Kansas Geological Survey, Bulletin 170, pt. 1, originally published in 1964
Originally published in 1964 as Kansas Geological Survey Bulletin 170, pt. 1. This is, in general, the original text as published. The information has not been updated.
Limestones of the Lansing Group (Missourian) in southeastern Kansas contain abundant remains of the calcareous marine alga Archaeolithophyllum. Two species, A. missouriense Johnson and A. lamellosum n. sp., are described and illustrated. The thallus of this genus consists of an undulating, irregularly-shaped crust characterized by a distinctive internal cellular structure similar to that in the coralline (red) algae. A. missouriense is believed to have developed as free or locally attached solitary crusts. A. lamellosum grew as encrusting masses made up of superimposed crusts or thalli, and it is intimately associated with bryozoans and encrusting foraminifers.
Archaeolithophyllum is interpreted as having generally inhabited the inner sublittoral marine environment and as having had the capacity to tolerate appreciable wave agitation. The thalli were semi-rigid crusts capable of providing both a self-supporting skeletal framework and a sediment-binding function at the time of deposition. Because of these properties, species of this alga were significant in the formation of initially porous carbonate buildups or banks.
Recent work by Harbaugh (1959, 1960, 1962) has focused attention on marine limestone buildups or banks of late Pennsylvanian age (Lansing Group) in southeastern Kansas. Although many types of marine organisms contributed material to these buildups, Harbaugh has shown that calcareous algae are a dominant biotic constituent. These algae are also believed to have been a depositional influence in the development of these marine banks (Harbaugh, 1962, p. 43). Harbaugh (1959, p. 303; 1960, p. 194) suggested in his early reports that some of these algae, whose appearance is that of leaf-like fragments or crusts, superficially resemble the genus Anchicodium (Johnson, 1946). Others he placed in the algal "family" Spongiostromata. He emphasized, however, that the general absence of preserved internal structure within these algal remains prevented positive identification.
Since 1957 the writer has investigated Pennsylvanian and Permian leaf-like calcareous algae and the limestones that contain them in several areas of the western United States, and some of these studies have been published (Wray, 1959; Konishi and Wray, 1961; Wray, 1962; Pray and Wray, 1963). Kenji Konishi, formerly with the U.S. Geological Survey, and the writer had the opportunity to examine selected samples of the Lansing Group algal facies that were sent to them by Harbaugh in 1959. Neither of us were able to recognize any diagnostic internal structure that would permit positive generic identification. Since Anchicodium had been described from Pennsylvanian rocks in Kansas by Johnson in 1946, it seemed probable that many of these leaf-like forms belonged to this genus.
In September, 1962, the writer attended the Kansas Geological Society's 27th Field Conference on "Pennsylvanian Marine Banks in Southeast Kansas," which was led by John Harbaugh. This provided a first-hand opportunity to examine Lansing Group limestones in the field and to collect algal-rich samples for detailed examination. Subsequently, thin sections of these limestones were prepared and examined. Most of the algae in these carbonate rocks appear in thin section as ribbon-like strips or stringers of sparry calcite devoid of any organic structure, as Harbaugh had pointed out previously; however, remnants of internal structure were found preserved in some fragments. Closer examination revealed that many of the leaf-like fragments or crusts in the Lansing Group limestones are identical with the algal genus Archaeolithophyllum Johnson (1956). Anchicodium is a minor constituent or is absent in the samples studied. Archaeolithophyllum has been recognized previously in limestones of the Lansing Group in Kansas. Eastwood (1958, p. 28) reported Archaeolithophyllum (identified by Kenji Konishi) from the Captain Creek Limestone, but the abundance and significance of this alga throughout the Lansing Group was not discussed.
The purpose of this report is 1) to describe species of Archaeolithophyllum found in limestones of the Lansing Group in southeastern Kansas, 2) to document the occurrence of this genus in rocks that have been the subject of recent petrographic and stratigraphic investigations, and 3) to discuss the paleoecologic and petrologic significance of Archaeolithophyllum. Two species (one new) of Archaeolithophyllum are described and illustrated from two outcrop localities of the Plattsburg Limestone and the Stanton Limestone in Wilson County, Kansas, and from a subsurface occurrence of the Lansing Group in Greenwood County, Kansas (Fig. 1, 2). Locally, Archaeolithophyllum is the dominant biotic constituent in Lansing Group limestones. This alga was an important control in the development of initially porous carbonate facies, because it provided a self-supporting skeletal framework. In some facies secondary porosity of a moldic type has resulted from the dissolution of this alga. For these reasons, rocks characterized by Archaeolithophyllum can be potential petroleum reservoirs.
Figure 1--Map of Wilson and Greenwood counties, Kansas, showing sample localities.
