Kansas Geological Survey, Bulletin 47, pt. 1, originally published in 1943

Originally published in 1943 as Kansas Geological Survey Bulletin 47, pt. 1. This is, in general, the original text as published. The information has not been updated. An Acrobat PDF version (7.4 MB) is also available.
The following Permian forms are described as new species referred to genera previously established: Heliospongia vokesi, Guadalupia williamsi, and Girtyocoelia dunbari. The following Permian genera and species are described as new: Laubenfelsia regularis, Pseudopemmatites skinneri, Defordia defuncta, Stylopegma dulce, S. conica, Polyphymaspongia explanata, and Talpaspongia clavata. All the Permian forms are calcisponges and tetracladine silicisponges. One new Pennsylvania calcisponge, Wewokella contorta, is referred to an established genus, and one, Cystauletes mammilosus, is assigned to a new genus. A new genus and species of Mississippian calcisponge is described as Cotyliscus ewersi.
The only comprehensive work on Permian sponges of North America is Girty's monographic treatment of the Guadalupian fauna (1908), in which are described sponges from the Capitan limestone only. The sponges of the Lower Permian beds are more abundant, more varied, and better preserved, and some of those of the Wolfcampian and Leonardian series are likely to be found in other areas, notably Kansas and Oklahoma, where Capitan equivalents are unrepresented. It is the purpose of this report to make known and to name at least the common, easily recognized forms, in order that they can be differentiated and their geographic and stratigraphic distribution can be observed and recorded.
Three Carboniferous sponges are included to supplement the somewhat more extensive records of those forms. (Girty, 1908a, 1911, 1915; King, 1933, 1938.)
The Permian sponges herein described are about equally divided between the calcisponges and the tetracladine lithistid sponges, other groups being so far unrepresented. The unusual feature of the assemblage is the relative abundance of the Tetracladina. The Permian sponges of Timor (Gerth, 1929) are principally lithistids belonging to suborders other than Tetracladina. No tetracladine sponges have been described from India or other parts of continental Asia, although Permian rocks are widely distributed in India, Indo-China, China, and Japan. The Permian rocks of Sicily may contain numerous tetracladine sponges, but the only important work on the sponges of that area (Parona, 1933) is inaccessible and is not entirely reliable, inasmuch as at least three species are defintiely known to be referred to the wrong genera. In any event, the Tetracladina probably do not constitute an important element of the sponge fauna of the Permian rocks of Sicily.
I am very grateful to Harold E. Vokes, of the American Museum of Natural History, and to Carl O. Dunbar, of Peabody Museum of Natural History, Yale University, for opportunity to study the Permian sponges in the collections of those institutions; to Raymond C. Moore, Director of the Kansas Geological Survey, for criticism and assistance in preparation of the manuscript and illustrations and for providing an opportunity to study the Permian faunas at first hand in the Glass Mountains area; to Ronald K. DeFord, for his encouragement and for help in obtaining loan of specimens; and to Robert E. King, John E. Adams, and John Skinner, for advice and guidance to collecting localities in the Glass Mountains. Preparation of the illustrations was aided by a grant from the Graduate Research Fund of the University of Kansas. To all these persons and to the institutions that they represent I extend my sincere thanks. Their courtesy and aid are appreciated.
The detailed classification of living sponges proposed by Laubenfels (1936) seems to require, for its application to fossil forms, knowledge of some structural details that are not commonly preserved. Most fossil sponges can at least be referred to the proper main division, however; and, because these main divisions (that is, the classes Demospongia, Calcispongia, and Hyalospongia) have special significance in the study of paleoecology, they are employed here. Minor subdivisions are designated by the terms of older classifications.
In Laubenfel's classification, the living lithistids are divided among most of the orders of the Demospongia, but the principal basis for such division is the character of the microscleres, which are not observable in most fossil forms. It is recognized that the massive (lithistid) mode of growth is common to many orders, and is of little classificatory significance.
According to Zittel, the tetracladine sponges range from Cambrian to Recent, but, as previously stated, living forms are divided on a somewhat different basis. In general, the spicules of the earlier forms seem to be relatively simple and scarcely differentiated; whereas, those of later forms are more complex and are differentiated into several kinds. Although I have studied somewhat better material than any that was available to Girty, and have had the benefit of his observations, I, like Girty (1908, pp. 71-72), hesitate to attempt to place the various genera in families, because of doubt as to what should constitute a family.
This genus comprises large, straight, bent, or branching subcylindrical sponges that are traversed throughout their length by a relatively large central cloaca. The surface is penetrated by pores (incurrent apertures) but not by ostia (excurrent apertures). The canals are approximately radial. The spicules are so oriented as to form three patterns. In transverse sections one of these patterns consists of concentric circles and another consists of radial lines. In longitudinal sections the radial pattern is seen to bend downward in approaching the exterior of the sponge body. The third pattern consists of lines almost parallel to the axis of the sponge, bending slightly outward in approaching the top of the sponge. Girty (1908a) interpreted these patterns as indicating hexactine spicules, so oriented that the rays of any spicule are in contact with the corresponding rays of the adjacent spicules at their tips. Opposing this view, I am convinced that the spicules are tetractine rather than hexactine. In the specimens studied by Girty the silica had been replaced by calcite, so that individual spicules were not discernible. I have examined specimens in which the spicules have been cemented by secondary silica but from which the limestone matrix has been removed by hydrochloric acid. The spicules seem to be typical tetracladine spicules of average size.
Genotype—Heliospongia ramosa Girty, 1908
Heliospongia vokesi, n. sp.
Plate 1, Figures 1, 2; Text Figure 1.
