Kansas Geological Survey, Subsurface Geology Series 4, originally published in 1981

Originally published in 1981 as Kansas Geological Survey Subsurface Geology Series 4. This is, in general, the original text as published. The information has not been updated. An Acrobat PDF file of this report is available (1.4 MB).
It is important for those engaged in petroleum exploration to be able to correctly classify the age of rocks. Solving these stratigraphic problems is aided by palynology, the study of fossil plant pollen and spores. Different kinds of organic-walled microfossils prevail in different ages of earth history. Land plant spores are abundant in rocks as old as the Devonian (400 to 345 million years ago); pollen grains are useful to date Mesozoic (225 to 65 million years ago) and Cenozoic (65 million years ago to present) rock layers, with different kinds of grains characterizing different time periods. Lower and middle Paleozoic (570 to 345 million years ago) rocks, however, do not contain these fossils common to younger sedimentary rocks. Instead they contain acritarchs and chitinozoans that were part of the oceanic plankton during the time the sediments were deposited.
We have examined new material to establish a reference section for further correlation of groups of fossils of the same age from other basins. The following report is a description of the microplankton from the Ordovician (500 to 440 million years ago) in the subsurface of northeastern Kansas. In biostratigraphy (the study of rock units through their fossil contents), the first step is to verify a series of well-defined and easily recognizable fossil assemblages from independently dated stratigraphic horizons. These assemblages from rocks of known age can then be used to correlate with assemblages from rocks not well dated. The Maquoketa Shale microplankton described in this report will serve as an important reference for the Late Ordovician in other areas. Our ultimate goal is to assist with oil exploration programs in new areas by routinely answering questions of stratigraphic correlation and environmental interpretation using acritarchs and chitinozoans. Although many questions remain unanswered concerning acritarchs and chitinozoans, the list of stratigraphic horizons in which these groups have been found is growing. This definition of a Maquoketa-Viola assemblage is the first reported occurrence of lower Paleozoic microplankton from Kansas.
The purpose of this study is to record the acritarchs and chitinozoans from the Upper Ordovician of northeastern Kansas. In so doing, the census of these fossils, not previously reported from this area, is available for comparative biostratigraphic studies. Well-preserved acritarchs and chitinozoans were recovered from conventional core samples of the Maquoketa Shale and Viola Limestone, a marine sequence from the Cities Service Company #1-A Steele-Griffee well located in Nemaha County. Four species of chitinozoans and 27 species of acritarchs were found and referred to species of previously described genera. Of these species, all occur in the Maquoketa Shale whereas only five acritarch species were found in the Viola Limestone. This greater diversity of microplankton in the shale units of the Maquoketa versus the lack of it in the carbonate units of the Viola indicates that the Maquoketa sediments were deposited in quiet water of a more offshore area than those of the Viola. More specifically, the baltisphaerid-Polygonium groups (open sea) dominate the Maquoketa Shale assemblage whereas the leiosphaerid-veryhachid groups (nearshore shallow water) dominate the Viola assemblage. These findings are consistent with those of Jacobson (1979) for Middle and Upper Ordovician rocks from Kentucky, Ohio, and New York and indicate that the species diversity of microplankton may be a useful tool to predict specific marine environments.
Palynology is well suited to solving stratigraphic problems for petroleum exploration programs because different kinds of organic-walled microfossils prevail in different ages of earth history. Land plant spores are abundant in rocks as old as the Devonian; pollen grains are particularly useful to date Mesozoic and Cenozoic strata, with striate bisaccate grains characteristic of Permo-Triassic rocks; and dinoflagellates are important biostratigraphic tools in Middle Jurassic through Tertiary rocks. Lower and middle Paleozoic rocks do not contain these fossils common to younger sedimentary rocks but often contain assemblages rich in acritarchs and chitinozoans that were part of the marine plankton during the time of deposition of these sediments.
Cities Service Company has exploration interests and drilling programs in interior basins where Paleozoic rocks are encountered and where acritarchs and chitinozoans have been used as time-stratigraphic markers. In the future, regions such as the eastern overthrust belt of the Appalachians and the western overthrust belt of the Cordillera may put acritarchs and chitinozoans to the test as biostratigraphic tools. In order to begin to construct a correlation between assemblages from different geographic areas, new material is being examined at the Research Laboratory to establish reference sections by their assemblages. The following report is a description of the Ordovician microplankton from the subsurface of northeastern Kansas. In biostratigraphy, the first step prior to the application of a biostratigraphic tool is to verify a series of well-defined and easily recognizable fossil assemblages from independently dated stratigraphic horizons. These assemblages then can be used to correlate with assemblages from rocks of uncertain age. The sequence of microplankton from the Maquoketa Shale will serve as an initial reference section for the Late Ordovician in Kansas. Our ultimate goal is to assist with oil-exploration programs in new areas by routinely answering questions of stratigraphic correlation and environmental interpretation using acritarchs and chitinozoans. Although many unanswered questions of acritarch and chitinozoan synonymy, identification, and stratigraphic correlation exist, the list of stratigraphic horizons in which species of these groups have been found is growing. This description of a Maquoketa-Viola assemblage adds to that list as it is the first reported occurrence of lower Paleozoic microplankton from Kansas.
During the past few years, acritarchs and chitinozoans have been found to be common in Ordovician rocks in North America. Their biostratigraphic utility is demonstrated by the fact that Upper Ordovician strata in the Williston Basin of Montana and South Dakota, and Anticosti Island, Canada (Jacobson, personal communication), and in Ohio, Indiana, Kentucky, and Oklahoma contain comparable microplankton assemblages (Colbath, 1979; Loeblich, 1970; Loeblich and Tappan, 1978; Jacobson, 1978, 1979). Because acritarchs and chitinozoans are present in many marine rocks of early and middle Paleozoic age, are microscopic in size, and are easily extracted from the enclosing rock matrix, they lend themselves to solving stratigraphic problems dealing with subsurface geology. They are particularly useful in subsurface units that lack other diagnostic fossils such as corals, brachiopods, conodonts, stromatoporoids, and ostracods. The biostratigrapher may, therefore, use acritarchs and chitinozoans in conjunction with electric logs and lithologic analyses to provide a picture of the subsurface stratigraphic framework that is important in exploration.
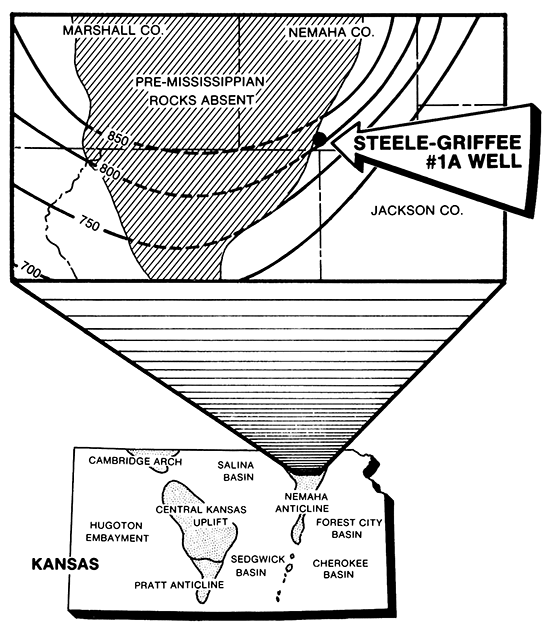
The purpose of this study is to describe a well-preserved assemblage of fossil microplankton from the Maquoketa Shale/Viola Limestone (Ordovician) obtained from the subsurface of Kansas. The fossils described in this report were recovered from a core in the Cities Service Company #1-A Steele-Griffee well located in Nemaha County, Kansas (Fig. 1). Preliminary examination of a few samples revealed the presence of acritarchs and chitinozoans. Additional sampling was done to determine if there were differences in assemblages above and below the unconformity between the Maquoketa Shale and Viola Limestone. The fossils described here extend our knowledge of Late Ordovician acritarchs and chitinozoans to the subsurface of northeastern Kansas. It is our hope that this study will stimulate interest to gather similar data on these fossils and apply that data to biostratigraphic problems in oil and gas exploration as well as to correlation in the subsurface of Kansas.
Figure 1--Location of C.S.C. #1-A Steele-Griffee well and thickness of Ordovician and Silurian rocks in feet.
Dry samples, averaging 50 grams in weight, were taken from a conventional core at closely spaced vertical intervals (Fig. 2). The samples were crushed to fragments approximately five millimeters in diameter, placed in beakers, and covered with water. Cold dilute HCl was added to begin dissolution of the carbonates. When the bubbling reaction ceased, concentrated HCl was added to ensure complete carbonate dissolution. The samples were then washed four times with distilled water, and the liquid decanted each time after the residue settled to the bottom of the beaker. Concentrated HF was then added to remove silicates. The HF bath was allowed to stand for two days before waterwashing. The palynomorphs were then separated from the remaining residue by using the zinc bromide flotation method. No oxidation treatment was employed and the palynomorphs were not darkened with a safranine stain. Where possible, scanning electron microscope photographs accompany Nomarski photomicrographs taken using a Zeiss Photomicroscope III. The fossil material is stored in the palynological collections of Cities Service Company's Research Laboratory, Tulsa, Oklahoma.
Figure 2--Characteristic shale lithology of the Maquoketa with the clastic zone at the base just above the unconformity with Viola limestone. Rock sample locations shown.