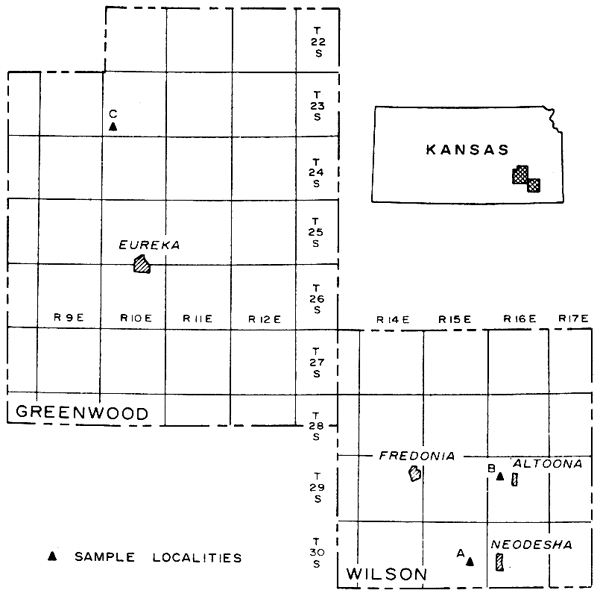
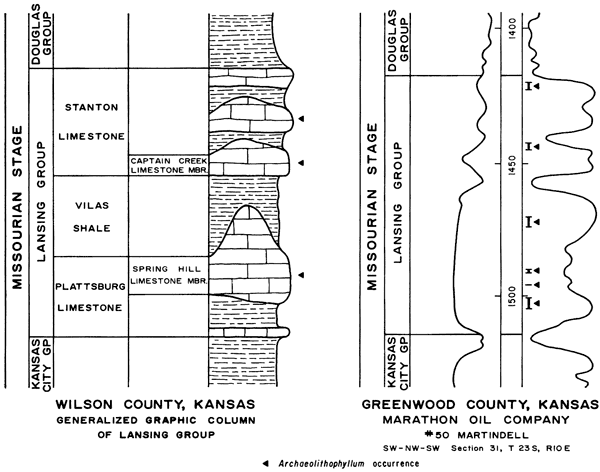
Figure 2--Generalized graphic column of Lansing Group in Wilson County, Kansas (Localities A and B), and subsurface section of Lansing Group in Greenwood County, Kansas (Locality C). Stratigraphic occurrence of Archaeolithophyllum is shown.
Type specimens have been deposited in the U.S. Geological Survey Algal Collection, U.S. National Museum, Washington, D.C.
J. Harlan Johnson, Colorado School of Mines, has been most helpful in supplying source materials and information and in reviewing the manuscript. A. S. Horowitz and L. C. Pray, Marathon Oil Company, read the manuscript and offered suggestions and criticisms. B. A. Upchurch, Marathon Oil Company, did the photography.
The Lansing Group in Wilson County consists of three units. In upward succession, these are the Plattsburg Limestone, Vilas Shale, and Stanton Limestone (Fig. 2). The thickness of these units varies widely. According to Wagner (1962) the Plattsburg Limestone ranges in thickness from less than one foot to about 120 feet. The Vilas Shale ranges from five feet to about 110 feet; the Stanton Limestone varies from 15 to 75 feet. Harbaugh (1959, 1962) has shown that the principal cause of variations in thickness of these units is related to the development of lens-shaped banks in the limestone members. The thickness of the Vilas Shale appears to be inversely related to the thickness of the Plattsburg Limestone in most cases.
The Plattsburg Limestone is divided into two limestone members separated by a shale member. In the present study, species of Archaeolithophyllum are reported from the upper limestone unit (Spring Hill Limestone Member) of the Plattsburg Limestone at two localities.
The Stanton Limestone is divided into three limestone and two shale members. Archaeolithophyllum described herein is from the lowermost member (Captain Creek Limestone Member) at one locality and from the undifferentiated Stanton Limestone at a second locality where members are not distinguished.
Archaeolithophyllum is described from well-core samples from a subsurface section in Greenwood County, Kansas. At this locality, the Lansing Group is 98 feet thick and consists principally of limestone. The electrical-log characteristics of the Lansing Group and parts of the underlying and overlying groups are shown in Figure 2.
Specimens of Archaeolithophyllum described in this report were obtained from the following localities:
| Locality A. | Road cut section along south side of Highway K-96, approximately three miles west of Neodesha, Wilson County, Kansas (Sec. 23, T. 30 S., R. 15 E.). |
| Sample 4 | Spring Hill Limestone Member, Plattsburg Limestone, approximately 30 feet from base of Spring Hill Limestone Member. |
| Sample 11 | Captain Creek Limestone Member, Stanton Limestone, six inches above base of Captain Creek Limestone. |
| Locality B. | Highway K-47, approximately two miles west of Altoona, Wilson County, Kansas (Sec. 18, T. 29 S., R. 16 E.). |
| Sample 2 | Spring Hill Limestone Member, Plattsburg Limestone (U.S.G.S. Paleobotany Loc. D3000). |
| Sample 4 | Stanton Limestone, approximately 10 feet above base. |
| Locality C. | Marathon Oil Company, #50 Martindell, SW NW SW, sec. 31, T. 23 S., R. 10 E., Greenwood County, Kansas. |
| Sample 1420 | Lansing Group (depth 1420 ft.). |
These plates are also available as a higher-resolution Acrobat PDF file (7.4 MB).
