Kansas Geological Survey, Bulletin 157, Part 5, originally published in 1962
Originally published in 1962 as Kansas Geological Survey Bulletin 157, Part 5. This is, in general, the original text as published. The information has not been updated.
A profusely fossiliferous limestone bed in the Beil Member of the Lecompton Limestone near Grover Station, Douglas County, Kansas, contains essentially in situ remains of an ancient community of organisms. The extremely diversified faunal assemblage, rock texture, and cyclothemic position of the bed suggest offshore deposition in relatively warm, quiet marine water of normal salinity. The Beil bryozoan fauna, as evaluated on collections from the Grover Station locality and three other exposures, is dominated by fenestrate cryptostomes, Fistulipora decora (Moore and Dudley), and Rhombopora lepidodendroides Meek; the latter species is probably the more abundant. Other bryozoan faunal elements described herein include two new species of Tabulipora Young and three new species distributed among the genera Fistulipora M'Coy, Eridopora Ulrich, and Stenopora Lonsdale. The collections include a single specimen that is assigned to the genus Meekopora Ulrich. Fenestrate bryozoans are not described.
During the spring of 1953, this author examined 13 good exposures of the Beil Limestone Member of the Lecompton Limestone (Virgilian) in east-central Kansas. The Beil contains abundant fossils at many localities, but large numbers of specimens are difficult to collect from a single bed because most Beil exposures are in roadcuts or stream banks where strata can be viewed only in nearly vertical exposures. In the SW NE sec. 35, T. 11 S., R. 17 E., approximately 1 mile east of Grover Station, Douglas County, Kansas (Loc. 1), a thin but profusely fossiliferous Beil stratum, hereafter termed "the fossil bed," is exposed across the floor of a small quarry and affords an excellent opportunity to collect extensively from a deposit that represents but a moment in geologic time (Fig. 1, arrow). This author noted bryozoans in most Beil exposures, but only at the Grover locality could sufficient material be collected from a single bed to insure adequate representation of the preserved bryozoan assemblage. Hattin supplemented his 1953 collections by obtaining more material in 1955 and 1959. Staff members of the Kansas Geological Survey later collected additional specimens from nearby Beil exposures (Loc. 2-4) for comparative purposes.
Figure 1--Graphic section of Lecompton Limestone near Grover Station. Kansas (Loc. 1). Arrow points to the fossil bed.
The present study broadens the rather scanty knowledge of Pennsylvanian Bryozoa and is the outgrowth of a special research project assigned to Perkins, then a graduate student in the Indiana University Department of Geology. The pre-systematic part of this publication was prepared by me; the systematic description of the bryozoan fauna was undertaken by Perkins in collaboration with Perry, who made all thin sections utilized in this study and prepared the plates.
Bryozoans from the fossil bed at the Grover locality provide an unusually fine opportunity to study skeletal remains of the bryozoan portion of an ancient community of marine organisms and allow delineation of structural variation in species that lived in a very small area of the sea floor during a very short interval in earth history. Members of a species living in such a restricted community would expectably show less structural variation than representatives of the species obtained from an appreciably greater thickness of strata such as, for example, the entire Beil Member. In the fossil bed we have the advantage of dealing with portions of interbreeding populations, each representative of a true biological species.
Bryozoans and other fossils forming the basis of this study were obtained from the following exposures of the Beil Member:
All specimens studied are in the Indiana University Paleontological Collections.
The Beil is one of four fusulinid-bearing members of the Lecompton Limestone. Moore (1936, p. 28) stated that fusulinid limestones represent phases of maximum marine transgression in cyclothems that characterize the Shawnee Group in Kansas. According to this interpretation the fusulinid part of the Beil is the center bed of one of four cyclothems represented in the Lecompton. Each cyclothem does not reflect the same environmental succession because the lithologic sequence in each cyclothem differs from the others in one or more basic aspects. Moreover, individual cyclic phases show considerable lateral variation. For example, the Beil, which includes three cyclic phases in the type area and in northern and south-central Kansas, is represented wholly by rocks of biostromal character in parts of Osage and Chautauqua Counties, Kansas (Brown, 1956). Thus, the Lecompton Limestone cannot be regarded as resulting from sedimentation during simple strandline oscillations, giving rise to symmetrical repetition of the same kinds of rock in each cyclothem, but rather as reflecting a complex succession of environments, some of which were established only once during the deposition of the formation.