Paleozoic rocks are present in the subsurface of Kansas. Rising arches and subsiding basins in this area of the craton influenced the early and middle Paleozoic depositional framework and later affected the subcrop distribution of the strata. In eastern Kansas, a change in structural development took place in post-Mississippian pre-Desmoinesian time (Merriam, 1963) with the rise of the Nemaha Anticline. The anticline eliminated the North Kansas Basin, which was the depositional site of Ordovician through Mississippian sediments, by dividing it into the Salina and Sedgwick Basins on the west and the Forest City and Cherokee Basins on the east. The trace of the Nemaha Anticline crosses Kansas from Nemaha County in the north to Sumer County in the south. The anticline is pronounced in the subsurface with Precambrian granite subcropping along its crest, faulting along its east flank, and upturned and truncated pre-Pennsylvanian rocks along both flanks. This structural trend has been the site of intense oil exploration.
The C.S.C. #1-A Steele-Griffee well is situated on the eastern flank of the Nemaha Anticline in Nemaha County and penetrates Pennsylvanian through Ordovician strata. The Maquoketa Shale, which lies within the 800-foot thick Ordovician and Silurian rock sequence, occurs between 3095 and 3168 feet in the well and has a characteristic electric-log response (Fig. 3). The pre-Mississippian subcrop pattern indicates the Maquoketa is truncated and eroded, being absent in the subsurface south of a transect extending from Leavenworth County in the northeast to Sedgwick County in the southwest.
Figure 3--E-log characteristics of Maquoketa Shale and adjacent rocks.
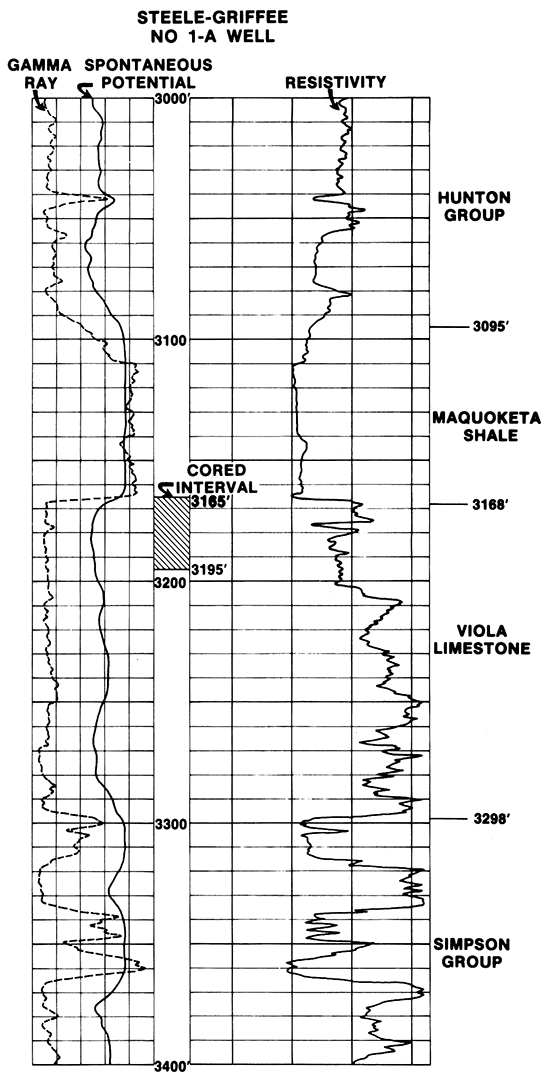
In the C.S.C. #1-A Steele-Griffee well, the Maquoketa is greenish-gray shale. The formation unconformably overlies the Viola Limestone. The unconformity is considered to occur just below the pyrite-laden clastic zone that contains shale with reworked dolomite and crinoidal debris (Fig. 2). This clastic zone is recognized in the Kansas subsurface elsewhere (Lee, 1956) and is possibly similar to the pyritic, shell-clastic zone at the base of the Maquoketa Shale reported from Illinois (Willman and Buschbach, 1975).
The acritarchs and chitinozoans recovered from all the samples are well preserved. The chitinozoans are light amber-brown and the acritarchs are light yellow, indicating that they have a low thermal alteration. In general most of the samples of the Maquoketa Shale yielded abundant acritarchs along with chitinozoans. Microplankton from the Viola Limestone were either scarce or absent (Table 1). Although relatively little information has been published concerning the distribution of acritarchs and chitinozoans in terms of paleoenvironments and lithotypes, Staplin (1961, p. 396, 397) concluded in his study of the microplankton distribution of Devonian forms in the Leduc Reef region of Alberta that (1) "hystrichosphaerids [acritarchs] are much more abundant in off-reef than in near-reef strata," and that (2) "simple, spherical forms occur all the way from shales interbedded with reef carbonates to off-reef areas; and thin-spined forms are also wide-spread but are seldom found within one mile of reef, and that thick-spined and polyhedral forms occur in off-reef areas." Similarly, Wall (1965) observed from the Early Jurassic (Lias) of England that species-rich acritarch assemblages accumulated in offshore environments whereas species diversity decreased and the dominance by a single species increased inshore. Relative to the Upper Ordovician, Jacobson (1979) found that leiosphaerids (simple unornamented spheres) dominated nearshore areas, and the baltisphaerid (acritarchs with processes) veryhachid-Polygonium-micrhystridid association represents offshore open-marine environments. Unpublished results by one of the authors of this paper (R.P. Wright) show that the number of species and different morphologic forms of acritarchs from the Ordovician Bromide Formation of Oklahoma increase in basinal shales deposited away from the nearshore areas.
Table 1--Stratigraphic occurrence of microplankton.
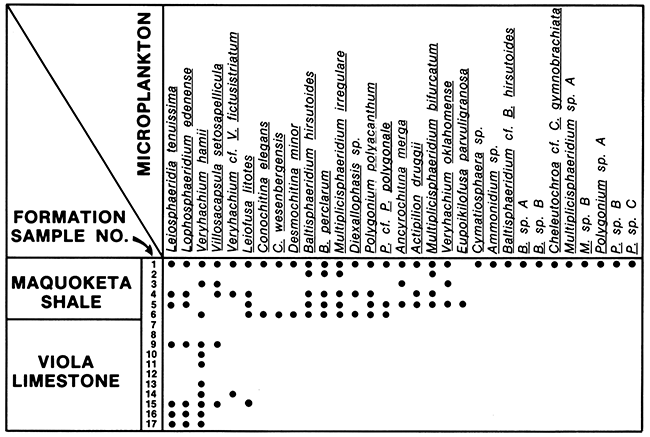
A census of the microplankton from this study indicates that the shales of the Maquoketa, dominated by the Polygonium-baltisphaerid association, represent the quiet, deeper water of an offshore area whereas the carbonates of the Viola, dominated by simple spheres (leiosphaerid-lophosphaerid), represent inshore areas. This interpretation, based independently on the microplankton data, is consistent with the results of Staplin, Wall, and Jacobson. As a diagnostic method it would seem, therefore, that assemblage diversity as mapped by numbers of different species and morphologic forms of acritarchs would be an effective tool to predict specific marine environments and transgressive-regressive phases of cycles of sedimentation.
Compared with previously studied Ordovician microplankton assemblages (Fig. 4), the Maquoketa Shale-Viola Limestone sequence in the C.S.C. #1-A Steele-Griffee well contains acritarchs characteristic of the Llandeilian, Caradocian, and Ashgillian (late Middle to Late Ordovician) ages.
Figure 4--Stratigraphic positions of formations examined for Ordovician microplankton.
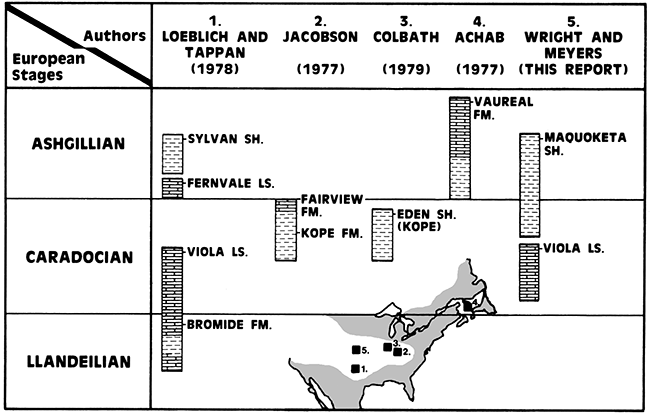
As documented in the systematic section, 27 species of acritarchs were recognized and assigned to 14 genera. Although several of the species are known from older Ordovician rocks in North America and Europe, their collective occurrence indicates a Late Ordovician age. The Ashgillian Sylvan Shale assemblage is the most similar by comparison with Leiofusa litotes, Baltisphaeridium perclarum, Cheleutochroa gymobrachiata, Eupoikilofusa parvuligranosa, Actipilion druggii, Veryhachium hamii, V. oklahomense, and Villosacapsula setosapellicula common to both the Sylvan and Maquoketa Shales. The older (Caradocian) Kope and Fairview Formations have Leiosphaeridia tenuissima, Baltisphaeridium hirsutoides, Polygonium polyacanthum, P. cf. P. polygonale, and Multiplicisphaeridium bifurcatum in common with the Maquoketa. Only Villosacapsula setosapellicula is common to both the Maquoketa and Eden Shale although both assemblages are similar at the generic level. Recently Martin (1980) has reported Multiplicisphaeridium irregulare and Baltisphaeridium cf. B. perclarum, species common to the Maquoketa, from the Caradoc-Ashgill White Head Formation of Quebec, Canada. Occurring along with the acritarchs are Ordovician chitinozoans. Conochitina wesenbergensis is known to occur in the Viola Limestone, and Desmochitina minor occurs in the Viola and Fernvale Limestones and lower part of the Sylvan Shale. Ancyrochitina merga is diagnostic of the Ashgillian Sylvan Shale and Vaureal Formation. Conochitina elegans.is common to both the Maquoketa and Sylvan Shales.