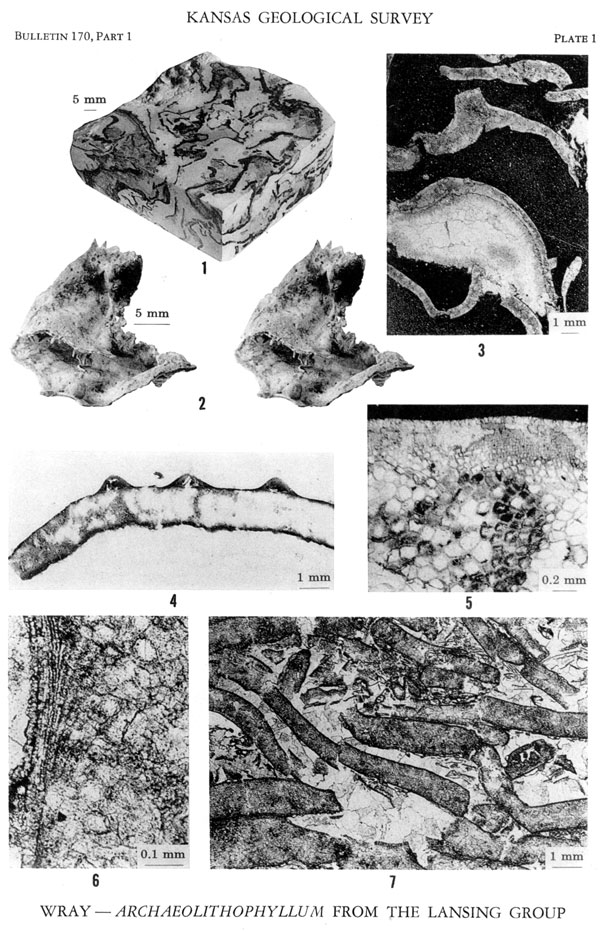
Figures 1, 3-7--Archaeolithophyllum missouriense Johnson. 1. Polished block of limestone composed dominantly of specimens of this species, × 1/2. Captain Creek Limestone Member, Stanton Limestone, Wilson County, Kansas (Locality A, Sample 11). 3. Vertically oriented thin section of specimens. Note sparry calcite filling of original void beneath algal crust, × 4. Capitan Creek Limestone Member, Stanton Limestone, Wilson County, Kansas (Locality A, Sample 11). 4. Polished surface of specimen showing transverse section of three conceptacles, × 7. Captain Creek Limestone Member, Stanton Limestone, Wilson County, Kansas (Locality A, Sample 11). 5. Thin section showing details of hypothallic and perithallic cellular tissue, × 32. Captain Creek Limestone, Stanton Limestone, Wilson County, Kansas (Locality A, Sample 11). 6. Thin section showing rows of perithallic cells arranged vertically on left-hand side of photograph. Larger hypothallic cells are poorly preserved, but discernible, on right, × 90. Lansing Group, Greenwood County, Kansas (Locality C, Sample 1420). 7. Vertically oriented thin section of closely packed specimens with sparry calcite cement filling interstitial areas, × 7. Lansing Group, Greenwood County, Kansas (Locality C, Sample 1420).
Figure 2--Archaeolithophyllum (cf. A. missouriense Johnson). Stereoscopic pair. Silicified specimen showing a compound or envelope growth habit with spines and knobby protuberance, × 1 1/2. Hueco Limestone, Lower Permian, Dona Ana County, New Mexico.
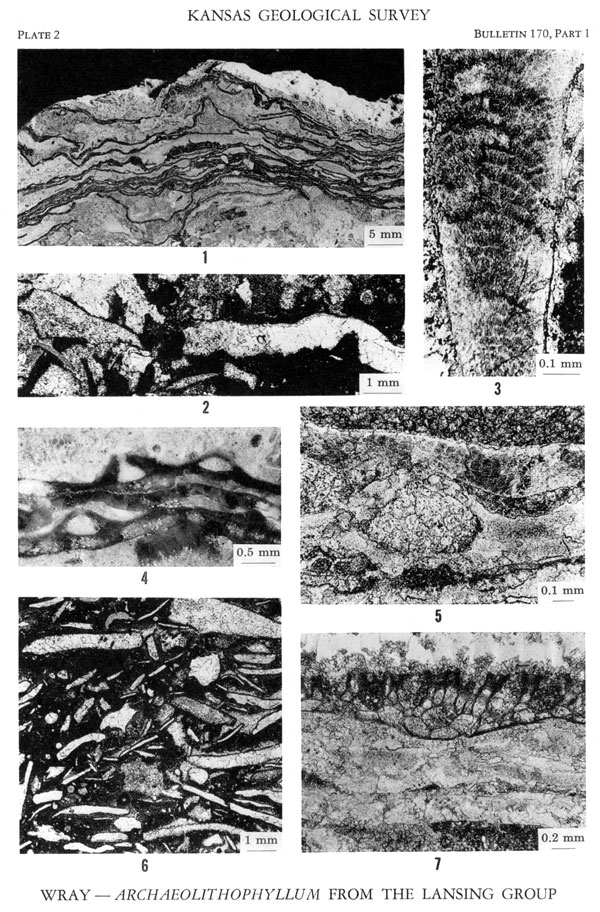
Figures 1, 3-5, 7--Archaeolithophyllum lamellosum, n. sp. Holotype and paratypes. Spring Hill Limestone Member, Plattsburg Limestone, Wilson County, Kansas (Locality B, Sample 2) [U.S.G.S. Paleobotany Loc. D3000]. 1. Vertically oriented polished surface of limestone showing multi-layered crustose growth habit, × 1 1/2 (USGS a1001). 3. Thin section of arcuate rows of hypothallic cells and thin marginal perithallus, × 90 (USGS a1002). 4. Polished surface of several crusts and three conceptacles. A single apical aperture is shown in two of the organs, × 17 (USGS a1003). 5. Holotype. Thin section showing cellular tissue, conceptacle, and multi-layered crustose habit, × 55 (USGS a1000). 7. Thin section of thallus with bryozoan attached to upper surface, × 32 (USGS a1004).