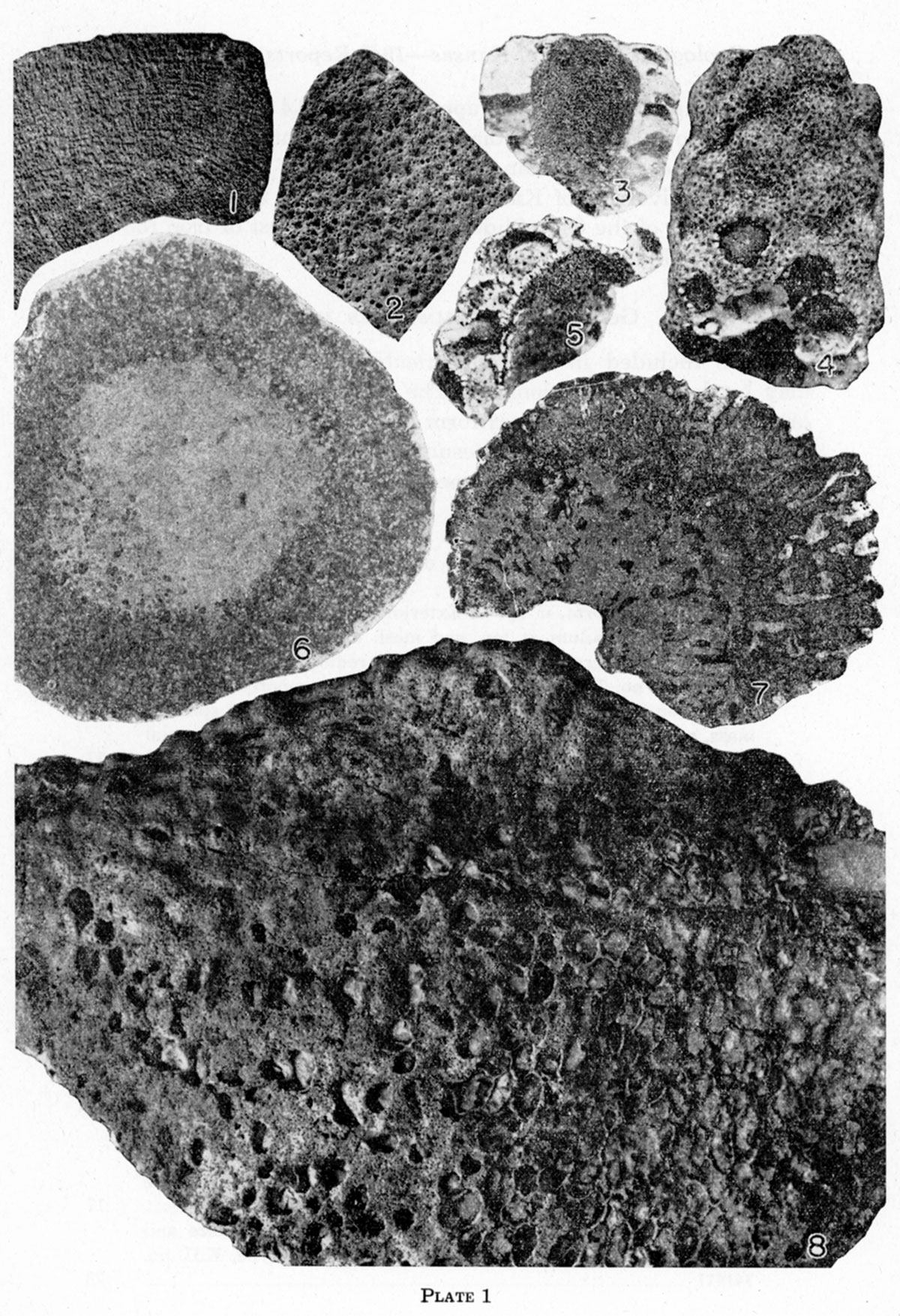
This species is much larger than any previously described representative of the genus, and the canals are actually and proportionately much larger than those of other species. The holotype is about 120 mm in diameter and its cloaca is about 40 mm in diameter. The diameter of the largest radial canal is 2.2 mm, but the average size is about 1.4 mm. Some of the pores are correspondingly large, but most are about the size of those in Heliospongia ramosa Girty; the canals increase gradually in size and anastamose as they approach the cloaca. The cloacal pores are therefore large, averaging about 1.8 mm in diameter.
Figure 1—Heliospongia vokesi, n. sp. Sketch of part of natural transverse section of type specimen, A.M.N.H. no. 25227, showing arrangement of canals and relative size of canals, cloaca, and sponge body. Prepared from an inked and bleached photograph.
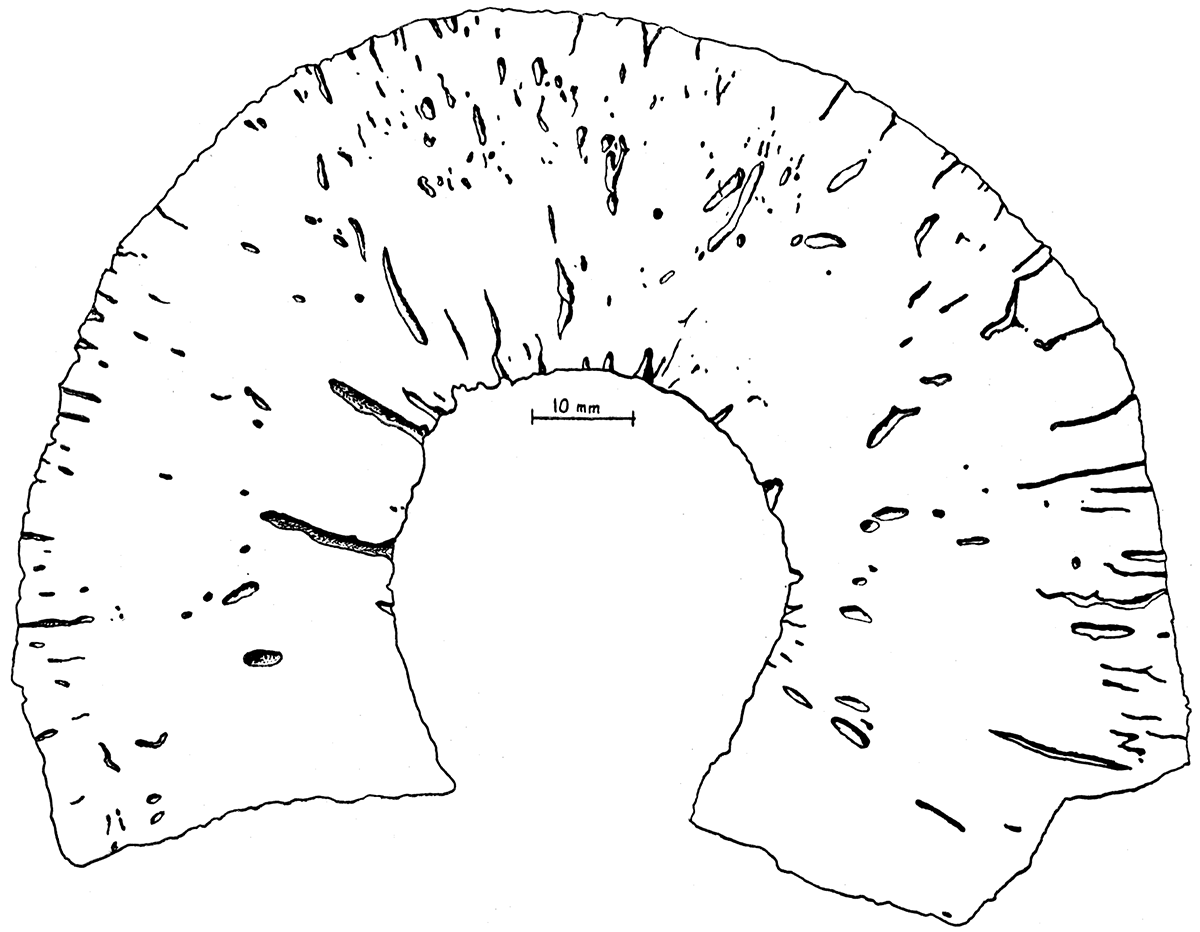
Discussion—The new species here described may be distinguished from the Pennsylvanian Heliospongia ramosa Girty, H. parallela Girty, and H. excavata King by its large size and by its large and numerous radial canals. It lacks the deep pits of H. excavata. The specimen described by Parona (1933, p. 51, pl. 9, figs. 5, 6) as Heliospongia ramosa Girty does not belong to this genus. The species is named for Harold E. Vokes of the American Museum of Natural History.
Occurrence—Upper part of the Wolfcampian series and Hess facies of the Leonardian series, Permian.
Type—American Museum of Natural History no. 25227, Leonardian series, from west side of hill back of Hess Ranch, Hess Canyon, Glass Mountains, Texas.
This genus is established for massive subconical sponges that are built of tetracladine spicules arranged in obvious horizontal layers and that are traversed longitudinally by several relatively large canals. I have not been able to determine whether these canals increase by splitting or by intercalation. Small irregularly radial canals connect the small scattered pores with the large vertical canals. The upper and lateral surfaces range from gently convex to gently concave. The openings of the longitudinal canals pit the upper surface of the sponge.