Both acritarchs and chitinozoans have organic walls and are concentrated in the laboratory by the same techniques. In spite of their common occurrence and abundance, the biological affinity of each group remains in doubt. Chitinozoa have been considered protozoans (Eisenack, 1931; Collinson and Schwalb, 1955), metazoans and possibly egg capsules of invertebrates such as annelids and gastropods (Kozlowski, 1963), graptolites (Jenkins, 1970), and crysophyte algae (Staplin, 1961). Most students of the group believe that they belong to the animal kingdom. The group Acritarcha was proposed by Evitt in 1963 for nondinoflagellate hystrichospheres. Acritarchs have been considered, at various times, as either a polyphyletic association of organisms representing for the most part the reproductive stages of marine planktic algae (Downie and others, 1963) or a homogeneous group of algae (Eisenack, 1938; Klement, 1960; Madler, 1963). In this report the acritarchs assignable to the genera Cymatiosphaera and Leiosphaeridia have been placed with the green alga Prasinophyta (Boalch and Parke, 1971), with the remainder of the acritarchs included in Algae incertae sedis.
We have listed the chitinozoans and acritarchs in alphabetical order because their unknown affinity precludes placing them in a biological classification scheme. Describing the genera in alphabetical order will also facilitate the use of this paper. A description of the morphologic terminology used here for chitinozoans is that of the Commission Internationale de Microflore du Paleozoique (Combaz and others, 1967; Laufeld, 1974), and for acritarchs that of Tappan and Loeblich (1971), Kjellstrom (1971), and Colbath (1979).
Type species: Ancyrochitina ancyrea Eisenack 1955
Ancyrochitina merga Jenkins 1970
Pl. 1, Figs. B, C
Diagnosis: The cylindro-conical vesicle is ornamented with numerous simple or branching basal-edge spines that are about 1/3 to 1/4 the length of the vesicle. The spines have a Y or T pattern. The oral tube may be slightly flared in an oral direction. The vesicle wall is smooth or possesses small spines.
Remarks: Jenkins (1970) described this species from the lower part of the Sylvan Shale in Oklahoma which is Ashgill in age determined by the occurrence of the graptolite Dicellograptus complanatus. The Maquoketa specimens, like those from the Sylvan Shale, possess the characteristic Y- and T-shaped basal-edge spines. Representatives of this species reported from the D. complanatus Zone of subsurface Vaureal Formation, Anticosti Island, Quebec, by Achab (1977) possess third-order branching unlike the Sylvan or Maquoketa specimens.
Occurrence: This species is found infrequently in samples 1, 3, and 5 of the Maquoketa Shale.
Type species: Conochitina claviformis Eisenack 1931
Conochitina elegans Eisenack 1931
Pl. 1, Figs. F, I, J, K
Diagnosis: The simple cylindrical vesicles may be up to 600µ in length although most forms are between 300 and 500µ. A slight aboral swelling is present on some specimens. The vesicle wall is usually smooth but the aperture may be fringed with small spines.
Remarks: Jenkins (1970) noted an abundance of C. elegans from the lower part of the Sylvan Shale in Oklahoma. Rhabdochitina hedlundi Taugourdeau 1965 from the Maquoketa Formation in Iowa is probably conspecific with C. elegans. Conochitina elegans is present from the upper Llandeilo through the Caradoc in Estonia and from the Caradoc in the Welsh Borderland (Jenkins and Legault, 1979).
Occurrence: This species is abundant in samples 1 and 6 of the Maquoketa Shale. Many specimens possess a long prosome that can be observed when the specimens are viewed in transmitted light.
Conochitina wesenbergensis Eisenack 1959a
Pl. 1, Figs. G, H
Diagnosis: The vesicle varies in shape from being conical to conical with an aboral bulge. Simple spines occur in the vesicle wall with their size decreasing in an oral direction and nearly aligned in rows parallel to the long axis of the vesicle.
Remarks: The Maquoketa specimens are similar in shape, size, and density of ornamentation to those described from the Viola Limestone in Oklahoma by Jenkins (1969) and more closely resemble C. micracantha wesenbergensis and C. micracantha micracantha as reported from the Ostseekalke of the Baltic region by Eisenack (1965).
Occurrence: This species is abundant in samples 1 and 6 of the Maquoketa Shale.
Type species: Desmochitina nodosa Eisenack 1931
Desmochitina minor Eisenack 1931
Pl. 1, Figs. A, D, E
Diagnosis: The vesicle is slightly longer than it is wide and possesses a collar around the aperture that flares outward in an oral direction. The vesicle wall may be smooth or ornamented with fine rugae.
Remarks: This species is widespread in the Ordovician of Europe. In North America, it has been reported from the Viola and Fernvale Limestones and lower part of the Sylvan Shale (D. complanatus Zone) in Oklahoma by Jenkins (1969, 1970) and from Upper Ordovician rocks in Ohio, Indiana, and Kentucky by Miller (1976). The Maquoketa specimens, like the Sylvan material, occur in chains more often than as isolated vesicles. In many specimens, the operculum is still attached and covers the aperture.
Occurrence: This species is common in samples 1 and 6 of the Maquoketa Shale.
Type species: Cymatiosphaera radiata O. Wetzel 1933 Cymatiosphaera sp.
Pl. 5, Figs. D, F
Diagnosis: The spherical vesicle is reticulate with membranous muri separating polygonal areas that have smooth floors.
Remarks: Only two specimens were found, both from sample 1 in the Maquoketa Shale. Unlike Dictyotidium Eisenack emend. Staplin 1961 (p. 417), the muri are high and are not the sites of echinate projections. The size of the polygonal fields is somewhat larger than those commonly found on Dictyotidium.
Type species: Leiosphaeridia baltica Eisenack 1958a
Leiosphaeridia tenuissima Eisenack 1958b
Pl. 3, Fig. G
Diagnosis: The vesicle is single-walled, smooth, with compressional folds that may be concentric and curved but are usually linear and sometimes overlapping and intersecting.
Remarks: The size range of the Maquoketa and Viola specimens is usually 100 to 200µ and is consistent with those of the Kope and Fairview Formations of Kentucky and Ohio (Jacobson, 1978).
Occurrence: This species is abundant in samples 1, 4, and 5 of the Maquoketa Shale and 9, 15, 16, and 17 of the Viola Limestone.
Type species: Actipilion druggii Loeblich 1970
Actipilion druggii Loeblich 1970
Pl. 3, Fig. D; Pl. 4, Figs. C, F
Diagnosis: This species apparently has a bilayered central body with the processes formed only from the outer layer, which accounts for their being delicate and thin. Processes do not communicate with the vesicle interior; they appear pitted and in places are granular.
Remarks: The length of the processes varies but all are thin- walled, almost membranous in appearance, wrinkled, and pitted. The processes are easily torn and separated from the vesicle, resulting in specimens that often have only a small number of the processes still attached to the vesicle. The Maquoketa specimens are within the range of morphologic variation shown by the type material from the Sylvan Shale in Oklahoma (Loeblich, 1970).
Occurrence: This species is not abundant. Several specimens were found in samples 1, 4, and 5 of the Maquoketa Shale.
Type species: Ammonidium microcladum (Downie) Lister 1970
Ammonidium sp.
Pl. 4, Figs. A, B
Diagnosis: The vesicle is circular, ornamented with hollow processes that are distally branched by a simple distal multifurcation with aculeate secondary tips. The surface of the wall is smooth.
Remarks: Only one specimen was found from the Maquoketa Shale. The closest comparable species is A. waldronensis described from the Silurian Waldron Shale from Indiana (Tappan and Loeblich, 1971). The Maquoketa specimens are 20µ in diameter with the processes being 8 to 10µ in length. A. waldronensis is slightly larger (20 by 34µ to 32 by 45µ).
Occurrence: This species is rare in sample 1 of the Maquoketa Shale.
Type species: Baltisphaeridium longispinosum (Eisenack) Eisenack 1959b
Baltisphaeridium hirsutoides (Eisenack) Eisenack 1959b
Pl. 2, Figs. D, E, F; Pl. 4, Figs. D, E, G
Diagnosis: The spherical vesicle is armed with hollow processes that are stiff with simple acuminate tips. The process length does not exceed vesicle diameter. The vesicle and processes are smooth to microgranulate.
Remarks: The Maquoketa specimens, which average 50µ in diameter, compare closely to those described from the Upper Ordovician of Ohio and Kentucky (Jacobson, 1978). As noted by Jacobson, B. hirsutoides is an intermediate morphologic form between B. longispinosum, with its longer processes, and B. multipilosum, with its numerous, short processes. B. hirsutoides is common and easy to separate from the more variable kinds of baltisphaerids. B. hirsutoides and B. parvigranosum appear to be conspecific. Baltisphaeridium parvulisidereum, from the Upper Ordovician Eden Shale, Indiana, differs from the aforementioned species by having large, widely spaced grana scattered about the process walls. It is our view that the style and not the density of ornamentation may be important in distinguishing one baltisphaerid from another in this complex, and that the density of grana is difficult to apply as a routine basis for separating species.
Occurrence: This species is common in samples 1, 2, 4, 5, and 6 of the Maquoketa Shale.
Baltisphaeridium perclarum Loeblich and Tappan 1978
Pl. 2, Figs. A, B, C
Diagnosis: The circular vesicle has long stout processes that are constricted at the proximal end and may possess small secondary branches. The vesicle wall surface is smooth, whereas the process wall is finely granulate.
Remarks: The Maquoketa specimens are remarkably similar in morphology to the Sylvan Shale type specimens described by Loeblich and Tappan (1978). The processes are constricted at their base and closed from the vesicle interior by a cup-shaped plug. Secondary branches of the main processes occur, but not on all specimens. Some of the secondary branches are more broadly based and larger than those figured as type specimens. Baltisphaeridium perclarum is similar to B. latiradiatum (Eisenack, 1959b) and B. constrictum Kjellstrom 1971 by possessing processes constricted at their base. In B. constrictum the vesicle is granulate and processes are smooth. B. latiradiatum has smooth processes and a shagrinate vesicle wall. Baltisphaeridium aff. B. perclarum from the Upper Ordovician White Head Formation of the Perce region, Gaspe Peninsula (Martin, 1980) may be conspecific with B. perclarum.