Figures 2, 6--Archaeolithophyllum sp. 2. Thin section of sample showing poorly preserved remnant cellular structure in fragment on left. Internal structure in right-hand end of elongate fragment has been completely obliterated by diagenesis, × 10. Spring Hill Limestone Member, Plattsburg Limestone, Wilson County, Kansas (Locality A, Sample 4). 6. Thin section of sample composed almost exclusively of recrystallized fragments of this alga. Large fragment in upper right-hand corner contains remnant cellular structure, × 7. Stanton Limestone, Wilson County, Kansas (Locality B, Sample 4).
Phylum RHODOPHYCOPHYTA Papenfuss 1946
Class RHODOPHYCEAE Ruprecht 1851
Family UNCERTAIN
Genus ARCHAEOLITHOPHYLLUM Johnson 1956
Types Species--Archaeolithophyllum missouriensum Johnson (1956, p. 54; pl. 14, fig. 1-3, 5) [recte Archaeolithophyllum missouriense].
Diagnosis--Thallus consisting of calcified, undulating, irregularly shaped crusts of variable thickness. Thalli occur as solitary crusts, or foliate and multilayered masses. Crusts attached (some exhibit epiphytic and epizoic habit), or developed free on depositional surface. Knobby and spinose protuberances present on basal and upper surfaces. Internal tissue differentiated into a thick central portion (medullary hypothallus) and thin outer layers (perithallus). Hypothallus composed of arcuate rows of polygonal cells; and perithallus made up of smaller, rectangular cells arranged in rows parallel to surfaces of the thallus. Subconical conceptacles with single apical aperature are distributed irregularly over upper surface of thallus.
Remarks--This genus was based originally on two species, A. missouriense (type species) and A. delicatium [recte A. delicatum], each represented by numerous specimens (Johnson, 1956, p. 53-55). Prior to the present report, no additional species have been described. A. missouriense and one new species, A. lamellosum, are described here from Lansing Group (Missourian) limestones in southeastern Kansas. The original material described by Johnson came from Desmoinesian, Missourian, and Virgilian limestones from seven localities in north-central Texas, Missouri, and Illinois. Johnson remarked that these algae occur abundantly in many Pennsylvanian limestones throughout much of the Midcontinent Region.
The two species designated by Johnson differ mainly in size. The overall thickness of A. delicatum is less than that of A. missouriense. In addition, the hypothallus of A. delicatum is appreciably thinner than in A. missouriense and is composed of smaller cells. A similar, but less pronounced difference, is noted in the perithallus of the two species. The dimensions of the thalli, cells, and conceptacles of these two species as given by Johnson (1956, p. 54-55) are listed in Table 1. Johnson reported that A. missouriense is the most common and widespread species of the genus and reported it from all of the seven localities studied. He stated that Archaeolithophyllum has morphologic features characteristic of the family Corallinaceae and, in particular, the Recent genus Lithophyllum. The internal tissue, consisting of hypothallus and perithallus, and conceptacles are remarkably similar in the fossil and Recent genera. Accordingly, Johnson (1956, p. 54) placed Archaeolithophyllum in the family Corallinaceae. More recently, Johnson (1960, p. 49; 1961, p. 85; 1963, p. 6) has elaborated on the description, interpretation, and systematic position of Archaeolithophyllum, and he now considers it to be one of several genera of calcareous red algae of uncertain affinity, which possess characteristics of the Solenoporaceae and Corallinaceae.
Table 1--Species of Archaeolithophyllum described by Johnson (1956) (measurements in microns).
| Species | Thallus thickness |
Hypothallus thickness |
Hypothallic Cells | Perithallus thickness |
Perithallic Cells | Conceptacles | |||
|---|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | Height | Diameter | ||||
| A. missouriense Johnson 1956 |
500-1800 | 400-1100 | 37-108 | 27-90 | 150-180 | 7-16 | 7-14 | 270-425 | 585-990 |
| A. delicatum Johnson 1956 |
˜200 | ˜135 | 14-36 | 14-22 | ˜135 | 13-15 | 8-10 | 391 | 468 |
Pl. 1, fig. 1, 3-7; Text-fig. 3, 4.
Description--Thallus consisting of calcified undulating crust at least several centimeters in length and width and averaging about 825 µ thick. Crusts developed free on depositional surfaces or are locally attached. Thalli exhibit extreme irregularity in shape and size because of variation in thickness, bifurcations, and degree of curling. Irregularly distributed knobby and spinose protuberances about 500 µ in diameter and up to 1.5 mm in length occur on basal and upper surfaces.
Internal tissue differentiated into a thick central portion (medullary hypothallus) and thin outer layers (perithallus). Hypothallus consisting of arcuate rows of elongate cells (commonly hexagonal) whose long dimension is perpendicular to the rows. Largest cells occur near center of rows and are smallest at margins. Perithallus composed of small rectangular cells arranged in rows parallel to surfaces of thallus. Preservation of perithallic cell structure is generally poor. Perithallus is thickened around conceptacles.
Figure 3--Partial section of thallus of Archaeolithophyllum missouriense showing arrangement of cells in hypothallus and perithallus. Perithallic layers occur on both sides of hypothallus; ×75.

Figure 4--Reconstruction of possible growth habit of Archaeolithophyllum missouriense; approximately natural size.
