![]()
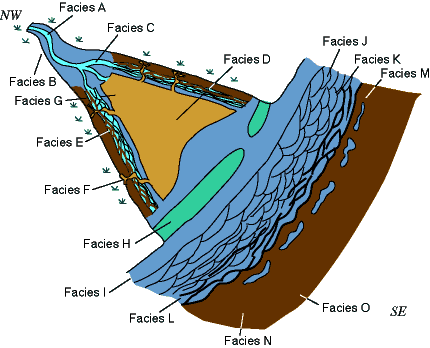
Fig. 62. Depositional model of the lower Morrow Sandstone in the Arroyo and Gentzler fields.

Fig. 63. Idealized vertical succession of Morrow estuarine valley deposits.
General distribution of ichnofossils along the estuarine valley was controlled by the salinity gradient. Trace-fossil analysis suggests a transition from freshwater to brackish-water to near-normal salinity conditions at the seaward end of the estuary valley. However, other parameters, such as oxygenation, substrate and energy, were important at a more local scale, and can be invoked to explain changes in ethology and trophic types from assemblages in high-energy, oxygenated sandy channels to ichnofaunas in low-energy, less-oxygenated, muddy bays.
Open-marine, Morrow trace-fossil assemblages are more diverse and represent the activity of a K-selected, climax fauna developed under stable conditions in a normal marine environment. Monospecific suites are rare and restricted to the upper shoreface or, more rarely, to the middle shoreface. Integration of sedimentologic and ichnologic information allows delineation of a zonation within shallow, open-marine facies (figs. 62 and 64). Ichnologic models of shoreface sandstones are well established for Mesozoic and Cenozoic strata and have been applied to the study of wave-dominated, shallow-marine successions in the subsurface (e.g., Bockelie, 1991; MacEachern and Pemberton, 1992; Pemberton, Van Wagoner et al., 1992; Martin and Pollard, 1996; Howell et al., 1996). However, the ichnology of comparable facies in Paleozoic cores remains poorly understood.
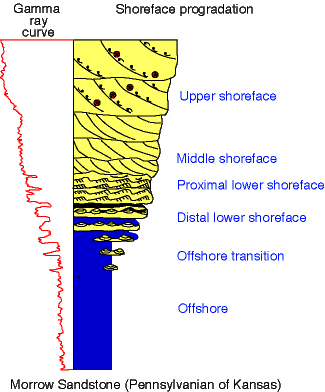
Fig. 64. Idealized vertical succession of Morrow open-marine deposits.
Although shoreface sequences are best developed in the southeast part of the study area in the Nell well, thick intervals, mostly of upper-shoreface facies, also occur towards the top of Gaskill and Kendrick wells, indicating the replacement of restricted valley-fill clastics by regionally extensive, open-marine sandstones. Morrow upper-shoreface facies (facies I) consists of planar crossbedded sandstones and typically lacks trace fossils. In a few places, Palaeophycus isp. and Ophiomorpha irregulaire are present. The common situation, however, is that high-energy, shifting sandy substrates, and high rates of sedimentation and erosion associated with the migration of sand dunes across the upper shoreface, prevented the preservation of biogenic structures. Middle-shoreface facies (facies J) contains abundant dwelling structures of suspension-feeders (Ophiomorpha irregulaire, Skolithos isp., Palaeophycus isp., Diplocraterion isp.) and carnivores (Conichnus isp. and Bergaueria isp.). Escape structures and domiciles of deposit-feeders (Cylindrichnus concentricus, Rosselia isp.) are also common, while feeding traces of deposit-feeders (Teichichnus isp.) are very rare. Trace fossils in the proximal lower-shoreface deposits (facies K) are similar to those of the middle shoreface. However, the degree of bioturbation in the proximal lower shoreface is higher and feeding traces of deposit-feeders (Planolites montanus, Rhizocorallium isp.) are more common. The Morrow distal lower shoreface (facies L) contains an ichnofauna dominated by feeding traces of deposit-feeders (Planolites montanus, Chondrites isp., Rhizocorallium isp., Teichichnus isp., Zoophycos isp.). Domiciles of deposit-feeders (Cylindrichnus concentricus, Asterosoma isp.) are also abundant, while dwelling burrows of suspension-feeders (Palaeophycus isp., Ophiomorpha irregulaire, Skolithos isp., Arenicolites isp.) are subordinated. The offshore-transition facies (facies M) is dominated by feeding traces of deposit-feeders (Chondrites isp., Planolites montanus). Vertical domiciles of suspension-feeders are absent, the only dwelling structure of suspension-feeders being the horizontal Palaeophycus associated with storm deposits. Finally, offshore deposits (facies N) are totally bioturbated, and the ichnofabric is dominated by patterned, deposit-feeding traces, such as Chondrites isp. and Phycosiphon incertum.
The ichnologic-sedimentologic model established for open-marine deposits of the Morrow Sandstone is similar to those proposed for Mesozoic shallow-marine sequences of the Western Interior Seaway by MacEachern and Pemberton (1992) and Pemberton, Van Wagoner et al. (1992). MacEachern and Pemberton (1992) characterized three types of shorefaces based on intensity and frequency of storms: intense, moderate, and weak. The lower-Morrow shorefaces exemplify the weakly storm-affected shorefaces of these authors. These shorefaces display well-developed, fair-weather ichnofaunas reflecting the activity of the resident biota. An onshore-offshore replacement of the Skolithos ichnofacies (dominated by domiciles of suspension-feeders and predators) by the Cruziana ichnofacies (with abundant feeding traces of deposit-feeders) is evident. A transitional zone between both ichnofacies has been detected in the proximal lower-shoreface deposits. The degree of bioturbation increases from the upper shoreface to the offshore deposits. Additionally, while upper- to middle-shoreface facies may display monospecific assemblages, lower-shoreface units typically host multispecific suites.
Similarities between late Paleozoic and Mesozoic, shallow-marine ichnofaunas are remarkable even at the scale of occurrences of individual ichnotaxa. Two differences were noted--namely (1) the presence of small specimens of Ophiomorpha irregulaire with poorly developed wall-linings instead of large specimens of O. nodosa, with thick, pelletoidal walls, typical of post-Paleozoic, high-energy nearshore settings, and (2) burrows of the Glossifungites ichnofacies that are less robust and shallower than those of their post-Paleozoic equivalents.
Moslow and Pemberton (1988) noted that shoreface and delta-front sequences look almost identical on gamma-ray well-log signatures and that distinguishing them requires careful analysis of physical and biogenic sedimentary structures. Deltaic sequences have been described from the Morrow Sandstone in adjacent areas (e.g., Swanson, 1979). As in shoreface sequences, trace-fossil assemblages of delta-front sandstones are characterized by elements of the Skolithos ichnofacies proximally and by traces of the Cruziana ichnofacies distally. However, delta-front sandstones typically contain low-diversity ichnofaunas due to harsh environmental conditions, such as high turbidity and fluctuating salinity and sedimentation and erosion rates (Moslow and Pemberton, 1988). Additionally, evidence of sediment gravity-flow deposition due to slope instability is common. In the cores analyzed, progradational sandstone units display a high diversity of biogenic structures even in relatively proximal situations. This high diversity and abundance of trace fossils suggest deposition in open-marine shorefaces, rather than delta fronts. Moreover, facies analysis indicates interplay of waves and currents, but little evidence of sediment gravity-flow processes.
Kansas Geological Survey
Web version November 9,1999
http://www.kgs.ku.edu/Current/1999/buatois/buatois11.html
email:lbrosius@kgs.ku.edu