Baltisphaeridium sp. a
Pl. 4, Figs. H, I
Diagnosis: The vesicle is circular with long flexible processes that have acuminate to broadly rounded tips. The processes are not constricted at their bases and vary in width. The vesicle and process surface is smooth.
Remarks: Baltisphaeridium oligopsakium from the Sylvan Shale of Oklahoma (Loeblich and Tappan, 1978) and B. exilibrachium from the Eden Shale of Indiana (Colbath, 1979) both resemble this species in size, shape, number, and length of processes. The former species has grana and spinules on the processes and grana and pits on the vesicle and process walls with coarser grana on the processes.
Occurrence: This species occurs infrequently in sample 1 of the Maquoketa Shale.
Baltisphaeridium sp. b
Pl. 5, Figs. A, B, C
Diagnosis: The vesicle is circular, 40 to 50µ in diameter, and ornamented with short broad processes whose length is about 1/4 the vesicle diameter. The processes have broad-based spinules.
Remarks: Because only two specimens of this species were found and both are compressed, it is not possible to make a specific taxonomic identification nor propose a new species. Baltisphaeridium accinctum from the Bromide Formation (Loeblich and Tappan, 1978) most closely resembles the Maquoketa specimens. B. accinctum has a greater number of processes and grana scattered over the vesicle wall.
Occurrence: Two specimens were found in sample 1 of the Maquoketa Shale.
Baltisphaeridium cf. B. hirsutoides
Pl. 3, Fig. A, C
Diagnosis: The vesicle is subcircular to subpolygonal in outline, ornamented with long conical processes that appear to be plugged at their bases with no communication with the vesicle interior. Some of the processes appear to have secondary fillings. Processes have pointed tips. Process and vesicle wall is smooth.
Remarks: This species resembles B. hirsutoides except that it has a subpolygonal outline. Gyalorethium chondrodes from the Bromide Formation of Oklahoma (Loeblich and Tappan, 1978) has a subpolygonal outline similar to B. cf. B. hirsutoides, but the processes are hollow and communicate with the vesicle interior.
Occurrence: This species is rare in sample 1 of the Maquoketa Shale.
Type species: Cheleutochroa gymnobrachiata Loeblich and Tappan 1978
Cheleutochroa cf. C. gymnobrachiata
Pl. 5, Figs. E, G, H, I
Diagnosis: The circular vesicle has conical processes. The vesicle wall has a reticulate surface pattern that grades into ridges at the process base. These ridges then blend into the process, which has a smooth surface. The processes are simple or multibranched at their tips.
Remarks: The species, described by Loeblich and Tappan (1978) from the Sylvan Shale of Oklahoma, has unbranched processes with acuminate tips. The Maquoketa specimen possesses both simple and branched processes, indicating that branching may not be an important specific character. Acanthomorphic acritarchs (those with processes) are often characterized by heteromorphic processes at the specific level (e.g., Baltisphaeridium stamineum, Diexallophasis remota).
Occurrence: This species is rare in sample 1 of the Maquoketa Shale.
Type species: Diexallophasis remota (Deunff) emend. Playford 1977
Diexallophasis sp.
Pl. 6, Fig. C
Diagnosis: The vesicle is spherical to subspherical with 6 to 10 hollow processes that communicate freely with the vesicle interior. The processes are broad, unbranched, bifurcate, or multifurcate. The process and vesicle wall are sculptured with scattered grana and an occasional broad-based spine.
Remarks: The Maquoketa specimens are 20 to 25µ in diameter with the processes, in some cases, longer than the vesicle diameter. They most closely resemble Diexallophasis caperoradiola Loeblich 1970 from the Silurian Maplewood Shale of New York. The Maquoketa specimens have fewer grana and are not striated around the process bases; otherwise they are comparable in size and shape.
Occurrence: The species is common in samples 1, 4, and 6 of the Maquoketa Shale.
Type species: Eupoikilofusa striatifera (Cramer) Cramer 1971
Eupoikilofusa parvuligranosa Loeblich and Tappan 1978
Pl. 3, Fig. H
Diagnosis: The vesicle is thin-walled, fusiform, and ornamented with discontinuous ridges that do not extend to the polar tips. Small grana cover the vesicle wall and some are aligned in rows parallel to the long axis of the vesicle.
Remarks: The type specimens from the Sylvan Shale of Oklahoma (Loeblich and Tappan, 1978) have a larger number of smaller ridges than the Maquoketa specimen.
Occurrence: This species is represented by a single incomplete specimen from sample 5 of the Maquoketa Shale.
Type Species: Leiofusa fusiformis (Eisenack) Eisenack 1938
Leiofusa litotes Loeblich and Tappan 1978
Pl. 3, Fig. B
Diagnosis: The elongate vesicle has polar processes drawn out in acuminate tips with gradual thinning of the vesicle so no boundary exists between vesicle and process. The surface is smooth.
Occurrence: The species is rare in samples 1, 4, 5, and 6 of the Maquoketa Shale and 15 of the Viola Limestone.
Type species: Lophosphaeridium rarum Timofeev ex Downie 1963
Lophosphaeridium edenense Loeblich and Tappan 1978
Pl. 3, Fig. F
Diagnosis: The vesicle is circular in outline. The wall is thin and covered with large solid grana. The vesicle ranges from 18 to 30µ in diameter.
Remarks: Lophosphaeridium edenense from the Maquoketa and Viola is smaller than the specimens from the Eden Formation of Indiana (Loeblich and Tappan, 1978).
Occurrence: This species is comm n in samples 1, 4, and 5 of the Maquoketa Shale and 9, 15, 16, and 17 in the Viola Limestone.
Type species: Multiplicisphaeridium ramispinosum Staplin 1961
Multiplicisphaeridium bifurcatum Staplin, Jansonius, and Pocock 1965
Pl. 3, Fig. E; Pl. 6, Fig. E
Diagnosis: A species of Multiplicisphaeridium with homomorphic processes that are conical with simple or bifurcate tips. The vesicle wall is smooth.
Remarks: Multiplicisphaeridium bifurcatum is an easily recognized form having a small vesicle diameter (±20p) and conical processes with bifurcate tips. The type specimens are reported from Middle Ordovician rocks of Anticosti Island, Nova Scotia (Staplin and others, 1965) and are larger (25 to 35µ) than the Maquoketa specimens that agree more in size with those of the Upper Ordovician, Trenton Formation, Anticosti Island (Loeblich and Tappan, 1978), and the Upper Ordovician White Head Formation, Gaspe Peninsula, Quebec (Martin, 1980).
Occurrence: This species is common in samples 1, 2, 4, and 5 of the Maquoketa Shale.
Multiplicisphaeridium irregulare Staplin, Jansonius, and Pocock 1965
Pl. 6, Figs. D, F, H
Diagnosis: The semi-spherical vesicle has broadly based hollow processes that are multibranched in an irregular manner. Some specimens have heteromorphic processes that are both bifurcate and multibranched. Vesicle and process surface is smooth.
Remarks: The Maquoketa specimens have a range of variability in process branching that includes forms that are more frequently multibranched than those illustrated by Loeblich and Tappan (1978) from the Upper Ordovician, Trenton Formation, Anticosti Island, Quebec, for the type material (Staplin and others, 1965). M. irregulare from the Upper Ordovician White Head Formation of the Perce region, Gaspe Peninsula, Quebec (Martin, 1980) resembles the Maquoketa specimens more closely in the degree of process branching.
Occurrence: This species is common in samples 1, 2, 4, 5, and 6 of the Maquoketa Shale.
?Multiplicisphaeridium sp. a
Pl. 6, Fig. A
Diagnosis: The spherical vesicle (15µ in diameter) has at least 11 broad processes with cylindrical shafts that are multifurcate on the distal ends. The processes are as long as the vesicle is wide; process and vesicle wall is smooth.
Remarks: Only one specimen was found in an SEM mounting. Because of the opacity of this specimen, it cannot be determined whether the processes communicate with the vesicle interior; therefore, placing this specimen in the genus Multiplicisphaeridium is tentative. Baltisphaeridium pilaris, from the Silurian, has a process branching pattern reminiscent of ?Multiplicisphaeridium sp. a, but it is a larger form with more slender processes.
Occurrence: One specimen was found in sample 1 of the Maquoketa Shale.
Multiplicisphaeridium sp. b
Pl. 6, Fig. B
Diagnosis: The small (18µ in diameter) subspherical vesicle is differentiated from the processes, which are broadly based, conical, with rounded or bifurcated tips. Over 30 processes are visible in equatorial view. The surface of the vesicle and processes is smooth.
Remarks: Multiplicisphaeridium martae from the Middle Cambrian of Spain (Cramer and Diez, 1972) resembles the Maquoketa specimens.
Occurrence: This species is rare in sample 1 of the Maquoketa Shale.
Type species: Polygonium gracilis Vavrdova 1966
Polygonium polyacanthum (Eisenack 1965) comb. nov.
Pl. 3, Figs. K, L; Pl. 7, Figs. A-D
Diagnosis: The vesicle outline is polygonal to subcircular. Hollow processes communicate with vesicle interior. The process tips are simple, acuminate. The vesicle wall is smooth, whereas the process wall is granulate to echinate.