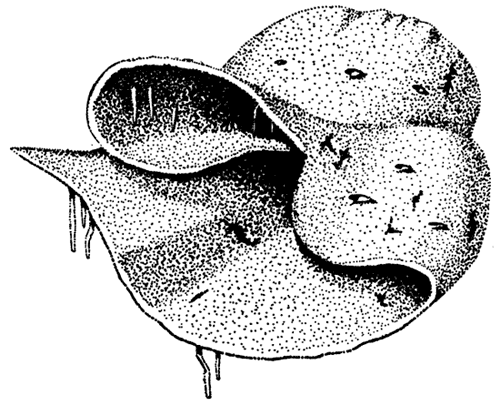
Prominent conceptacles are subconical in shape and distributed irregularly over upper surface of crust. Conceptacle apertures were not observed in Lansing Group specimens. Specimens of this species from Locality C are all infertile, as evidenced by their lack of conceptacles.
Dimensions of thallus, cells, and conceptacles are given in Table 2.
Table 2--Archaeolithophyllum missouriense Johnson (measurements in microns; values given are ranges; means are in parentheses).
| Species | Thallus thickness |
Hypothallus thickness |
Hypothallic Cells | Perithallus thickness |
Perithallic Cells | Conceptacles | |||
|---|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | Height | Diameter | ||||
| A, sample 11. | 300-1500 (775) |
150-1050 (450) |
50-180 (121) |
48-125 (92) |
125-200 (145) |
13-15 (14) |
10-15 (12) |
405-525 (451) |
890-1000 (954) |
| C, sample 1420. | 625-1300 (873) |
490-1175 (850) |
72-152 (108) |
56-136 (86) |
50-65 (60) |
8-12 (10) |
7-10 (8) |
Not Observed | |
Occurrence--Descriptions and detailed measurements were made of specimens of A. missouriense from samples of Lansing Group limestones from Locality A, Sample 11, and Locality C, Sample 1420.
Pl. 2, fig. 1, 3-5, 7
Description--Thallus consisting of calcified, undulating, and foliate crusts at least several centimeters in length and width and averaging about 250 µ thick. Crusts are firmly attached and occur as multiple layers forming masses 2 to 3 cm thick and extending along bedding surfaces for at least 15 cm. Many thalli exhibit epiphytic and epizoic habit. Initial layer may encrust other biotic constituents, commonly bryozoans, or loose sediment. Subsequent layers closely encrust previous ones or are separated by sediment or cavities. Individual crusts occasionally bifurcate. Crusts appear to terminate abruptly without thinning. Knobby or spinose protuberances about 300 µ in diameter and up to 1 mm in length occur infrequently.
Internal cellular tissue differentiated into a prominent, thick central portion (medullary hypothallus), and less distinct, thin outer layers (perithallus). Hypothallus consisting of arcuate rows of elongate, polygonal cells (generally hexagonal) whose long dimension is arranged perpendicular to the rows. The largest cells occur near the center and the smallest at the margins. Perithallus averages only about 40 µ in thickness, but may approach 100 µ around conceptacles. Perithallic cells are nearly rectangular and are arranged in rows parallel to the surfaces of thallus. Precise measurement of the perithallic cells is difficult because of poor preservation in the outer part of the crusts.
Conceptacles are subconical in shape and are distributed irregularly over upper surface of thallus. Conceptacles possess a single apical aperture about 100 µ in diameter.
Dimensions of thallus, cells, and conceptacles are given in Table 3.
Table 3--Archaeolithophyllum lamellosum Wray, n. sp. (measurements in microns; values given are ranges; means are in parentheses).
| Thallus thickness |
Hypothallus thickness |
Hypothallic Cells | Perithallus thickness |
Perithallic Cells | Conceptacles | |||
|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | Height | Diameter | |||
| 30-400 (250) |
30-300 (175) |
20-58 (36) |
18-40 (27) |
20-100 (40) |
12-18 (15) |
10-15 (12) |
220-305 (265) |
450-575 (525) |
Remarks--The superimposed, multilayered encrusting growth form of A. lamellosum is a distinctive feature of this species. This growth habit is not present or well developed in the two previously described species, A. missouriense and A. delicatum. A. lamellosum most closely resembles A. delicatum because both species have comparable crust thicknesses, but the new species differs from A. delicatum, which has smaller hypothallic cells and more elevated conceptacle chambers. A. lamellosum has a thinner thallus, thinner hypothallus and perithallus, and smaller conceptacles than A. missouriense. The new species is named for its characteristic lamellar growth form.
Occurrence--Locality B, Sample 2. (U.S. G.S. Paleobotany Loc. D3000).
Holotype-U.S.G.S. a1000.
Paratypes--U.S.G.S. a1001, a1002, a1003, a1004.
Archaeolithophyllum occurs principally in clastic carbonate rocks in the Lansing Group. This alga and other fossils, both whole and fragmented, constitute the coarser-grained sedimentary particles of the rock. Interstitial or intergranular sediment consists largely of lime mud. Clear calcite cement has filled or partially filled sedimentary voids. Fossil constituents most commonly associated with Archaeolithophyllum are bryozoans, brachiopods, echinodermal fragments, and foraminifers, especially sedentary, tubiform types. Archaeolithophyllum is the dominant biotic constituent in many rocks and, in some, occurs to the exclusion of other fossils.