The Beil section near Grover Station consists upward of fusulinid limestone, gray shale containing thin beds and lenses of limestone, and a thin limestone bed that includes algal remains (Fig. 1). This sequence is representative of the C.5 (center bed) and C.6 and .7 (regressive) phases, respectively, of the C cyclothem in a typical Shawnee megacyclothem (Moore, 1936, p. 31). Algal limestone at the top of the Beil is usually not well developed in the Kansas River area (Brown, 1956, p. 89), but the thin limestone layer observed at the top of the Beil at the Grover locality contains calcareous structures that were built by algae. Fossils considered in this paper are from the uppermost part of the C.5 (fusulinid limestone) phase of the Beil cyclothem and apparently reflect a nearly static sedimentary condition existing immediately prior to the influx of terrigenous detritus that heralded regressional phases of sedimentation in the cyclothem.
Invertebrate remains collected from the fossil bed at the Grover Station locality are well-preserved, mostly whole specimens and show little sign of abrasion. All observed specimens of ?Syringopora are oriented so that the corallites are directed upward. Most of the numerous brachiopod valves remain articulated. Thin-section study shows that the limestone is an argillaceous biomicrite which is partly recrystallized to biomicrosparite. The microcrystalline-calcite-ooze matrix represents deposition in quiet waters where winnowing action of currents was at a minimum. Such observations suggest that we are investigating the remains of a fossil community preserved at the original site of growth in quiet water.
In addition to Bryozoa--and various invertebrates collected by others but not included in collections that furnish the basis of this study-the fossil bed at the Grover locality includes the following species of macrofossils, as identified by me:
| Coelenterata | |
| Caninia torquia (Owen) | |
| Stereostylus sp. | |
| ?Syringopora multattenuata McChesney | |
| Brachiopoda | |
| Antiquatonia portlockianus crassicostatus (Dunbar and Condra) | |
| Cancrinella boonensis (Swallow) | |
| Neochonetes granulifer (Owen) | |
| Composita subtilita (Hall) | |
| Crurithyris planoconvexa (Shumard) | |
| Derbyia bennetti Hall and Clarke | |
| Dielasma bovidens (Morton) | |
| Echinaria moorei (Dunbar and Condra) | |
| Hustedia mormoni (Marcou) | |
| Hystricula wabashensis armata (Dunbar and Condra) | |
| Juresania nebrascensis (Owen) | |
| Linoproductus prattenianus (Norwood and Pratten) | |
| Meekella striatocostata (Cox) | |
| Neospirifer dunbari King | |
| Phricodothyris perplexa (McChesney) | |
| Punctospirifer kentuckyensis (Shumard) | |
| Mollusca | |
| Allorisma sp. (molds) | |
| Amphiscapha muricata Knight | |
| Aviculopecten sp. | |
| Bellerophontid molds | |
| Euconospira sp. cf. E. turbiniformis (Meek and Worthen) | |
| Septimyalina sp. | |
| Trepospira sp. | |
| Echinodermata | |
| Crinoid columnals and plates | |
| Echinoid spines and plates | |
Microfossils include abundant Triticites, ostracodes, and arenaceous Foraminifera. The abundance and diversity of the fauna suggest optimum conditions for growth and proliferation of the benthos during deposition of the Beil cyclothem. For such a community to prosper, the waters were obviously well oxygenated, probably by gentle current circulation, and the food supply was large. Had the salinity been far from normal, the fauna would probably contain much larger numbers of only a few species. Relatively warm water is indicated by development of large numbers of species having calcareous skeletons of which many are quite robust (Sverdrup and others, 1942, p. 853).
