![]()
Estuarine ichnofaunas from the lower Morrow resemble brackish-water trace-fossil assemblages documented from post-Paleozoic marginal-marine successions. Morrow estuarine ichnofaunas are characterized by low trace-fossil diversity (nine ichnospecies), reflecting the stressful and harsh conditions of the brackish-water ecosystem. Very few animals have the physiological adaptations necessary to survive under the strong fluctuations in salinity, temperature, and turbidity that characterize marginal-marine environments; consequently, brackish-water biotas are less diverse than marine and freshwater faunas (Croghan, 1983; Barnes, 1984; McLusky, 1989; Hudson, 1990; Pickerill and Brenchley, 1991). Although trace-fossil diversity differs from animal-species diversity (see Bromley, 1996, p. 213-215, for a discussion), the former may provide some basic information on trends in species richness in marginal-marine environments (fig. 57). Marginal-marine, brackish-water ichnofaunas are less diverse than open-marine assemblages (Miller, 1984; Ekdale, 1988; Wightman et al., 1987; Pemberton and Wightman, 1992). The diversity of freshwater animals tends to decline rapidly as salinity increases, but diversity of marine organisms decreases more gradually with the dilution of normal marine salinity (Pemberton and Wightman, 1992; Benyon and Pemberton, 1992) (fig. 57). Therefore, brackish-water trace-fossil assemblages record the activity of depauperate marine biotas, rather than a mixture of freshwater and marine faunas, and are represented by a mixed, impoverished Skolithos and Cruziana ichnofacies (Wightman et al., 1987; Pemberton and Wightman, 1992; Benyon and Pemberton, 1992; Ranger and Pemberton, 1992). Morrow estuarine assemblages consist of marine ichnotaxa (e.g., Teichichnus, Asterosoma), as well as facies-crossing forms (e.g., Planolites, Palaeophycus), but typical freshwater components are absent. In terms of Seilacherian ichnofacies, valley assemblages from the lower Morrow contain representatives of both the Skolithos (e.g., Skolithos, Monocraterion, Arenicolites, Diplocraterion) and Cruziana ichnofacies (e.g., Cruziana, Asterosoma, Teichichnus). Buatois et al. (1997) noted that marginal-marine, brackish-water ichnofaunas are essentially different from those present in the freshwater portions of estuaries, which are dominated by surface trails and trackways. Brackish-water trace-fossil assemblages are dominated by structures of infaunal organisms because the deep-infaunal habitat is a refuge from the rapid and extreme salinity fluctuations at the sediment surface (Sanders et al., 1965; Rhoads, 1975). Estuarine ichnofaunas of the Arroyo and Gentzler fields consist of both shallow-tiered (e.g., Palaeophycus) and relatively deep-tiered (e.g., Arenicolites) structures of infaunal organisms. Additionally, Morrow brackish-water ichnofaunas also are characterized by the presence of very simple forms (e.g., Planolites, Palaeophycus) produced by nonspecialized opportunistic organisms displaying R-selected strategies that enable adaptation to stressful environments (Pemberton and Wightman, 1992; Benyon and Pemberton, 1992). Monospecific suites are present in the lower Morrow valley-fill deposits, particularly in upper- to middle-estuarine settings. Monospecific ichnocoenosis of Palaeophycus are common in upper-estuary channel sandstones, and Planolites tends to occur in monospecific suites in central bay, fine-grained sediments. Small size has been detected in several ichnotaxa from the Morrow estuarine deposits. For example, tiny Palaeophycus and Skolithos are relatively common in upper-estuary channel facies.
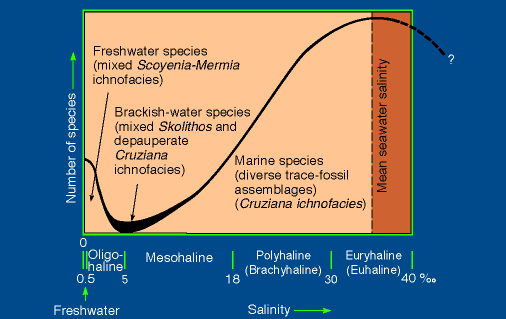
Fig. 57. Relationships among species diversity, ichofaunas, and salinity along a salinity gradient (after Buatois et al., 1997).