Remarks: The sculpture of P. polyacanthum makes this form easy to separate from P. polygonale, which has a smooth vesicle wall and processes. This distinction warrants raising P. polygonale polyacanthum (Eisenack, 1965) to the specific rank (see comments by Jacobson, 1978). Polygonium celestum reported from the Upper Ordovician Eden Shale of Indiana (Colbath, 1979) is considered here to be a synonym of P. polyacanthum. As reported by Colbath, the Eden Shale specimens possess "baculae" that are apparent on the process walls. Polygonium uncinatum (Downie) Martin 1966 appears to have larger echinate projections on the processes than P. polyacanthum. Polygonium polyacanthum is reported from the Sylvan Shale, Oklahoma (Hedlund, 1960); Ordovician and Silurian erratics, Baltic Region (Eisenack, 1931, 1938, 1959a, 1963, 1965, 1968); subsurface, Lower Ordovician, Poland (Gorka, 1969); and Upper Ordovician, New York, Ohio, and Kentucky (Jacobson, 1978).
Occurrence: This species is abundant in samples 1, 4, 5, and 6 of the Maquoketa Shale.
Polygonium cf. P. polygonale
Pl. 7, Figs. E, F, H
Diagnosis: The vesicle is polygonal, has broadly based conical processes with acuminate tips. The surface sculpture is smooth to microgranulate.
Remarks: Polygonium cf. P. polygonale from the Upper Ordovician of Kentucky and New York (Jacobson, 1978), as well as the material from the Middle Ordovician through Silurian rocks in Europe, are larger than the Maquoketa specimens, and there is some doubt as to how closely the Maquoketa specimens relate to these previously described species.
Occurrence: This species is abundant in samples 1, 4, and 6 of the Maquoketa Shale.
Polygonium sp. a
Pl. 7, Fig. G
Diagnosis: A form of Polygonium with a distinct triangular outline and very broad-based processes; vesicle and process wall smooth.
Occurrence: This species is rare in sample 1 of Maquoketa Shale.
Polygonium sp. b
Pl. 7, Fig. I
Diagnosis: The vesicle is polygonal to subcircular in outline (20µ in diameter) with more than 35 thin conical processes visible. The vesicle and process wall is smooth.
Occurrence: This species is rare in sample 1 of Maquoketa Shale.
Polygonium sp. c
Pl. 7, Fig. J
Diagnosis: The small (±15p) subcircular vesicle has more than 35 conical processes visible. The processes' length is 1/2 the vesicle diameter. The processes have acuminate tips. The vesicle and process wall is smooth.
Occurrence: This species is rare in sample 1 of Maquoketa Shale.
Type species: Veryhachium trisulcum (Deunff) Deunff 1959
Veryhachium cf. V. fictusistriatum
Pl. 8, Fig. E
Diagnosis: The triangular-shaped vesicle has slightly convex sides and three hollow processes that extend from the corners and communicate freely with the vesicle interior. The processes end in a blunt point and their length is equal to or less than the vesicle width. The surface of the vesicle and processes is smooth to finely granulate.
Remarks: Veryhachium fictusistriatum from the Upper Ordovician Eden Shale of Indiana (Colbath, 1979) is described as having a finely granulate wall with the grana appearing to be aligned in rows, giving the specimen a striate microstructure parallel to the vesicle sides. Not all the illustrations by Colbath show what could be interpreted as striate microstructure. The Maquoketa specimens have a size and shape similar to V. fictusistriatum and are certainly part of the same veryhachid group.
Occurrence: This species is common in samples 1 and 4 of the Maquoketa Shale, rare in sample 14 of the Viola Limestone.
Veryhachium hamii Loeblich 1970
Pl. 3, Figs. J, N; Pl. 6, Figs. G, I; Pl. 8, Fig. G
Diagnosis: This is a species of Veryhachium with a triangular-shaped central body, which is commonly inflated, with three long, hollow processes in the plane of the central body and with secondary processes arising from the face of the central body. The surface is smooth to microgranulate.
Remarks: Loeblich (1970) reported this species from the Sylvan Shale in Oklahoma. The basic body shape is a triangle with rounded apices, each possessing a process that communicates freely with the vesicle interior. What gives this species a varied appearance is the existence of secondary processes that vary in size and arise from the face of the vesicle and extend out from the vesicle face at right angles to the main processes. Some speci- mens are almost pyraform in shape, reminiscent of Tectitheca. Compaction of this form results in a wide array of shapes as illustrated here.
Occurrence: This species is rare in samples 1, 3, and 6 of the Maquoketa Shale and common in samples 9-11 and 13-17 of the Viola Limestone.
Veryhachium oklahomense Loeblich 1970
Pl. 3, Fig. M; Pl. 8, Figs. D, F
Diagnosis: The rectangular body is nearly square in outline. The sides are nearly straight. Four long flexible processes are presented. The vesicle and process wall is smooth.