Discussion—Laubenfelsia is readily distinguishable from other associated sponges by the regularity of the horizontal banding that results from the uniform orientation of the spicules, and by the numerous large and closely spaced openings of the canals on the upper surface. Weathered specimens somewhat resemble stromatoporids, but of course the spicular structure serves to characterize them as sponges.
Girty (1908) described some spicular aggregates that are similar to those of Laubenfelsia, and applied to them the name Stromatidium. Inasmuch as the canal system and general form of Stromatidium are unknown, I regard the genus as unrecognizable, at least until study of topotype material reveals additional characteristics.
Genotype—Laubenfelsia regularis, n. sp. The genus is named for M. W. de Laubenfels, prominent contemporary American student of sponges.
Laubenfelsia regularis, n. sp.
Plate 2, Figures 3, 4, 5.
The general characteristics have been indicated in the foregoing generic description. The largest specimen studied is 83 mm high and 89 mm in greatest transverse measurement. The type is 46 mm high and 50 mm in greatest transverse measurement. On the upper surface of the type there are the openings of at least 24 canals, which average 3 mm in diameter and which are separated by spaces of the same order of magnitude. There are 16 to 18 layers of spicules in a space of 5 mm. The pores average 0.4 mm in diameter and are about 2 mm apart. They are somewhat elongate parallel to the axis of the sponge body. The small radial canals within the sponge are approximately 0.4 mm in diameter, and seem to be of uniform diameter throughout.
Occurrence—Leonardian series, Permian.Type—University of Kansas no. 74147, from the slope beside the road east of the Clay Slide, about 2 miles west of Iron Mountain, Glass Mountains, Texas.
Forms included in Pseudopemmatites are subspherical and may be depressed or elongate. The tetracladine spicules are so oriented that the inters paces form small canals radiating from a small area of the periphery, presumably the base. These tubes are the same size throughout the sponge body, new tubes being added as the surface area of the sponge increases with growth. The surface is pitted by numerous closely spaced small "pores" of almost uniform size. There are no large canals nor cloaca. On none of the available specimens is the base well enough preserved to show whether there is a dermal layer over any part of that region, but, if there is one, it is restricted to a very small area. I doubt that there is a dermal layer.
Discussion—Pseudopemmatites resembles Pemmatites Dunikowski in surface appearance and radiate structure, but the spicular element is tetracladine rather than rhizomorine. In Hindia Duncan the canals radiate in all directions from the center of the sponge body, instead of only upward and outward from the base, and the spicules, if tetracladine, are not typically so.
Genotype—Pseudopemmatites skinneri, n. sp.
Pseudopemmatites skinneri, n. sp.
Plate 3, Figures 10, 12.
Specimens of this species are as large as 70 mm in diameter and most specimens are somewhat depressed. The height exceeds the diameter in some. In longitudinal section there are six to eight canals in a segment 5 mm wide, new canals being intercalated as the diameter of the sponge increases. The transverse rays of the spicules give the canals a calibrated appearance.
Discussion—This species is not likely to be confused with any other. Its finely radiate structure distinguishes it from Defordia defuncta, n. sp., which it resembles in size and shape. Furthermore, it lacks the basal dermal layer that is characteristic of that form. Its spherical shape distinguishes it from other sponges found in the same area. Features that distinguish it from Hindia and Pemmatites are discussed in the generic description. The species is named for John Skinner, whose guidance in the field facilitated the work of collecting specimens.
Occurrence—Leonardian series, Permian.
Type—University of Kansas no. 74167, from the saddle north of Leonard Mountain on Hess ranch, Glass Mountains, Texas.
The genus Defordia comprises rough, robust sponges provided with a dermal layer on the lower portion. The canal system consists of numerous moderately large canals extending outward in all directions from the base, branching as they approach the surface of the sponge body, and a few canals parallel to the surface of the body. The spicular element has not been clearly observed, but it is a tetractine, seemingly a very simple tetracladine spicule. The relatively large openings on the surface are separated by narrow high interspaces so that the surface of the upper portion of the sponge body is very rough, but the dermal layer on the lower surface is smooth or somewhat wrinkled. In addition to the small sharp protuberances on the upper surface there are large low mounds, in some of which there is an opening somewhat larger than the ordinary canals. I do not regard these openings as the apertures of cloacae. The sponge as a whole resembles an ordinary bath sponge in appearance.
Discussion—This genus is not likely to be confused with any other with which I am acquainted, as its very rough surface sets it apart from any other except Pseudopemmatites, n. gen., from which it differs in having a basal dermal layer, large irregularly branching radial canals instead of small straight continuous canals, and low hummocks on the upper surface, some of which are penetrated by larger canals.
Specimens assigned to this genus occur in the Leonardian series, Permian, of the Glass Mountains, Texas. A single specimen from rocks classified as Word (?) formation, Permian, Chinati Mountains, Texas, belongs to this genus, though perhaps not to the genotype species.
Genotype—Defordia defuncta, n. sp. The genus is named for Ronald K. DeFord, one of the leading students of Permian stratigraphy.
Defordia defuncta, n. sp.
Plate 1, Figure 7; Plate 2, Figure 9; Plate 3, Figure 9.
Specimens of this species attain a diameter of 85 mm and are subspherical or, more commonly, depressed. On most specimens the base is a very short peduncle, which in some specimens becomes invaginated as a result of downward growth at the margins; but on some specimens the basal surface is concave. The dermal covering, which is about 0.5 mm thick, is confined to the lower surface, and may be only a few square centimeters in extent or may cover virtually the whole lower part of the surface. It is smooth or slightly wrinkled, but not rough, in contrast to the upper part of the sponge, which is covered with protuberances 1 to 2 mm high and 1 to 2 mm across, consisting of the interspaces between the openings of the canals. The height of these raised knobs represents the extent of weathering. The knobs are very irregular, almost vermiform. In addition to the relatively sharp elevations, the surface of the sponge is marked by eight to twelve broad low hummocks, the number depending somewhat on the size of the specimen. They resemble the monticules of bryozoa, except that in some of them there is an opening, 2 to 3 mm in diameter, which seems to be merely the aperture of a somewhat enlarged canal, rather than an osculum. The diameter of the radial canals is somewhat variable, but averages about 1.5 mm, excepting the few enlarged canals just mentioned. The spaces between canals are generally less than 3 mm across. The canals parallel to the surface are about the same size as the radial canals, but they are much less numerous.
Discussion—As indicated in the generic description, Defordia defuncta is not likely to be confused with any other form, except possibly Pseudopemmatites skinneri, n. sp., but that form has a distinctly different canal system and somewhat different surface texture, and lacks a basal dermal layer.
Occurrence—Leonardian series, Permian.
Type—American Museum of Natural History no. 25229, from the west side of the hill back of Hess ranch house (Hess Canyon side), Glass Mountains, Texas.
This genus includes cylindrical to subconical sponges that are covered with a dermal layer, which is perforated by pores, and that are traversed throughout by a cloaca, which is lined with a similar perforated layer. The structure of the wall is unusual and striking; successive layers of spicules are supported by pillar-like rays. Seemingly there are no canals, although the spicular mesh is very open; the whole cribwork may have constituted the canal system. The spicular element could not be isolated, but it is a tetractin, probably tetracladine, and somewhat large.