Estuarine assemblages from the lower Morrow compare favorably with other brackish-water ichnofaunas recorded from Carboniferous units of the United States. Low-diversity associations, commonly monospecific, of opportunistic infaunal animals in Carboniferous, marginal-marine sequences have been documented by Hakes (1976, 1985), Archer and Maples (1984), Miller and Knox (1985), Devera (1989), Martino (1989), Rindsberg (1990, 1994), Archer (1993), and Mángano and Buatois (1997). Martin (1993) and Miller and Woodrow (1991) also noted similar characteristics in Ordovician and Devonian estuarine ichnofaunas, respectively.
Pennsylvanian, brackish-water ichnofaunas differ from Mesozoic ones in various ways. The Pennsylvanian ichnofaunas are less diverse and are characterized by lower degree of bioturbation, scarcity of crustacean burrows, absence of the Glossifungites ichnofacies in firmground surfaces within the estuary, and absence of specific architectural adaptations to escape extreme salinity fluctuations (e.g., Gyrolithes). These differences are most likely related to evolutionary innovations in Mesozoic estuarine ecosystems, essentially the adaptation of crustaceans to brackish-water, soft to firm biotopes.
Estuarine systems were not recognized in the lower Morrow in Kansas prior to this study. Sedimentologic and ichnologic signatures indicate the presence of a estuarine paleovalley oriented along a northwest-southeast axis and draining to the open sea. Trangressively filled, estuarine successions have been recorded in the lower intervals of Kendrick and Gaskill wells towards the northwest, while open-marine shoreface parasequences are dominant in Nell well, in the southeast part of the study area. Lateral to Kendrick, at Fretz well, interfluve deposits occur. Integration of ichnologic data with sedimentologic, stratigraphic, and paleogeographic information was crucial in the recognition of valley-fill sandstones in the lower Morrow and in their distinction from open-marine shoreface deposits. The discovery of a low-diversity trace-fossil assemblage that records the activity of a depauparate marine fauna was essential in the recognition of estuarine valley facies. This trace-fossil assemblage contrasts with the more diverse and abundant ichnocoenoses of the open-marine shoreface facies.
The Lower Morrow estuarine system is similar to the wave-dominated estuary characterized by Dalrymple et al. (1992). The valley system recorded in the Arroyo and Getzler fields displays a clearly defined, tripartite division into (1) a high-energy, marine-sand plug formed at the seaward end of the valley (estuary mouth), (2) a low-energy, fine-grained middle zone (central bay), and (3) a high-energy inner zone dominated by the discharge of fluvial tributaries (bay-head delta). Tidal influence, however, is suggested by the presence of reactivation surfaces, mud drapes, and cross-lamination dipping in opposite directions in bay-head delta sandstones (facies C) and tidal-flat heterolithic deposits (facies E), as well as the existence of tidal rhythmites (facies F) in intertidal channels.
The base of the estuarine valley was recognized in the Gaskill well at the bottom of facies C interval (6,065.4 ft; 1,848.7 m) and is coincident with the Mississippian-Pennsylvanian boundary (320 +/- 10 my). A major sea-level fall that took place during this time (Ross and Ross, 1988) is thought to be responsible for valley incision. The estuarine valley was subsequently filled during sea-level transgression. The replacement of low-diversity, brackish-water trace-fossil assemblages by the open-marine Cruziana ichnofacies supports this interpretation. A second sequence boundary was identified at the base of a forced-regression shoreface package in the Gentzler field, where the firmground Glossifungites ichnofacies has been detected (figs. 58, 59, 60, 61).
 |
 |
|||||
| Figure 58. | Figure 59. | Figure 60. | Figure 61. |
Kansas Geological Survey
Web version November 9,1999
http://www.kgs.ku.edu/Current/1999/buatois/buatois10.html
email:lbrosius@kgs.ku.edu