Conventional limestone classifications are inadequate to characterize accurately most of the rocks of the Lansing Group in which Archaeolithophyllum occurs, although, in general, they could be classified as calcarenites and calcirudites. The recent textural classification of carbonate rocks proposed by Dunham (1962), however, offers a meaningful terminology for these lithologies. This classification utilizes the concept of grain-support versus mud-support and is not dependent on the shape or size of the component particles, except to distinguish grains (> about 0.02 mm) from matrix (< about 0.02 mm). Accordingly, most of the lithologies containing abundant Archaeolithophyllum in the Lansing Group possess grain-supported fabrics and can be classified as packstones or grainstones. Locally where A. lamellosum has encrusted sediment, the rock would be a boundstone.
Unusually large and complete specimens of A. missouriense were found at Locality A, Sample 11 (Pl. 1, fig. 1, 3) in the Captain Creek Limestone Member. Here, undulating crusts up to at least 5 cm in length and width occur in contact with each other and form a grain-supported sediment having a matrix of lime mud. Calcite cement occupies the site of initial pores or cavities in the sediment. The localization of original voids beneath algal crusts in the Lansing Group limestones has been pointed out by Harbaugh (1959, 1960, 1962), using criteria developed by Bathurst (1958). Algal crusts in this rock appear to have developed free on the depositional surface or were only locally attached to it. Lack of obvious fragmentation suggests that they had formed essentially in situ on a predominantly muddy bottom. Other occurrences of A. missouriense in the Lansing Group in Wilson County, Kansas, show various degrees of fragmentation and packing, and the rocks have varying amounts of lime mud matrix and calcite cement.
In contrast to the mud-rich Archaeolithophyllum rocks (wackestones and packstones) studied from Wilson County, the subsurface rocks of the Lansing Group in Greenwood County, Kansas (Locality C), are grain-supported sediments (grainstones) that are almost free of matrix constituents (Pl. 1, fig. 7). These rocks consist principally of two elements, A. missouriense and sparry calcite cement. Algal crusts, many up to 3 cm in length, show extremely close packing. To some extent this is the result of physical compaction, as evidenced by minor brecciation of the algal fragments. These specimens of A. missouriense show a less undulate form and more platy shape that results in better packing. The degree of conformation of thalli may result from superimposed, in situ growth rather than a clastic accumulation, although the evidence for this interpretation is not conclusive. The scarcity of lime mud suggests either a high degree of bottom turbulence, or a high relative rate of production of algae compared to the rate at which mud was produced or transported into the environment. The sparry calcite is interpreted as filling initial open space between algal fragments. Unfortunately, from the standpoint of petroleum reservoirs, the original void space in this rock has been almost completely filled with calcite cement. Analyses of two core plugs of this lithology indicate porosities of 1.7 percent and 3.4 percent with accompanying permeabilities of less than 0.15 md and about 1.0 md. Although these Lansing Group rocks are "tight," rocks of this type can have considerable primary porosities. This rock is remarkably similar to the middle Pennsylvanian porous algal (Ivanovia) limestones that are important as petroleum reservoirs in the Paradox Basin of southeastern Utah (Pray and Wray, 1963). Porosities up to 26 percent and permeabilities as high as about 900 md have been reported from Paradox algal carbonates (McComas, 1963).
The growth habit of Archaeolithophyllum can be reasonably interpreted from its size and shape and sedimentary fabric associations. A. lamellosum had an encrusting growth form that developed multilayered masses several centimeters thick and extended along bedding surfaces for at least 15 cm. At the single occurrence described here (Locality B), this alga locally forms a boundstone in the Spring Hill Limestone Member. This species encrusts trepostome and fenestrate bryozoans, shells, and fine-grained sedimentary particles. Trepostome bryozoans and sedentary, tubiform foraminifers are epiphytic on surfaces of this alga (Pl. 2, fig. 7). A. lamellosum is presumed to have had potential wave-resistant qualities.
A polished block of limestone (Pl. 1, fig. 1) containing A. missouriense shows the configuration of this species in three dimensions. Silicified specimens of a species of Archaeolithophyllum (cf. A. missouriense) (Pl. 1, fig. 2) recovered by acid etching from lower Permian limestones in south central New Mexico further illustrates its growth form. Spinose and knobby protuberances are well shown on this specimen. The comparison of A. missouriense with the form of an ordinary potato chip is a ludicrous one, but it represents a valid comparison of its shape and size with a familiar object. A reconstruction of a possible growth form of A. missouriense is illustrated in Figure 4. Available evidence suggests that in the Lansing Group this species developed as free or locally attached crusts on the sea bottom, though they may have been firmly attached in early stages of growth. This species was apparently able to exist on both carbonite mud bottoms and those characterized by coarser sediment.
The strikingly similar structure of Archaeolithophyllum and the Recent coralline alga Lithophyllum has been pointed out by Johnson (1956, 1956a, 1960, 1961, 1963), and this comparison seems valid to the writer. Johnson indicates that Archaeolithophyllum has an extremely advanced structure, one that did not become common among red algae until Late Mesozoic time. On the basis of morphology alone (size, shape, internal tissue, and reproductory organs), the two genera are very similar. Caution is certainly justifiable in suggesting relationships between these two genera for systematic purposes, and to extrapolate from the known ecology of Recent Lithophyllum to the late Paleozoic Archaeolithophyllum is indeed hazardous. Nevertheless, consideration of the composition, growth form, and habitat of Recent Lithophyllum may provide some useful analogies.
Recent Lithophyllum is exclusively marine and is extensively developed in littoral and sublittoral environments, particularly rocky areas, but it may also occur to depths of at least 100 feet. Since metabolic processes in all algae depend upon sunlight, this ecologic factor strongly affects the development and distribution of species. Consequently, marine algae occur most abundantly in shallow waters.