Discussion—Externally, Stylopegma closely resembles Maeandrostia Girty, of the Calcispongiae, but the wall is divided into obvious horizontal layers and the spicules are distinct. A weathered specimen resembles several of the chambered Calcispongiae, but the horizontal partitions are close together and are supported by numerous pillars. This structure contrasts with that of Amblysiphonella Steinmann, for example, in which the chambers are about as high as wide and are empty except for adventitious vesicular matter. The wall structure is morphologically most similar to that of Laubenfelsia, but is somewhat coarser; Laubenfelsia has numerous vertical canals instead of a cloaca, and it lacks a dermal layer.
Occurrence—Wolfcampian and Leonardian series, Permian, Glass Mountains, Texas, and Permian rocks of the Las Delicias area, Mexico.
Genotype—Stylopegma dulce, n. sp.
Stylopegma dulce, n. sp.
Plate 2, Figures 6, 7; Plate 3, Figure 4.
Specimens of this species attain a diameter of 18 mm and are not appreciably tapered. The surface is smooth, but there are a few broadly rounded constrictions, seemingly unconnected with any structural feature. The pores are about 0.3 mm in diame.er and are separated by distances of 0.5 to 1 mm. There are about five layers of spicules in a space of 10 mm, but the spaces between layers range from about 1.5 to 3 mm, so the number of layers in a given space is somewhat variable, and seemingly decreases with growth; that is, there is a greater space between layers in the larger, more mature specimens. The diameter of the cloaca is about 5 mm in a specimen 17 mm in diameter, and the wall is 5 to 6 mm thick.
Discussion—This species is not likely to be confused with forms belonging to any other genus. From Stylopegma conica, n. sp., it differs in being cylindrical and in having nearly fiat, widely spaced layers of spicules, instead of closely spaced, arched layers.
Occurrence—Leonardian series, Permian, Glass Mountains, Texas, and Permian rocks of the Las Delicias area, Mexico.
Type—Yale Peabody Museum no. 16594, from R. E. King's beds 5-6, Leonardian series, Permian, section on Baylor Mountains, Texas; collected by R. E. King.
Stylopegma conica, n. sp.
Plate 2, Figure 2.
This species is based on a single specimen from which the dermal layer has been eroded, but it differs so markedly from Stylopegma dulce that it seems feasible to describe the species on this otherwise inadequate material. The maximum diameter of the fragment is 28 mm and the height is 45 mm, but both were originally somewhat greater. The size and spacing of the dermal pores are unknown. There are eight or nine strongly curved layers of spicules in a space of 10 mm; the center of each layer is several millimeters higher than the edges, the convexity being greater in the upper portion of the sponge body than in the lower. Where exposed in the natural oblique section, the cloaca is nearly 5 mm in diameter; the sponge body is 26 mm in diameter at the same height.
Discussion—This species can be distinguished from Stylopegma dulce by its conical shape and by its strongly convex, closely spaced layers of spicules, which differ markedly from those of S. dulce.
Occurrence—Wolfcampian series, Permian, Glass Mountains, Texas.
Type—Yale Peabody Museum no. 16596, from bed 4, Wolfcampian series, Permian, of P. B. and R. E. King's section 24, measured at the west end of the Wolf Camp Hills along the western side of a small canyon draining through the hills from the main scarp of the mountains, Glass Mountains, Texas.
Girty (1908, p. 72) describes this genus in the following words:
Shape turbinate or pyriform, attached by the smaller end, without a peduncle or with but a small, ill-defined one. Cloaca represented by a slight depression on the upper surface, from which descend several (three of four) tubular openings through the axial region of the sponge. Ostia small and numerous. The spicular structure consists of minute, regular tetraxons, more or less digitate toward the extremities of the arms (?), where they unite with one another to form a rigid skeleton. Loose monaxial spicules associated with typical specimens may belong to the same genus.Type—Anthracosycon ficus [Girty, 1908, p. 72].
Examination of several specimens representing species referable to this genus reveals that the shape is variable, the "slight depression on the upper surface" is not a constant feature, and the number of tubular openings visible is extremely variable, for they are not even differentiable on all specimens. The largest openings observed on the specimens that I have examined are 1.5 mm in diameter, and Girty's descriptions and figures do not indicate that he observed any larger openings. On most specimens there are openings of several sizes, as would be expected if new tubes developed at any and all stages of growth. It should be mentioned that on the genotype species these tubules are visible only on the upper surface; the lateral surface is perforated by numerous fine, evenly spaced openings of almost uniform size, about 0.4 mm, separated by minutely perforate areas of the same order of magnitude.
Removal of the limestone matrix from a partly silicified specimen revealed that monactine spicules of different sorts are definitely embedded in the body of the sponge among the tetractine spicules. Furthermore, two very spinose hexactine spicules were freed, but it is realized that these may have been extraneous.
Although the canal system is an important feature in the classification of sponges, Girty did not discuss this, except to state that there are several tubular openings. In this genus the canal system consists of a series of tubules extending upward and outward from the base, augmented by a series of tubules extending radially, roughly parallel to the upper surface of the sponge body, and a poorly developed series of concentric tubules in approximately horizontal planes. None of these series is very regular, however.
The statement that the cloaca is represented by a slight depression on the upper surface is misleading. Girty's illustrations show no such feature; instead, they depict a sponge that is gently convex over the upper surface. Some of the specimens that I have studied exhibit a similar gently convex upper surface, but others actually are depressed. In one specimen the depression extends laterally into an indentation in the rim and in the side of the sponge body. Similarly, the configuration of the base ranges from slightly pedunculate to concave.
Ten specimens referable to this genus were studied by me; of these, two were definitely identified as Anthracosycon ficus Girty. The other specimens differed from these two and from each other in various minor details, so that it was not possible to decide whether they represent new species or whether the description of A. ficus should be modified to include all the specimens. Because of the present interest in problems connected with the Permian rocks it seems likely that the additional specimens needed to settle the question may be collected soon. Accordingly, it is advisable to suspend judgment.
The class Calcispongia includes sponges that are constructed of spicules originally composed of calcium carbonate. Inasmuch as this skeletal substance may be replaced by another in the process of fossilization, the present composition of the spicules is not reliable, and other criteria are used in determining the classificatory position of fossil forms. Recognizable spicules are rare in fossil calcisponges; commonly the spicular parts of the sponge are preserved as a fibrous mass. Presence of identifiable spicules, therefore, is generally regarded as indicative of a silicisponge, belonging to either the Demospongia or the Hyalospongia. Similarly, regularity of arrangement of the spicules, even though individual spicules are not discernible, commonly denotes a silicisponge. A massive wall penetrated by numerous irregular vermiform canals, as in Maeandrostia Girty, suggests a calcisponge. Now included among the Calcispongia are numerous genera that are constructed of hollow chambers, which are joined together in various ways. Typical of the Permian genera in this group is Steinmannia Waagen and Wentzel. It consists of a single row of perforated cells that communicate only through the perforations. In every attribute except its large size this genus resembles some of the Foraminifera, and accordingly its position among the sponges might be questioned. Other genera of this group, however, such as Amblysiphonella Steinmann, are traversed by a central tube, which seems to be a normal sponge cloaca. The group is somewhat anomalous, nevertheless, and some day it may be transferred to a different phylum. At present I see no justification for such transfer.
I have made no attempt to subdivide the Permian genera of the Calcispongia into groups larger than families.