Remarks: The length of the processes varies but they are usually long and thin, extending from a rectangular vesicle. In most specimens, the length of the processes is greater than the width of the vesicle. Representatives of this species are usually small with the vesicle being less than 25µ in diameter. In some specimens the sides of the vesicle are slightly concave. Loeblich (1970) has reported (from Sylvan Shale specimens) the rare occurrence of a fifth process that arises on the face of the vesicle, a feature not observed in the Maquoketa specimens.
Occurrence: This species is common in samples 1, 3, and 5 of the Maquoketa Shale.
Type species: Villosacapsula setosapellicula (Loeblich) Loeblich and Tappan 1976
Villosacapsula setosapellicula (Loeblich) Loeblich and Tappan 1976
Pl. 3, Fig. I; Pl. 8, Figs. A, B, C
Diagnosis: The triangular-shaped vesicle has an inflated appearance, three processes that are relatively short and pointed at their ends. The vesicle and process wall is ornamented with short, scattered microspines.
Remarks: This species is characterized by the short spines, appearing as grana in transmitted light, distributed evenly over the vesicle and process wall. Although not mentioned in the original description, the small spines may terminate in an expanded blunt tip resembling a mushroom head. Villosacapsula setosapellicula is also reported from the Upper Ordovician Eden Shale of Indiana (Colbath, 1979) and the Sylvan Shale of Oklahoma (Loeblich, 1970).
Occurrence: This species is common in samples 1, 3, and 4 of the Maquoketa Shale; rare in samples 9 and 15 of the Viola Limestone.
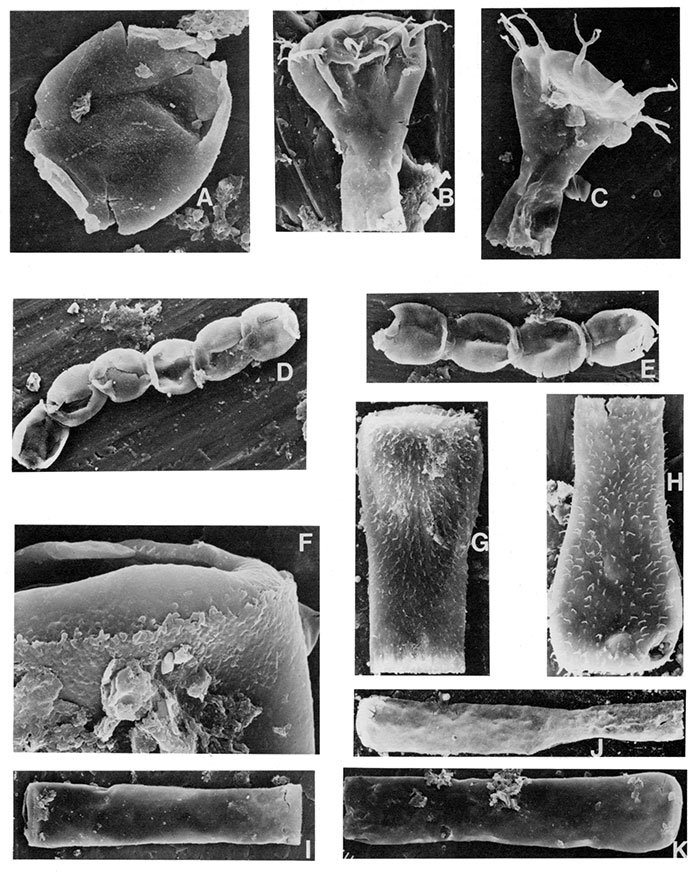
| Figure | |
|---|---|
| B,C | Ancyrochitina merga Jenkins 1970, both specimens X600 and about 90µ in length. |
| F,I,J,K | Conochitina elegans Eisenack 1931, I, note the smooth vesicle wall, small spines around the oral edge (enlarged in Fig. F X2000), X240, 375µ in length; J and K X300. |
| G,H | Conochitina wesenbergensis Eisenack 1959a, note the near-parallel alignment of the surface spines in rows parallel to the long direction of the vesicle, both specimens X440 and about 170µ in length. |
| A,D,E | Desmochitina minor Eisenack 1931, A, a single vesicle with the operculum still preserved, X660; D and E, chains of individual vesicles, note how the oral lip and aperture of one specimen is oppressed to the aboral surface of another, both chains X260. |
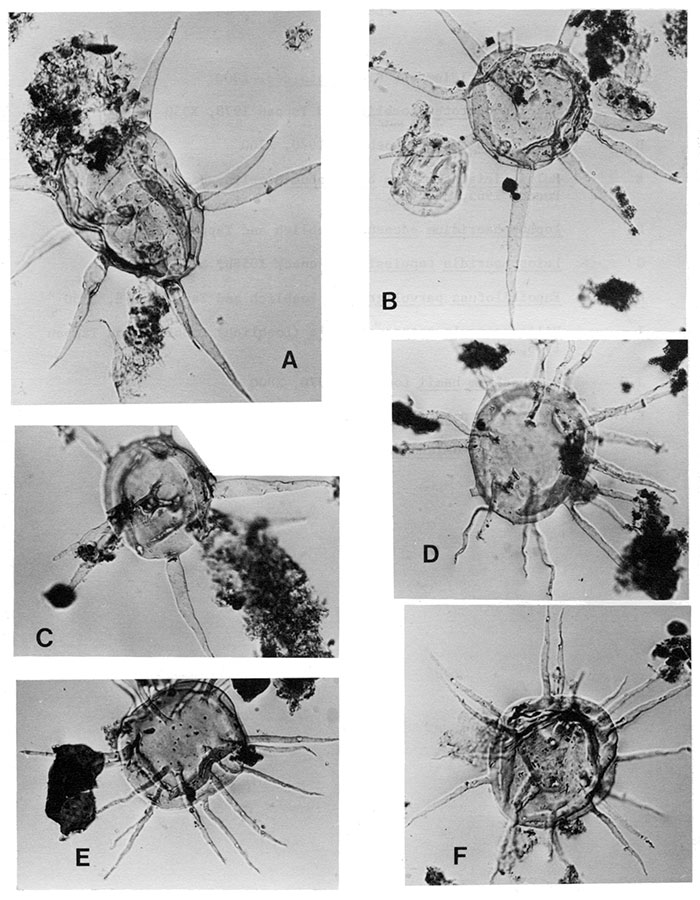
| Figure | |
|---|---|
| A,B,C | Baltisphaeridium perclarum Loeblich and Tappan 1978, note the constriction of the processes at their proximal end, all X450. |
| D,E,F | Baltisphaeridium hirsutoides Eisenack 1959b, all X500. |
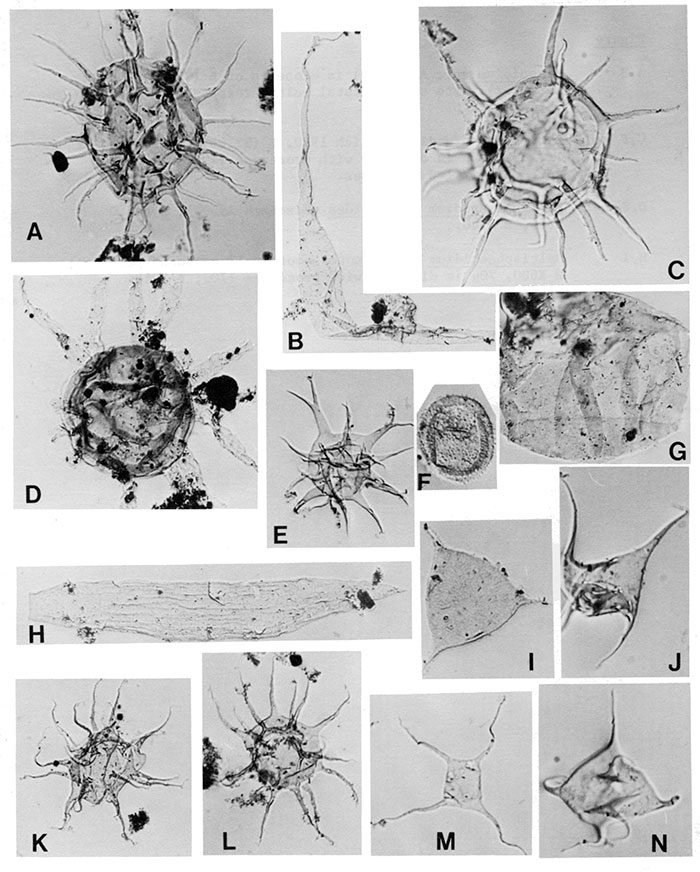
| Figure | |
|---|---|
| A,C | Baltisphaeridium cf. B. hirsutoides, X800 |
| B | Leiofusa litotes Loeblich and Tappan 1978, X350 |
| D | Actipilion druggii Loeblich 1970, X700 |
| E | Multiplicisphaeridium bifurcatum Staplin, Jansonius, and Pocock 1965, X800 |
| F | Lophosphaeridium edenense Loeblich and Tappan 1978, X800 |
| G | Leiosphaeridia tenuissima Eisenack 1958b, X800 |
| H | Eupoikilofusa parvuligranosa Loeblich and Tappan 1978, X350 |
| I | Villosacapsula setosapellicula (Loeblich) Loeblich and Tappan 1976, X800 |
| J,N | Veryhachium hamii Loeblich 1970, X800 |
| K,L | Polygonium polyacanthum Eisenack 1965, X800 |
| m | Veryhachium oklahomense Loeblich 1970, X800 |
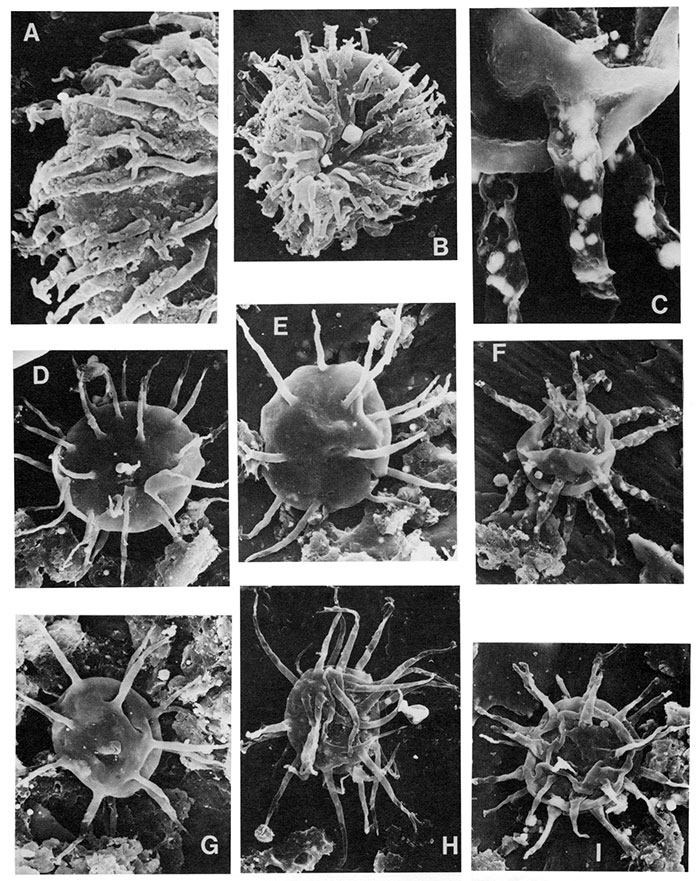
| Figure | |
|---|---|
| A,B | Anmonidium sp., A (X6600) is close-up of B (X3000, 20µ in diameter), note simple distal multifurcation with aculeate secondary tips. |
| C,F | Actipilion druggii Loeblich 1970, C (X2200) is close-up of F (X600, 70µ in diameter with processes ±40µ in length), note thin-walled processes. |
| D,E,G | Baltisphaeridium hirsutoides (Eisenack 1959b), D and G X660, E X940. |
| H,I | Baltisphaeridium sp. a, note smooth wall on both specimens; H X600, 70µ in diameter with processes ±70p; I X940. |
| Figure | |
|---|---|
| A,B,C | Baltisphaeridium sp. b, A is X1300 and B is close-up of A (X3200) showing scalloped appearance of processes caused by presence of broadly based spinules, C X1500. |
| D,F | Cymatiosphaera sp., D X1800 (30µ in diameter), E X1600. |
| E,G,H,I | Cheleutochroa cf. C. gymnobrachiata, E is Y,1100 (30µ in diameter with process ±25µ in length), note simple and branched spines; G is close-up of E at X6000, note reticulate pattern on vesicle wall, ribs at base of processes, and smooth process wall above basal ribs; H X1300; I is close-up of H X6600. |
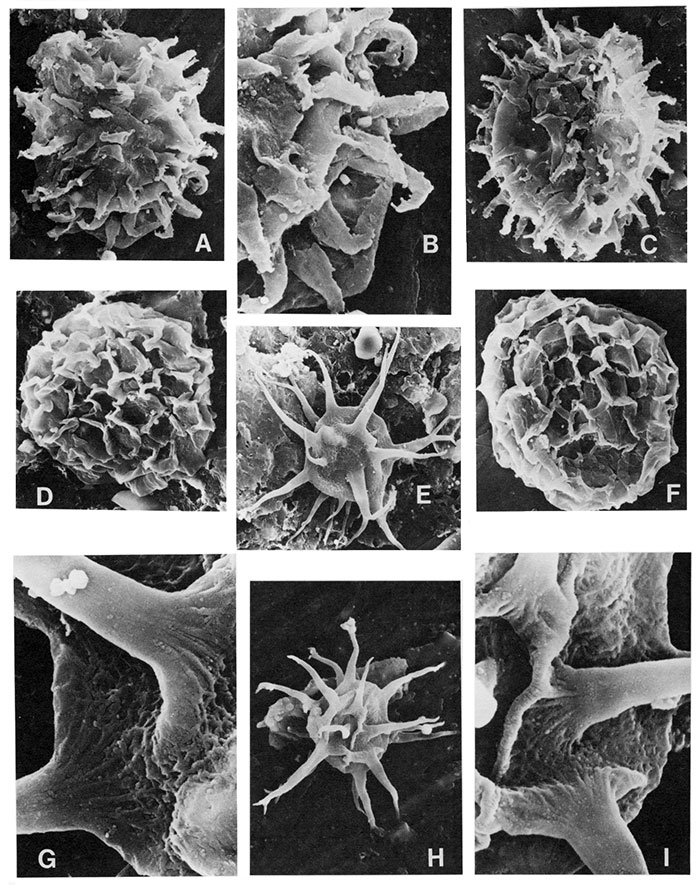
| Figure | |
|---|---|
| A | ?Multiplicisphaeridium sp. a, X2400 |
| B | Multiplicisphaeridium sp. b, X3600 (15µ in diameter) |
| C | Diexallophasis sp., X860 |
| D,F,H | Multiplicisphaeridium irregulare Staplin, Jansonius, and Pocock 1965, D, note the multifurcate processes, X1300; F X1600; H X1300 |