The growth form of Recent Lithophyllum varies widely. Most species are crustose masses firmly attached to the substratum, although many have extensive free margins. Some species have erect branches or blades that arise from an encrusting base. Thalli range from very thin to thick and massive. It is known to grow on mollusk shells, dead corals, other calcareous algae, and coarse-grained sediment; it does not appear to grow on muddy substrata. Although attachment to hard objects appears essential for most species of Recent Lithophyllum, exceptions have been cited. L. diguettii (Harlot) Heydrich from the Pacific Ocean off the Mexico coast develops free on the sea bottom and has a growth form and size comparable to species of Archaeolithophyllum. Dawson (1960, p. 38-39) describes this species as follows: "Thallus free, 3-5 cm in diameter, consisting of several thin, undulate, anastomosing blades 1-3 cm wide or high arranged somewhat as in a rosette or undulate and in opposing planes, these 1-2 mm thick, . . . ." Foslie and Weber van Bosse (1904, p. 69) note that L. moluccense Foslie, from the East Indies, initially develops attached to hard objects, but it is sometimes detached and continues to grow as a free thallus.
Lithophyllum and other Recent coralline algae typically are composed of high-magnesium calcites. Johnson (1961, p. 18) has summarized the results of 24 chemical analyses of Recent Lithophyllum and reports an average magnesium carbonate content of about 15 percent, with a range between 5 and 25 percent. Studies of the solubility of skeletal carbonates in aqueous solutions by Chave, et al. (1962) indicates that calcites with very high magnesium content are most soluble, low-magnesium calcites least soluble, and aragonites are intermediate. The explanation for the prevalent diagenetic alteration of the wall structure of Archaeolithophyllum is obscure, but, by analogy with Recent Lithophyllum, it may be related to its presumably unstable mineralogy. However, the effect of solubility due to mineralogy was undoubtedly only one factor influencing the alteration of these kinds of algae. The nature of the matrix, chemistry of formation waters, and other factors may have been more important in promoting or retarding diagenetic alteration.
Harbaugh (1959, 1960, 1962, 1963) has fully discussed the importance of leaf-like algae (Archaeolithophyllum) in the development of porous carbonate buildups or banks in the Lansing Group of southeastern Kansas. These algae provided two important influences in the depositional environment: (1) They localized carbonate sedimentation through the mechanism of trapping and binding fine carbonate sediment; and (2) they permitted initial porosity to develop beneath or between leaf-like fragments because of a sheltering effect. Primary intergranular porosity also was formed between coarse algal fragments when finer sediment was not deposited.
Similar depositional influences in the development of porous carbonate facies have been noted for other Pennsylvanian leaf-like algae, namely, Eugonophyllum in Virgilian rocks in New Mexico (Konishi and Wray, 1961; Wray, 1962), and Ivanovia in Desmoinesian rocks in southeastern Utah (Pray and Wray, 1963). Porosity in these rocks also has resulted from the dissolution of leaf-like algae, creating a moldic fabric. Carbonate facies containing leaf-like algal remains are now generally known to be widespread and common throughout marine strata of Pennsylvanian and early Permian ages. Since these algae had the potential capacity to develop porous facies of both biohermal and biostromal character, their proper recognition and interpretation may provide an important clue in the exploration for petroleum reservoirs in carbonate rocks.
In summarizing the paleoecology of Archaeolithophyllum, the following seem significant:
Archaeolithophyllum is relatively simple to identify when its characteristic internal cellular structure is preserved. On the other hand, if the cell structure is destroyed through the process of diagenesis, recognition may be difficult or impossible. As first pointed out by Parks (1958), several genera of calcareous algae with similar leaf-like shapes and sizes existed during Pennsylvanian and Permian time. With their internal structure obliterated, all of these algae appear in two dimensions as ribbon-like strips or stringers of calcite. Unfortunately, the internal structure of these algae is more commonly destroyed than preserved. The problem of identifying recrystallized algal remains is, of course, the reason Archaeolithophyllum has not been more positively recorded in the Lansing Group of southeastern Kansas, even though detailed petrographic and stratigraphic studies of these rocks have been made.
In a discussion of the problems associated with the identification of specific types of late Paleozoic leaf-like algae (Pray and Wray, 1963, p. 209), the term "phylloid" (i.e., resembling a leaf) was proposed to refer to the remains of several commonly occurring calcareous algae having similar leaf-like shapes and sizes which did not retain sufficient internal structure to permit identification. The genera Archaeolithophyllum, Anchicodium, Eugonophyllum, Ivanovia and others can be included in this group. The term "phylloid" applies only to the shape of the algal remains as now observed and is deliberately applied without regard to taxonomy. Phylloid has no implication as to whether the algae developed as encrusting growths, or upright "leaves" or blades. This term simply permits a convenient grouping of similar appearing algal remains. In addition, all phylloid algae are believed to have exerted a similar kind of influence in the depositional environment.