Girty assigned to Guadalupia several species of sponges that differ chiefly in their mode of growth, which ranges from massive to cylindrical or to frond-like. These species have in common a peculiar type of wall structure, which is somewhat similar to that of Cystothalamia Girty and that of Polyphymaspongia, n. gen. The principal part of the wall is composed of parallel, approximately cylindrical tubes oriented perpendicular to the surface of the sponge body. Girty (1908, p. 80) characterized these tubes as "rather closely arranged, usually almost in contact." I interpret this statement to indicate that the tubes have discrete walls and are separated by small interstices, resembling in this respect the zoecia of bryozoa, which are separated by mesopores. Girty's figure 1c on plate 6, however, indicates that the tubes are all in contact, the wall between tubes being shared by both tubes, as, in Cystothalamia and in Polyphymaspongia. Likewise, the specimens that I have examined are composed of tubes separated by a single wall rather than a double wall. As Girty (1908, p. 80) states, the tubes are slightly curved and are crossed by flat diaphragms, but the diaphragms are rare in all the specimens that I have examined. Girty's figure 1d on plate 6 seems to indicate a second thinner layer of small chambers; and such a layer is present in the specimens studied by me, but I was unable to determine the nature of these smaller cells. Guadalupia resembles Polyphymaspongia in that the walls are composed of two layers of cells. The two genera are readily distinguished, however, by the size and shape of the large chambers.
Occurrence—Leonardian to Guadalupian (Capitan), Permian, Trans-Pecos Texas.
Genotype—Guadalupia zitteliana Girty.
Guadalupia williamsi, n. sp.
Plate 2, Figure 10; Plate 3, Figures 7, 11.
This species comprises large flat specimens about 3 to 4 mm thick. The type specimen is about 90 by 108 mm, but is not all exposed, and part is broken away. The central portion is depressed, but the specimen seems to have grown on an uneven surface, with which it conforms in shape. The thickness seems to be almost uniform despite this irregularity. There are about 6 tubes in a space of 5 mm, as measured on a polished section of a paratype specimen.
Discussion— This species differs from all others except Guadalupia zitteliana Girty and G. zitteliana var. Girty in its flat mode of growth. G. williamsi differs from the former in being only 3 or 4 mm thick instead of 6 or 7 mm; from the latter it differs in having about six tubes in a space of 5 mm instead of eight or nine.
Occurrence—Upper part of the Leonardian series, Permian.
Type—University of Kansas no. 74149, from the Leonardian series, Permian, exposed beside the road east of the Clay Slide, about 2 miles west of Iron Mountain, Glass Mountains, Texas. Collected by Charles C. Williams, for whom the species is named,
Sponges of this genus are large, gently convex, subdiscoidal forms composed of two layers of chambers. The cells constituting the upper layer are relatively large; and the "top" of one cell forms part of the "floor" of succeeding cells, except that the direction of growth is horizontal instead of vertical. The chambers communicate with each other, with the lower layer, and with the exterior through small pores. Vesicles are sparse. The lower layer of cells is much thinner than the upper layer, and the nature of the individual chambers is not clear, but they seem to be small and irregular. Seemingly, there is a continuous dermal layer over the entire lower surface of the sponge body. The upper surface is mammillate, each protuberance indicating the position of a chamber in the underlying upper layer of cells.
Discussion—This genus is referred to the Calcispongia because of its cellular structure, which resembles somewhat that of Guadalupia, Cystothalamia, and, to a lesser extent, the Sphaerocoelidae. Polyphymaspongia is not likely to be confused with any of the Sphaerocoelidae, however, because it is discoidal rather than cylindrical, and has two distinct layers of cells. It is distinguished from Cystothalamia by its mode of growth and also by the absence of ostia, and possibly by the presence of two layers of cells, although the chambers in Cystothalamia may be of two kinds. Only the explanate species of Guadalupia bear any close resemblance to Polyphymaspongia, but Girty (1908, p. 80) states that in Guadalupia the cells have discrete walls (although I question his statement); whereas, the chambers of Polyphymaspongia are built on the preceding chambers, using their tops for a floor. The largest chambers in Guadalupia, however, are much smaller than any of those in Polyphymaspongia, and are cyclindrical and curved, rather than shield-shaped or quadrangular in section.
Genotype—Polyphymaspongia explanata, n. sp.
Polyphymaspongia explanata, n. sp.
Plate 1, Figure 8; Text Figure 2.
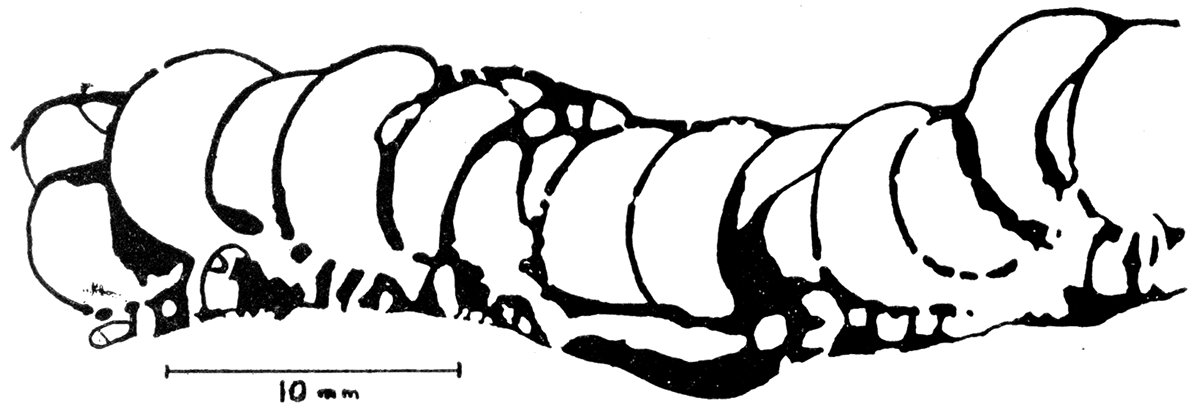
The type, a silicified specimen, is a fragment more than 165 mm in diameter. The thickness is almost uniform and averages about 12.8 mm, the upper layer of larger cells being about 9 mm thick and the lower layer of smaller cells being about 3.8 mm thick. The thickness of the individual layers is more irregular than that of the entire sponge, however, one layer thinning as the other thickens. Individual chambers in the upper layer of the type specimen average about 4 mm by 6 mm, but in another specimen there are a few compressed chambers 2 mm by 10.5 mm. Individual cells in the lower layer seem to be less than 1 mm in diameter, but they are very irregular, so this measurement is only an approximation. The center of the specimen is 33 mm above the plane of the edges, but most of the edge is broken, so that the actual convexity may be somewhat less or more, although probably the ratio is not greatly different.
The pores and internal structure were studied in more detail in a sectioned specimen preserved as calcite. The walls of the large chambers are mostly about 0.2 mm thick, although locally they are 0.5 mm thick; the walls of the chambers in the lower layer are about 0.4 mm thick. There is a very small amount of vesicular material in the chambers of the upper layer, and it is much thinner than the walls of the chambers. The pores are about 0.2 mm in diameter, or smaller, and are relatively sparse.
Discussion—An entire specimen is not likely to be confused with any other organism, but the specimens commonly found are imbedded in limestone matrix and appear as natural sections. In transverse section the upper layer resembles Cystothalamia somewhat, but it is obviously not a cylindrical form, so it is readily distinguishable from that genus. The vertical section presents some similarity to that of Girtycoelia, but the differentiation into two layers of cells distinguishes it from that genus,
Occurrence—Leonardian series, Permian, Glass Mountains, Texas.
