.: Fig. 1 (X 0.85), 2, 3 (X 7), 4 (X 3.5), 5 (X 7), 6 (X 2.7), 7 (X 7),8 (X 2.7), 9, 10 (X 0.85), 11 (X 4.5), 12 (X 2.7), 13 (X 0.85), 14 (X 1.1), 15 (X 13), 16 (X 0.85), 17 (X 2.7), 18 (X 3.3), 19 (X 1.3), 20 (X 8), 21 (X 20),22 (X 0.85), 23 (X 1.5), 24 (X 27), 25 (X 0.85), 26 (X 1.8), 27 (X 2), 28 (X 0.85), 29 (X 7). Source: Treatise on Invertebrate Paleontology, Geological Society of America and University of Kansas Press.]

Figure 7--Organisms of Knightites and some other ecologic assemblages. [Mags.: Fig. 1 (X 7), 2 (X 0.85), 3 (X 0.7),4 (X 13), 5, 6 (X 1.3),7 (X 1.8),8, 9 (X 0.85), 10 (X 2.7), 11 (X 1.3), 12 (X 1.8), 13 (X 1.1),14 (X 0.85), 15 (X 1),16 (X 1.8), 17 (X 7), 18 (X 13), 19 (X 2.7), 20 (X 0.7), 21 (X 3.3), 22 (X 0.45). Source: Treatise on Invertebrate Paleontology, Geological Society of America and University of Kansas Press]

Figure 8--Organisms of Archaeolithophyllum,Osagia, Listracanthus, and some other ecologic assemblages. [Sources: Figs. 1-12, from Treatise on Invertebrate Paleontology, Geological Society of America and University of Kansas Press; Figs 14a, 14b, from Wray, Kansas Geological Survey.]