The rather common and related genera Anchicodium, Eugonophyllum, and Ivanovia have a phylloid thallus similar to that of Archaeolithophyllum, although they possess a distinctly different internal structure (Konishi and Wray, 1961). However, recrystallized fragmental remains of these genera are almost impossible to distinguish at the generic level. Anchicodium, Eugonophyllum, and Ivanovia are believed to be green algae (possibly Codiaceae), which have a blade-shaped thallus with an erect growth habit, in contrast to the encrusting habit of Archaeolithophyllum. These related genera also are believed to have inhabited environments similar to that of Archaeolithophyllum, but because of their upright growth habit they probably could not thrive in as highly agitated waters as the crustose types. In thin section, Epimastopora may be confused with other phylloid algae because of a similar size and shape of fragmental remains, but its large distinctive pores serve to differentiate it from others even when it is completely recrystallized.
Bathurst, R. G. C., 1958, Diagenetic fabrics in some British Dinantian limestones: Liverpool and Manchester Geol. Jour., v. 2, p. 11-36.
Chave, K. E., Deffeyes, K. S., Weyl, P. K., Garrels, R. M., and Thompson, M. E., 1962, Observations on the solubility of skeletal carbonates in aqueous solutions: Science, v. 137, p. 33-34.
Dawson, E. Y., 1960, Marine red algae of Pacific Mexico. Part 3, Cryptonemiales, Corallinaceae, subf. Melobesioideae: Pacific Naturalist, v. 2, no. 1, 125 p.
Dunham, R. J., 1962, Classification of carbonate rocks according to depositional texture; in, Classification of carbonate rocks--a symposium: Am. Assoc. Petroleum Geologists, Memoir No. 1, p. 108-121.
Eastwood, W. P., 1958, Stratigraphy of the Captain Creek Limestone (Missourian) of eastern Kansas: Unpublished Master's thesis, Univ. of Kansas, 159 p.
Foslie, M., and Weber van Bosse, A., 1904, Corallinaceae of the Siboga expedition: Siboga Expeditie, Leiden, v. 61, P. 1-110.
Harbaugh, J. W., 1959, Marine bank development in Plattsburg Limestone (Pennsylvanian), Neodesha-Fredonia area, Kansas: Kansas Geol. Survey, Bull. 134, pt. 8, 289-331. [available online]
Harbaugh, J. W., 1960, Petrology of marine bank limestones of Lansing Group (Pennsylvanian), southeast Kansas: Kansas Geol. Survey, Bull. 142, pt. 5, 189-234. [available online]
Harbaugh, J. W., 1962, Geologic guide to Pennsylvanian marine banks, southeast Kansas; in, Pennsylvanian marine banks in southeast Kansas: Kansas Geol. Society, Guidebook, 27th Field Conference, p. 13-67.
Harbaugh, J. W., 1963, Influence of calcareous algae on sedimentation and porosity development in Pennsylvanian limestone banks (abs.): Program, Permian Basin Sec., S.E.P.M. 8th Annual Meeting, p. 23, 25.
Johnson, J. H., 1946, Lime-secreting algae from the Pennsylvanian and Permian of Kansas: Geol. Soc. America, Bull., v. 57, no. 12, p. 1087-1120.
Johnson, J. H., 1956, Archaeolithophyllum, a new genus of Paleozoic coralline algae: Jour. Paleontology, v. 30, no. 1, p. 53-55.
Johnson, J. H., 1956a, Ancestry of the coralline algae: Jour. Paleontology, v. 30, no. 3, p. 563-567.
Johnson, J. H., 1960, Paleozoic Solenoporaceae and related red algae: Colorado School Mines Quart., v. 55, no. 3, 77 p.
Johnson, J. H., 1961, Limestone-building algae and algal limestones: Colorado School Mines, Golden, 297 p.
Johnson, J. H., 1963, Pennsylvanian and Permian algae: Colorado School Mines Quart., v. 58, no. 3, 211 p.
Konishi, K., and Wray, J. L., 1961, Eugonophyllum, a new Pennsylvanian and Permian algal genus: Jour. Paleontology, v. 35, no. 4, p. 659-666.
McComas, M. R., 1963, Productive core analysis characteristics of carbonate rocks in the Four Corners area; in, Shelf carbonates of the Paradox basin: Four Corners Geol. Society, Symposium, 4th Field Conference, p. 149-156.
Parks, J. M., Jr., 1958, Plate-shaped calcareous algae in late Paleozoic rocks of Midcontinent (abs.): Geol. Soc. America, Bull., v. 69, no. 12, pt. 2, p. 1627.
Pray, L. C. and Wray, J. L., 1963, Porous algal facies (Pennsylvanian), Honaker Trail, San Juan Canyon, Utah; in, Shelf carbonates of the Paradox basin: Four Corners Geol. Society, Symposium, 4th Field Conference, p. 204-234.
Wagner, H. C., 1962, A summary of the geology of Wilson County, Kansas, in Pennsylvanian Marine banks in southeast Kansas: Kansas Geol. Society, Guidebook, 27th Field Conference, p. 138-148.
Wray, J. L., 1959, Origin of some Pennsylvanian algal bioherms in southwestern United States; in, Joint Field Conference in the Sacramento Mountains of Otero County, New Mexico: Permian Basin Sect., S.E.P.M. and Roswell Geol. Society Guidebook, p. 38-39.
Wray, J. L., 1962, Pennsylvanian algal banks, Sacramento Mountains, New Mexico; in, Pennsylvanian marine banks in southeast Kansas: Kansas Geol. Society, Guidebook, 27th Field Conference, p. 129-133.
Kansas Geological Survey, Archaeolithophyllum, Lansing Group (Pennsylvanian), Southeastern Kansas
Placed on web March 31, 2009; originally published in June 1964.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/170_1/index.html