Type—University of Kansas no. 74171, from the upper Leonardian; exposed northeast of Split Tank, 1 mile northeast of the old Word ranch house, 18 miles north of Marathon, Texas. Collected by Mrs. R. C. Moore.
Figure 2—Polyphymaspongia explanata, n. gen. n. sp. Sketch of part of polished section of a paratype specimen, K.U. no. 74172, perpendicular to plane of sponge body and parellel to long axis of chambers, showing size of pores and arrangement and relative size of the two layers of cells, Prepared from an inked and bleached photograph,
The family Wewokellidae is erected to include the Pennsylvanian and lower Permian genus Wewokella Girty and the lower Permian genus Talpaspongia, n. gen. These genera include relatively large, straight, bent, or branching cylindrical sponges that have a peculiar spicular element. The spicules consist of a large central node or sphere from which radiate numerous thick tapered rays. Hinde (1888, p. 173) based the genus Asteractinella on isolated spicules of this sort, but, inasmuch as the form of the sponge body and the canal system of Asteractinella are unknown, I regard that genus as unrecognizable.
Reference of this family to the Calcispongia may be protested on the ground that such complex spicules are unknown among the calcisponges; but, conversely, the fact that the spicules are very large, for silicisponges, is equally good reason for including the Wewokellidae in the Calcispongia. Furthermore, both Wewokella and Talpaspongia are found in typical shallow-water deposits, and the fact that the habitat of calcisponges is restricted to shallow water is my chief reason for classifying these genera in the Calcispongia. Among the numerous specimens of both genera that I have examined, there are none preserved as silica, although some specimens of Wewokella are partly replaced by barite. Admittedly, the evidence is not conclusive, for the Demospongia also can inhabit shallow water, and silica can be replaced by calcite; but, in the present state of our knowledge, the evidence seems to me to be slightly in favor of my interpretation.
Wewokella contorta, n. sp.
Plate 2, Figures 1, 8; Plate 3, Figure 5.
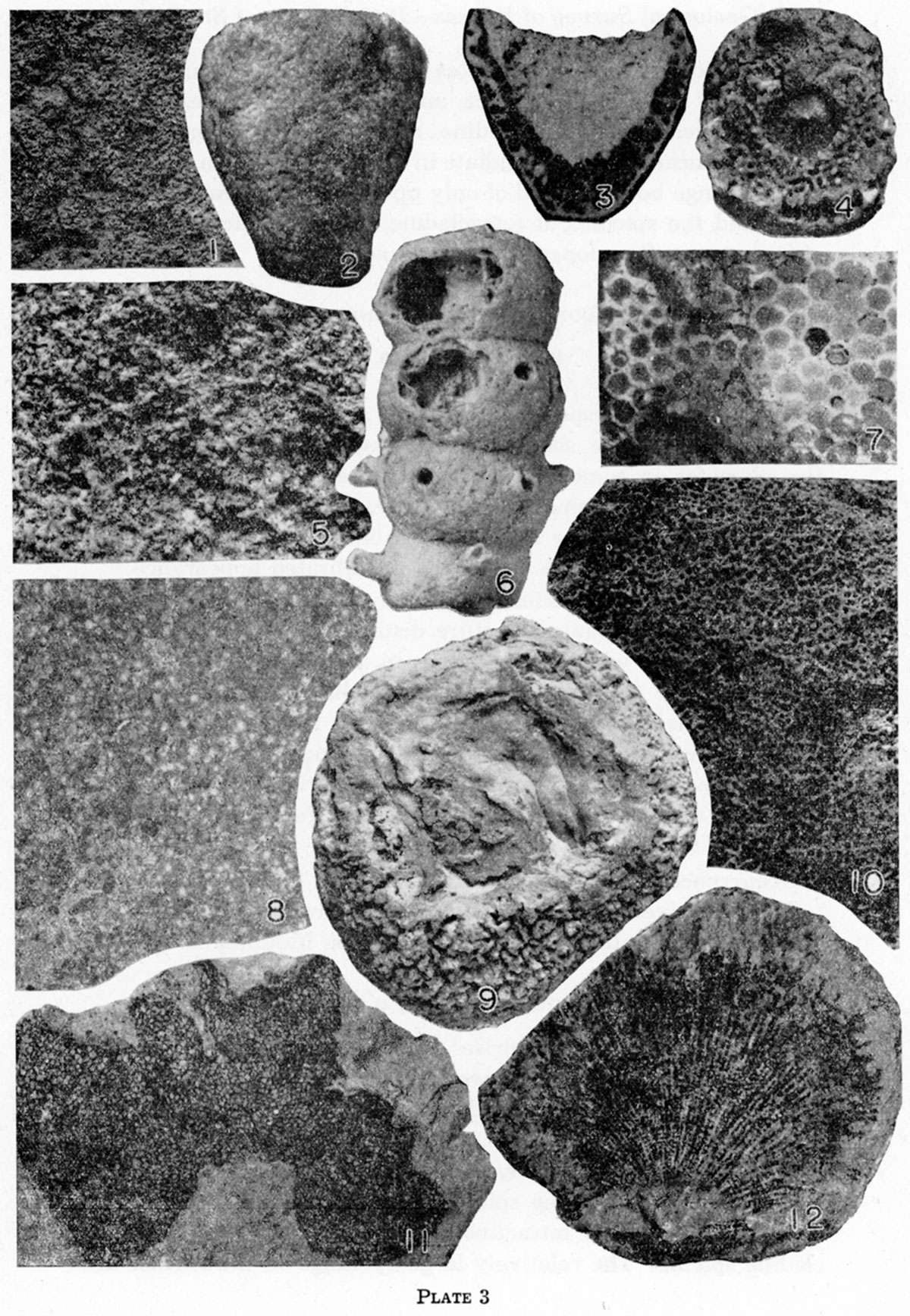
Specimens of this species are subcylindrical but are much distorted, bending somewhat abruptly through angles as great as 900, first in one direction, then in another. The maximum diameter of the largest specimen is 47 mm.
The upper end is subhemispherical, but the central portion is occupied by a conical cloaca, which has a diameter about one-third that of the sponge body at the upper surface and a depth about one or one and one-half times the maximum diameter of the cloaca. The largest cloaca measured has a maximum diameter of about 13 mm.
The dermal layer is relatively smooth, but on the specimens examined it is preserved only in small, scattered patches. The dermal spicules are relatively small; numerous rays diverge somewhat irregularly from a spherical nucleus, but all the outwardly directed rays are reduced to mere knobs.
Where the dermal layer has been removed by erosion, the skeletal spicules are exposed: They are much larger than the dermal spicules; the rays are thick at the base and they taper strongly; the nucleus is relatively small and inconspicuous. The arrangement of the rays is irregular, as in the Heteractinellidae.
Within the outer layers of spicules the body of the sponge is massive and is penetrated by somewhat tortuous, irregularly axial and radial canals. The internal diameter of the canals is about 0.4 mm and the thickness of the wall of the canals is about 0.5 mm. Much of the internal structure is only vaguely defined, because in the process of fossilization parts of the structural elements were replaced by crystalline calcite. In Wewokella solida Girty some of the body spicules are perfect octactins and are even smaller than the dermal spicules, and it is probable that those of this species are similar.
Discussion—This species differs from Wewokella solida Girty, the only other known species of the genus, in these respects. It is somewhat larger; it expands somewhat more rapidly; and it is much more contorted; both the dermal and the skeletal spicules are somewhat smaller than those of W. solida; the canals are larger, but the canal walls are relatively no thicker, and the canals do not branch as freely as do those of W. solida; the axial portion of the body does not differ from the rest, as it does in W. solida.