| E | Multiplicisphaeridium bifurcatum Staplin, Jansonius, and Pocock 1965, note the simple and bifurcate processes, X1300 |
| G,I | Veryhachium hamii Loeblich 1970, G X1300; I X2200 |
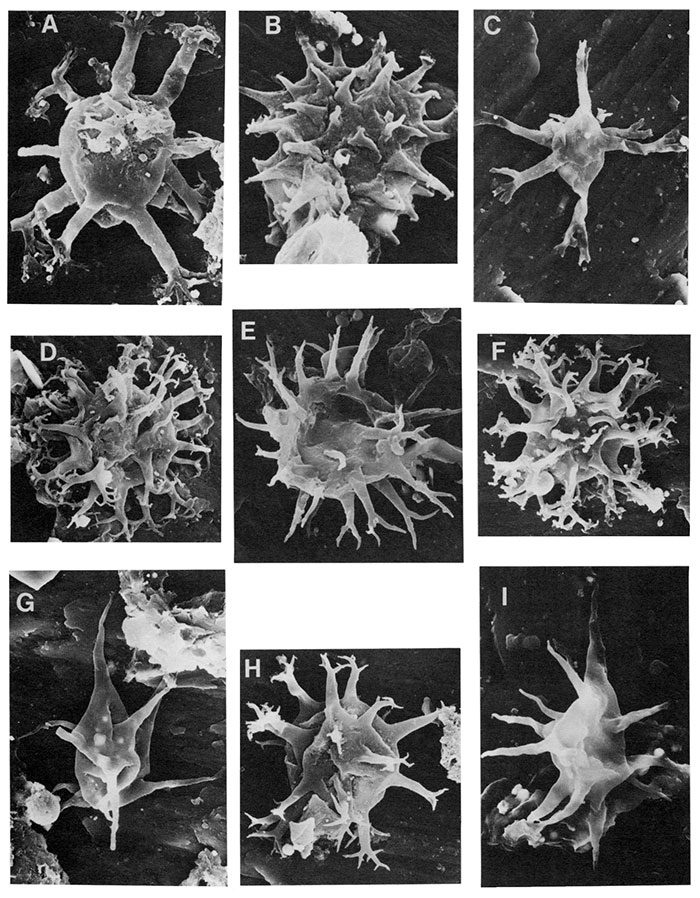
| Figure | |
|---|---|
| A-D | Polygonium polyacanthum (Eisenack 1965), comb. nov., A is enlargement (X4800) of B (X1100), note echinate projections on process wall and process base; C is enlargement (X4800) of D (X1200). Specimens are 20 to 30µ in diameter with process length often equaling or greater than vesicle diameter. |
| E,F,H | Polygonium cf. P. polygonale, E is enlargement (X4400) of F (Xl200, 25µ in diameter), note absence of echinate projections on processes; H X1000. |
| G | Polygonium sp. a, X1200, vesicle diameter ±30µ, note triangular outline of vesicle. |
| I | Polygonium sp. b, X1300, note the many thin conical processes, compare to G. |
| J | Polygonium sp. c, X2600. |
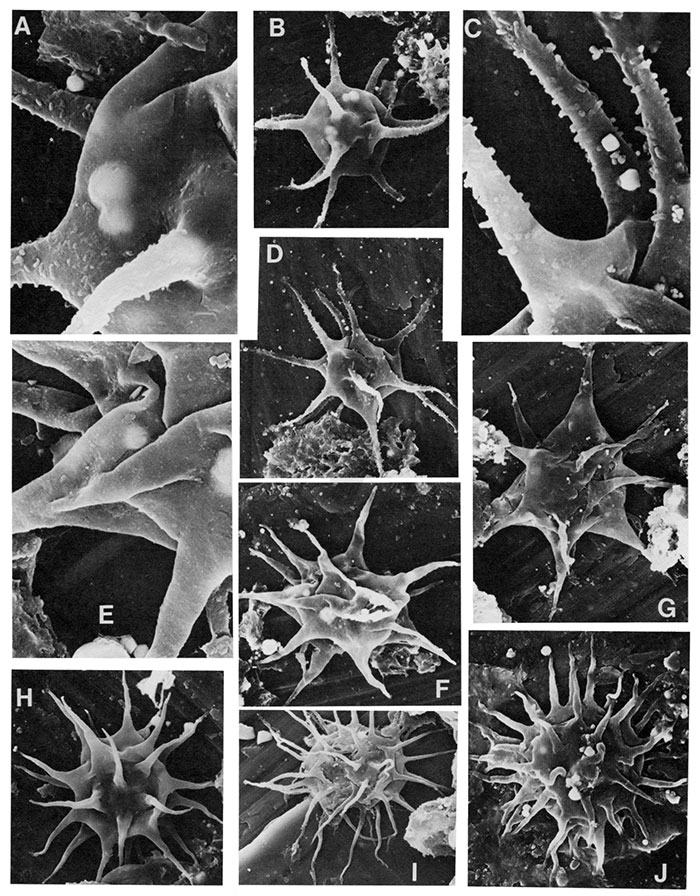
| Figure | |
|---|---|
| A,B,C | Villosacapsula setosapellicula (Loeblich) Loeblich and Tappan 1976, A X1200; B (X4000) is enlargement of A, note spines on vesicle wall; C X1500. |
| D,F | Veryhachium oklahomense Loeblich 1970, D X1500 (18µ in diameter, processes µ30µ long); F X1100. |
| E | Veryhachium cf. V. fictusistriatum, X1800, 20µ in width, processes ±20µ in length. |
| G | Veryhachium hamii Loeblich 1970, stereo-pair, X2000. |
Special thanks are due to Jocelyn A. Legault (University of Waterloo), and Stephen R. Jacobson (Chevron U.S.A., Inc.) for their helpful discussions concerning the systematics of acritarchs and chitinozoans. Jack Etter (Cities Service Company) read and offered constructive comments about the manuscript. Maggie Draughon (Cities Service Company) typed several versions of this report and Allen White (Cities Service Company) took the excellent scanning electron micrographs of the fossils. Permission to publish this report was kindly granted by Cities Service Company.
Achab, Aicha, 1977, Les chitinozoaires de la zone a Dicellagraptus complanatus Formation de Vaureal, Ordovicien superieur, Ile d'Anticosti, Quebec [The chitinozoans of the Dicellagraptus complanatus zone, Vaureal Formation, Upper Ordovician, Anticosti Island, Quebec]: Canadian Journal of Earth Sciences, v. 14, p. 413-425.
Boalch, G.T., and Parke, M., 1971, The Prasinophycean genera (Chlorophyta) possibly related to fossil genera, in particular the genus Tasmanites: Proceedings of the II Planktonic Conference Roma 1970, v. 1, p. 9 4-107.
Colbath, G. Kent, 1979, Organic-walled microphytoplankton from the Eden Shale (Upper Ordovician), Indiana, U.S.A.: Palaeontographica, Abteilung B, v. 171, p. 1-38.
Collinson, Charles, and Schwalb, H., 1955, North American Paleozoic Chitinozoa: Illinois State Geological Survey Report of Investigations 186, p. 1-33.
Combaz, A., Calandra, F., Jansonius, J., Millepied, P., Poumot, C., and Van Oyen, F.H., 1967, Microfossiles organiques du Paleozoique 2, Les chitinozoaires, Morphographie [Organic microfossils of the Paleozoic 2, The Chitinozoa, Morphography): Paris Centre National de la Recherche Scientifique, 43 p.
Cramer, F.H., 1971, Distribution of selected Silurian acritarchs: An account of the palynostratigraphy and paleogeography of selected Silurian acritarch taxa: Revista Espanola de Micropaleontologia, Numero Extraordinario, 203 p.
Cramer, F.H., and Diez, M. d. C.R., 1972, Acritarchs from the upper middle Cambrian Oville Formation of Leon, northwestern Spain: Revista Espanola de Micropaleontologia, Numero Extraordinario, XXX Aniversario, Empressa Nacional Adaro, Diciembre 1972, p. 3950.
Deflandre, G., 1954, Systematique des hystrichosphaerides: sur l'accep- tion de genre Cymatiosphaera O. Wetzel [Systematics of hystricho- spheres: with regard to the genus Cymatiosphaera O. Wetzel]: Societe Geologique de France Compte Rendu Sommaire des Seances, 1954/12, p. 257-258.
Deunff, Jean, 1959, Microorgaismes planctonique du primaire armoricain, 1. - Ordovicien du Veryhach (Presquile de Crozon) [Planctonic microorganisms of the armoricain primary, 1. - Ordovician Veryhachium (Presquile de Crozon)]: Bulletin Societe Geologique et Mineralogique de Bretagne, no. 2, p. 1-41.
Downie, Charles, 1959, Ilystrichospheres from the Silurian Wenlock Shale of England: Palaeontology, v. 2, p. 56-71.
Downie, Charles, Evitt, W.R., and Sarjeant, W.A.S., 1963, Dinoflagellates, hystrichospheres, and the classification of the acritarchs: Stanford University Publication, Geological Sciences, v. 7, p. 1-16.
Eisenack, Alfred, 1931, Neue Mikrofossilien des baltischen Silurs I [New microfossils of the Baltic Silurian]: Palaeontologische Zeitschrift, v. 12, p. 74-118.
Eisenack, Alfred, 1938, Hystrichosphaerideen und verwandte Formen im baltischen Silur [Hystrichospheres and related forms in the Baltic Silurian]: Zeitschrift Geschiebeforschung, v. 14, p. 1-30.