Type—University of Kansas no. 77671, from the shale in the "Jacksboro" limestone, Caddo Creek group, Missourian (Canyon) series, Pennsylvanian or Upper Carboniferous system, exposed near the top of a high escarpment north of and almost parallel to the left fork of the road east from Jacksboro, Texas, the right fork of which is the road to Wizard Wells. The precise locality is 1 mile east of the fork, which is about 3.3 miles east of the Jack County courthouse.
The genus Talpaspangia includes large straight, bent, or branching cylindrical sponges that are penetrated by a large central cloaca and small scattered radial canals. The surface of the sponge body is relatively smooth and is perforated by small pores. The weathered surface is rough and reveals parts of the large spicules. The spicular element resembles that of Wewokella, the kind of spicule for which Hinde proposed the generic name Asteractinella, and consists of a central spherical node from which numerous thick tapered arms radiate.
The position of Talpaspongia in the classification has been discussed in connection with the description of the new family Wewokellidae. Talpaspongia may be distinguished from Wewokella by its greater size, relatively finer general internal texture, and particularly by the presence of a cloaca throughout its entire length rather than in the terminal portion only, as in Wewokella.
Occurrence—Lower Permian of central Texas.
Genotype—Talpaspongia clavata, n. sp.
Talpaspongia clavata, n. sp.
Plate 1, Figure 6; Plate 3, Figures 1, 8.
The specimens studied are all fragments that are broken at both ends. The largest is about 60 mm in diameter; another specimen tapers rapidly from 35 to 10 mm in a length of 105 mm; but most fragments are about 30 or 40 mm in diameter. Four fragments that fit together form a semicircle that measures 600 mm from end to end around the are, and the diameter of the sponge body is almost uniformly 35 mm throughout this length. The diameter of the cloaca in a specimen 58 mm in diameter is 22 mm; but in another specimen, only 35 mm in diameter, it is 19 mm. The canals are 0.3 to 0.4 mm in diameter. They branch and anastamose very irregularly; they open to the exterior through pores, and into the cloaca through small round openings, Much of the cloaca is occupied by granular material, which may be adventitious, but may be part of the sponge, as similar material fills all but the uppermost part of the cloaca of Wewokella.
Discussion—I know of no other species with which Talpaspongia clavata is likely to be confused. Its large size sets it apart from all other known sponges from Pennsylvanian and Permian rocks except Heliospongia, which is easily distinguished by the fact that its spicules are oriented in distinct radial, concentric, and longitudinal rows. The morphologically more closely related Wewokella lacks a cloaca except at the extreme end, has a coarser spicular element, and is generally less than 25 mm in diameter.
Occurrence—Paint Rock and Talpa formations, Permian, central Texas. One poorly preserved fragment from the Hueco is identified tentatively as belonging to this species.
Type—University of Texas Bureau of Economic Geology no. 8740, from the Talpa formation exposed at Herrings Bluff, on Colorado River, Runnels County, Texas. Collected by J. W. Beede.
Girty (1908, p. 88) erected the family Cystothalamiidae to receive a single Permian genus and species, Cystothalamia nodulifera Girty. It is not surprising, therefore, that he interpreted some generic characteristics as family characteristics. The Pennsylvanian genus Cystauletes, herein described as new, seems to be a forerunner or ancestor of Cystothalamia and is evidently more closely related to that genus than to any other now known, and almost certainly it belongs to the same family. The characteristics common to these two genera, therefore, may be regarded as the characteristics of the family.
The general form is cylindrical or very slightly tapered. The upper end is depressed into a profound cloaca. Girty states that this feature does not extend to the base, and in the generic description of Cystothalamia, the only representative of the family known to him, he states that it is probable but not certain that a more or less profound cloaca was present in the upper end, All specimens of Cystauletes, however, possess a large, smooth, well-defined central cloaca. The depth of the cloaca, therefore, is a generic characteristic but its presence is a family characteristic.
The principal characteristic of the family is the cystose composition. In Cystauletes the cysts are arranged in more or less regular diagonal rows and there is only one layer of cysts surrounding the cloaca. In Cystothalamia, in which the cloaca is restricted to the distal portion, the outer layer of cysts is disposed in more or less regular diagonal lines but the cysts within, i.e., those that occupy the place of the cloaca, are irregularly arranged. In both genera the walls of the cysts are single, the roofs of the first-formed cysts serving as a floor for those formed next. The external surface is nodose, indicating the cystose construction. The wall of the cloaca, the partitions between cysts, and the external wall are all perforated by numerous small openings.
The external wall is thin and is perforated by numerous closely spaced pores, and, in Cystothalamia but not in Cystauletes, by questionable ostia. The perforation of the outer wall by many small openings is a family characteristic; presence or absence of so-called ostia is not significant. I am convinced that in the classification of fossil calcisponges especially, but to some extent in the classification of all fossil sponges, unwarranted importance has been ascribed to the size of the small openings in the walls. After all, the transition from pores of uniform size through pores of unequal size to pores of two sizes or pores and ostia is gradational, and the difference is a difference of degree; we are not justified in assuming that it is also a difference of kind, as it is in some living sponges.
Judging by characters of the two known genera, the evolutionary trend within this family was a progressive recession of the cloaca toward the distal portion of the body and increased irregularity of size and arrangement of the pores.
Most of the characteristics of the genus have been indicated in the emended description of the family. The specimens consist of straight, bent, or branching cylindrical bodies traversed by a large smooth central cloaca. The body is divided into hollow cysts arranged in more or less regular diagonal rows forming a single layer of cells surrounding the cloaca. All walls are single and are perforated by numerous small pores. The external surface is nodular, each node corresponding in position with a cyst in the body. The spicular structure is unknown.
Discussion—This genus differs from Cystothalamia in having pores of one size only, almost uniformly arranged, and in having a cloaca throughout its entire length rather than in the distal portion only.
Genotype—Cystauletes mammilosus, n. sp.
Cystauletes mammilosus, n. sp.
Plate 1, Figures 3, 4, 5.
This species consists of cylindrical straight, bent, or branching bodies as much as 20 mm in diameter. The external surface is a continuous series of low rounded elevations, which resemble the monticules of bryozoa. These elevations are disposed in more or less regular diagonal lines. On a specimen 10 mm in diameter they are about 4 mm across, and on a specimen 17 mm in diameter they are about 6 mm across.
The outer wall is perforated by numerous small, regularly spaced pores, most of which are about 0.3 mm in diameter. The spaces between are about one to two times as wide as the pores.
The cloaca is large and smooth and its diameter is approximately 8.5 mm in a sponge 18 mm in diameter, but only 3 mm in one 9 mm in diameter. It is perforated by numerous small vermiform openings. The spaces between are noticeably smaller than the pores.
The size of the cysts varies with the size of the sponge. In a specimen 17 mm in diameter the cysts are about 5 mm in inside diameter, and the walls are about 0.5 mm thick. In a specimen 10 mm in diameter the cysts are about 2.5 mm in inside diameter, and the walls are about 0.3 mm thick. The thickness of the walls may have been slightly increased or decreased, however, during the process of replacement of the original calcareous skeleton by silica, a process that has altered all the specimens. This same process has obliterated all trace of spicular structure.
No other species so far known is likely to be confused with this form.
Occurrence—Upper member of the Pawnee limestone, Marmaton group, Des Moinesian series, Pennsylvanian or Upper Carboniferous system.
Type—University of Kansas no. 74131, from the west bluff of Verdigris River, east of Nowata, Okla., on U.S. highway 60, which is the center of the north line of sec. 35, T. 26 N., R. 16 E. Collected by R. C. Moore.
(Heterocoelia Girty, 1908, not Dahlbom, 1854)
Sponges of the genus Girtyocoelia (not to be confused with Girtycoelia King, 1933) comprise straight or bent stems composed of a series of spherical chambers slightly overlapping, just in contact, or separated by a space bridged only by the central cloaca, which traverses the entire stem. The outer wall of each chamber is perforated by a few large openings, the margins of which may be extended outward as spouts. There seems to be no communication between cells except through the cloaca; pores open into the cloaca from each chamber. The whole sponge body resembles somewhat a string of beads.
Occurrence—Missourian series, Pennsylvanian, to Leonardian series, Permian.
Genotype—Heterocoelia beedei Girty.
Girtyocoelia dunbari, n. sp.
Plate 3, Figure 6.
Specimens referred to Girtyocoelia dunbari, n. sp., are somewhat large for the genus, attaining a diameter of 14 mm, although the average size is nearer 12 mm. The internal diameter of the cloaca on a specimen of average size is about 2 mm. The openings are much more extended than those of either of these diameter, and the margins of some of them are extended outward as far as 3.5 mm as prominent tubular spouts. The pores connecting the chambers with the cloaca are small and numerous, and seemingly tend to be grouped in two rings where the upper and lower walls of the chamber meet the cloaca.
Discussion—This species is larger than either Girtyocoelia beedei (Girty) or G. sphaerica (King) and the external openings are much more extended than those of either of these two species. In degree and uniformity of overlap of the chambers, G. dunbari resembles G. sphaerica but can be distinguished by the small size of the cloacal pores, as well as by the extreme extension of the external pores. G. dunbari is much larger than G. beedei; the chambers overlap more regularly; and the cloaca is actually and relatively much larger than that of G. beedei.
Occurrence—Leonardian series, Permian, Glass Mountains.
Type—Yale Peabody Museum no. 16595, from the Leonardian series, first limestone member, west end of Dugout Mountain, Glass Mountains, Texas, collected by R. E. King. The species is named for C. O. Dunbar, who graciously submitted the Yale material for study.
This genus is erected to include a single very primitive species of calcisponge. The characteristics of the genus are its small size, its subconical shape, its large, upwardly expanding central cavity, and its relatively thin walls, which are penetrated transversely by vermiform canals that open on the inner and outer surfaces of the sponge body in irregular small pores.
Discussion—I know of no other genus of calcisponges with which this form is likely to be confused. The primitiveness of its development is indicated by its small size, its simple subconical shape, and its simple, meandering canals. The canals resemble those of the Pennsylvanian genera Maeandrostia Girty (1908a, p. 284) and Fissispongia King, (1938, p. 499) but both those genera are straight, bent, or branching cylindrical stems. All three genera may belong to one family; but, nevertheless, all three differ distinctly from each other in the characteristics that are of generic importance. The outer surface of Cotyliscus is perforated by numerous small irregular pores; that of Maeandrostia Girty is covered by a dermal layer that is perforated by a few large scattered pores; and that of Fissispongia King is also covered by a dermal layer, which is penetrated by very minute openings and also bears scattered spinose projections. Furthermore, Cotyliscus has a large, subconical central cavity, hardly deserving to be dignified by the term cloaca; whereas, both the other genera mentioned possess a prominent tubular cloaca.
Occurrence—Osage series, Lower Carboniferous, central Texas.
Genotype—Cotyliscus ewersi, n. sp.
Cotyliscus ewersi, n. sp.
Plate 3, Figures 2, 3.
Specimens of this species reach a maximum height of about 5 mm and the maximum diameter is about 4.5 mm. The basal angle is acute, but the base is sharply to moderately rounded. The walls are actually and relatively thickest at the base. At the top of the specimens, the diameter of the central cavity is twothirds to three-fourths that of the entire sponge body. The cavity penetrates the upper three-fourths of the sponge body.
The thin walls are perforated by fine canals about 0.05 mm in diameter. These canals join and branch somewhat, but are predominantly directed perpendicular to the surface. There is a suggestion of differentiation of the wall into two layers.
The outer surface of the sponge body presents a stippled appearance, owing to the abundant fine pores, a few of which are long and narrow rather than circular or only slightly elongate. Also scattered over this surface there are several faint low elevations separated by distances of 0.5 to 1 mm. Most, if not all, are perforated by circular openings, which seem to be very slightly larger than those perforating the surface between the elevations. These openings might be termed ostia, although they seem to differ from the pores only in size and in elevation above the surface.
The inner surface of the sponge body is also perforated by numerous pores; but these are somewhat larger and more widely spaced than the external pores, and tend to be circular rather than elongate, as are those opening to the exterior.
Occurrence—Shaly basal portion of the Chappel limestone, Osageian series, Mississippian or Lower Carboniferous system.
Type—University of Kansas no. 60851, from the south side of Colorado River, about 1 mile east of Marble Falls, Texas, above the old dam site. Collected by J. D. Ewers, for whom the species is named,
[Note: scales listed on plate figure captions may be not be correct on web presentation.]

Cossman, Maurice, 1909, Rectification de nomenclature: Rev, crit. Paleozool., vol. 13, pp, 67-68.
Gerth, H., 1929, Die Spongien aus dem Perm von Timor: Palaont, v. Timor, Lief, 16, Abh, 27, pp. 1-35, pls. 219 (1) - 224 (6),
Girty, G. H., 1908, The Guadalupian fauna: U. S. Geol. Survey, Prof, Paper 58, pp. 1-651, pls. 1-31.
Girty, G. H., 1908a, On some new and old species of Carboniferous fossils: U. S, Nat, Mus., Proc., vol. 34, pp. 281-303, pls. 14-21.
Girty, G. H., 1911, On some new genera and species of Pennsylvanian fossils from the Wewoka formation of Oklahoma: New York Acad. Sci., Annals, vol. 21, pp. 119-156.
Girty, G. H., 1915, Fauna of the Wewoka formation of Oklahoma: U. S. Geol. Survey, Bull. 544, pp, 1-353, pls, 1-35,
Hinde, G. J., 1888, A monograph of the British fossil sponges, part 2, sponges of the Paleozoic group: Paleontographical Society, vol. 41, pp, 93-188, pl. 9.
King, Ralph H., 1933, A Pennsylvanian sponge fauna from Wise County, Texas: Univ. Texas, Bull. 3201, pp. 75-85, pls. 7-8,
King, Ralph H., 1938, Pennsylvanian sponges of north-central Texas: Jour. Paleontology, vol. 12, pp. 498-504, figs. 1-14.
Laubenfels, M. W, de, 1936, The sponge fauna of the Dry Tortugas: Carnegie Inst. Washington, Pub. 467, pp. 1-225, pls, 1-22.
Parona, C. F., 1933, Le spugne della fauna Permian a di Palazzo Adriano (Bacino del Sosio) in Sicilia: Soc. Geol. Italinia, Mem. vol. 1, pp, 1-58, figs. 1-7, pls. 1-12.
Kansas Geological Survey
Placed on web Jan. 16, 2019; originally published February 27, 1943.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/47_1/index.html