Eisenack, Alfred, 1951, Uber Hystrichosphaerideen und andere kleinformen aus baltischen Silur und Kambrium (About hystrichospheres and other microfossils from the Baltic Silurian and Cambrian]: Senkenbergiana Lethaea, v. 32, p. 1-4, 187-204.
Eisenack, Alfred, 1955, Chitinozoen, Hystrichospharen und andere Mikrofossilien aus dem Beyrichia-Kalk [Chitinozoa, hystrichospheres, and other microfossils from the Beyrichian-chalk]: Senckenbergiana Lethaea, v. 36, p. 157-188.
Eisenack, Alfred, 1958a, Tasmanites Newton 1875 und Leiosphaeridia n.g. als Gattungen der Ilystrichosphaeridea [Tasmanites Newton 1875 and Leiosphaeridia n.g. as Genera of Hystrichospheres]: Palaeontographica Abteilung A, v. 110, p. 1-19.
Eisenack, Alfred, 1958b, Mikrofossilien aus dem Ordovizium des Baltikums. 1. Markasitschist, Dictyonema-Schiefer, Glaukonitsand, Glaukonitkalk [Microfossils of the Baltic Ordovician. 1. Marcasite schist, Dictyonema-shale, glauconite sand, glauconite chalk]: Senkenbergiana Lethaea, v. 39, p. 389-405.
Eisenack, Alfred, 1958c, Mikroplankton aus dem norddeutschen Apt nebst einigen Bemerkungen uber fossile Dinoflagellaten [Microplankton of the north German Aptian in addition to some remarks covering fossil Dinoflagellates]: Neues Jahrbuch fur Geologie und Palaeontologie Abhandlugen, v. 106, p. 383-422.
Eisenack, Alfred, 1959a, Neotypen baltischer Silur Chitinozoen und neue Arten [New types of Baltic Silurian Chitinozoa and new species]: Neues Jahrbuch fur Geologie und Palaeontologie Abhandlungen, v. 108, p. 1-20.
Eisenack, Alfred, 1959b, Neotypen baltischer Silur Hystrichospharen und neue Arten [New types of Baltic Silurian Hystrichospheres and new species]: Palaeontographica, A, v. 112, p. 193-211.
Eisenack, Alfred, 1963, Mitteilungen zur Biologie der Hystrichospharen und uber neue Arten [Information about the biology of hystrichospheres and about new species]: Neues Jahrbuch fur Geologie und Palaeontologie Abhandlungen, v. 118, p. 207-216.
Eisenack, Alfred, 1965, Die Mikrofauna der Osteekalke, 1, Chitinozoen, Hystrichospharen [The microfauna of Baltic chalks, 1, Chitinozoa, Hystrichosphaeral: Neues Jahrbuch fur Geologie und Palaeontologie Abhandlungen, v. 123, p. 115-148.
Eisenack, Alfred, 1968, Microfossilien eines Geschiebes der Borkholmer Stufe, baltisches Ordovizium, F2 [Microfossils of the Borkholmer Stufe boulders, Baltic Ordovician, F2]: Mitteilung mineralgeologie Stattsinst., Hamburg, v. 38, p. 81-94.
Evitt, W.R., 1961, Observations on the morphology of fossil dinoflagellates: Micropaleontology, v. 7, p. 385-420.
Evitt, W.R., 1963, A discussion and proposals concerning fossil Dinoflagellates, Hystrichospheres, and Acritarchs, I, II: National Academy of Science, Proclamation 49, p. 158-164, 298-302.
Gorka, Hanna, 1969, Microorganismes de l'Ordovicien de Pologne [Microorganisms of the Ordovician of Poland]: Acta Palaeontologica Polonica, v. 22, 102 p.
Hedlund, R.W., 1960, Microfossils of the Sylvan Shale (Ordovician) of Oklahoma: M.S. Dissertation, University of Oklahoma, 90 p.
Jacobson, S.R., 1978, Acritarchs from Middle and Upper Ordovician rocks in New York State and the Cincinnati region in Ohio and Kentucky: Ph.D. Dissertation, Ohio State University, 303 p.
Jacobson, S.R., 1979, Acritarchs as paleoenvironmental indicators in Middle and Upper Ordovician rocks from Kentucky, Ohio and New York: Journal of Paleontology, v. 53, p. 1197-1212.
Jenkins, W.A.M., 1969, Chitinozoa from the Ordovician Viola and Fernvale Limestones of the Arbuckle Mountains, Oklahoma: Palaeontology, Special Paper 5, 44 p.
Jenkins, W.A.M., 1970, Chitinozoa: Geoscience and Man, I, p. 1-24.
Jenkins, W.A.M., and Legault, J.A., 1979, Stratigraphic ranges of selected Chitinozoa: Palynology, v. 3, p. 235-264.
Kjellstrom, Goran, 1971, Ordovician microplankton (baltisphaerids) from the Grotlingbo borehole No. 1 in Gotland, Sweden: Sveriges Geologiska Undersokning, Arsbok, v. 65, 75 p.
Klement, K.W., 1960, Dinoflagellaten und Hystrichospheres Permo-Triasiques d'Europe et d'Afrique [Permo-Triassic Dinoflagellates and Hystrichospheres of Europe and Africa]: Revue de Micropaleontologie, no. 3, p. 207-212.
Kozlowski, R., 1963, Sur la nature des chitinozoaires [On the nature of chitinozoans]: Acta Palaeontologia Polonica, v. 8, p. 425-449.
Laufeld, Sven, 1974, Silurian Chitinozoa from Gotland: Fossils and Strata, v. 5, 130 p.
Lee, W., 1956, Stratigraphy and structural development of the Salina Basin area: Kansas Geological Survey Bulletin 121, 167 p. [available online]
Lister, T.R., 1970, The acritarchs and chitinozoa from the Wenlock and Ludlow series of the Ludlow and Millichope areas, Shropshire, Part 1: Palaeontographical Society, Monographs, v. 124, 100 p.
Loeblich, A.R., Jr., 1970, Morphology, ultrastructure and distribution of Paleozoic acritarchs: Proceedings of the North American Paleontological Convention, Chicago, 1969, Part G, p. 705-788.
Loeblich, A.R., Jr., and Tappan, Helen, 1976, Some new and revised organic-walled phytoplankton microfossil genera: Journal of Paleontology, v. 50, p. 301-308.
Loeblich, A.R., Jr., and Tappan, Helen, 1978, Some Middle and Late Ordovician microphytoplankton from central North America: Journal of Paleontology, v. 52, p. 1233-1287.
Madler, K., 1963, Die figurierten organischen Bestandteile der Posidonienschiefer [The figured organic matter of the Posidonian-shale]: Beihefte zum Geologischen Jahrbuch, v. 58, p. 287-406.
Martin, Francine, 1966, Les acritarchs de Sart-Bernard (Ordovicien belge) [The acritarchs of Sart-Bernard (Belgium Ordovician)]: Bulletin de la Societe Belge de Geologie de Paleontologie et d'Hydrologie, v. 74, p. 423-444.
Martin, Francine, 1980, Quelques Chitinozoaires et Acritarches ordoviciens superieurs de la Formation de White Head en Gaspesie, Quebec [Some upper Ordovician chitinozoa and acritarchs of the White Head Formation in Gaspe, Quebec]: Canadian Journal of Earth Sciences, v. 17, p. 107-119.
Merriam, D.F., 1963, The geologic history of Kansas: Kansas Geological Survey Bulletin 162, 317 p. [available online]
Miller, Merrell, 1976, Maysvillian (Upper Ordovician) Chitinozoans from the Cincinnati Region of Ohio, Indiana and Kentucky: M.S. Dissertation, Ohio State University, 251 p.
Playford, Geoffrey, 1977, Lower and Middle Devonian acritarchs of the Moose River Basin, Ontario: Geological Survey of Canada, Bulletin 279, 87 p.
Rauscher, Raymond, 1973, Recherches micropaleontologiques et stratigraphiques dans l'Ordovicien et le Silurien en France [Micropaleontological and stratigraphic investigations of the Ordovician and Silurian of France]: Sciences Geologiques, Memoire 38, 224 p.
Staplin, F.L., 1961, Reef controlled distribution of Devonian microplankton in Alberta: Palaeontology, v. 4, p. 392-424.
Staplin, F.L., Jansonius, J., and Pocock, S.A.J., 1965, Evaluation of some acritarchous hystrichosphere genera: Neues Jahrbuch Geologie und Palaeontologie Abhandlungen, v. 123, p. 167-201.
Tappan, Helen, and Loeblich, A.R., Jr., 1971, Surface sculpture of the wall in Lower Paleozoic acritarchs: Micropaleontology, v. 17, p. 385-410.
Taugourdeau, Phillipe, 1965, Chitinozoaires de l'Ordovicien des U.S.A; comparaison avec les faunes de l'ancien monde [Ordovician Chitinozoa of the U.S.A., comparison with the faunas of the old world]: Revue de l'Institute Francais du Petrole, v. 20, p. 463-485.
Vavrdova, M., 1966, Paleozoic microplankton from central Bohemia: Casopis pro Mineralogie a Geologii, v. 11, p. 409-414.
Wall, David, 1965, Microplankton, pollen, and spores from the Lower Jurassic in Britain: Micropaleontology, v. 11, p. 151-190.
Wetzel, Otto, 1933, Die in organischer Substanz erhaltenen Microfossilien des baltischen Kreide-Feuersteins [The preserved microfossils in organic matter in the Baltic Cretaceous flints]: Palaeontographica Abteilung A, 77-78, p. 141-186.
Willman, H.B., and Buschbach, T.C., 1975, Ordovician System in Handbook of Illinois Stratigraphy: Illinois Geological Survey Bulletin 95, 261 p.
Kansas Geological Survey, Subsurface Ordovician of Northeastern Kansas
Placed on web Sept. 16, 2011; originally published in 1991.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/Sub4/index.html