Although the limestone was laid down as a soft oozy mud, many kinds of benthonic animals, including sessile forms, were able to thrive in the area. A few species, such as the bellerophontids and other gastropods, lived a vagrant life and did not require a firm substratum, and some or perhaps all of the pelecypods are infaunal elements. As Rudwick (1961, p. 476) has pointed out, articulate brachiopods can anchor their pedicles in a soft substratum, and some of the Beil species may have been held to the sea floor in this manner. As some of the foregoing forms of invertebrates became established on (in) this part of the sea floor, sessile species requiring solid attachment could gain a foothold by attaching themselves to skeletons of mobile animals either before or after the death of the latter. Ultimately the sessile forms themselves provided a firm substratum for more individuals.
The best record of epizoal attachment in the Beil fauna is displayed by corals and bryozoans. ?Syringopora is attached mostly to bellerophontid gastropods, but one colony became attached to a large Derbyia? and another grew upon a Fistulipora. The coral Caninia, attached at the apical end, grew essentially upright. As these solitary forms increased in size, topheaviness, combined perhaps with gentle current action, caused overturning of the substrata, sometimes slowly, as indicated by corallites having smoothly curved axes, and abruptly in those specimens that show angular changes in growth direction.
Among the Bryozoa, Fistulipora best shows the mode of attachment and kind of substratum. Members of this genus are attached to Caninia, Composita, Meekella, and fenestrate bryozoan fronds, but at least one specimen apparently lay directly on the sea floor. Fistuliporoids attached to Caninia commonly have all apertures directed toward the calyx and show, therefore, that the bryozoans grew while the corals were erect. A zoarium that almost completely embraces the corallite of a Caninia obviously grew while the coral was upright. On specimens of Composita, Fistulipora is usually attached to pedicle valves rather than to brachial valves, but this evidence does not prove that epizoal attachment occurred while the brachiopods were alive. Indeed, on one specimen of Enteletes a fistuliporoid grew across the line of commissure, which proves that the brachiopod was dead at the time of zoarial growth. Few ramose bryozoans in the fossil bed show evidence of attachment to a substratum. One specimen is fixed to the edge of the calyx of a Caninia and grew parallel to the coralline axis, apparently after the coral had died. Another ramose form is oriented nearly perpendicular to the concave side of a curved Caninia on which it grew after the coral had reached a semireclined position on the sea floor. A third ramose bryozoan grew on a large unidentified gastropod in a manner that suggests upward orientation of the gastropod aperture and hence attachment of the colony after the death of the mollusk. From the foregoing discussion we can conclude that bryozoan inhabitants of this community apparently utilized for attachment whatever solid substratum was available, the major control apparently being size rather than kind of object on which a zoarium grew. Shells or skeletons of the smaller organisms may have grown or lain too deep in ooze at the sediment-water interface to furnish suitable attachment for Bryozoa.
In conclusion, the texture of the limestone and the composition and mode of preservation of the fauna indicate that Bryozoa in the fossil bed at the Grover Station locality are part of an ancient community that occupied an oozy portion of a sea floor lying below wave base, relatively far from land; further, the community was surrounded by well-oxygenated and relatively warm water of normal salinity in which an abundant food supply insured proliferation of many species. Influx of terrigenous sediments heralding regression of the sea brought an abrupt end to life in this flourishing community.
Dr. D. F. Merriam, Mr. Richard Schuman, and Miss Polly Smith of the Kansas Geological Survey collected additional material for our study; technologic facilities, including photography by Mr. George Ringer of the Indiana Geological Survey, were provided by Indiana University.
Order CYCLOSTOMATA Busk, 1852
Family FISTULIPORIDAE Ulrich, 1882
Genus FISTULIPORA M'Coy, 1850
FISTULIPORA DECORA (Moore and Dudley), 1944
Pl. 1, fig. 2-6.
Cyclotrypa decora MOORE and DUDLEY, 1944, Geol. Survey Kansas Bull. 52, p. 275-276, pl. 5, fig. 8; pl. 10, fig. 7; pl. 18, fig. 1; pl. 24, fig. 1, 7; pl. 32, fig. 4.
External features--Zoarium typically hemispherical, having upper surface mildly to moderately convex, and displaying concave, less commonly planar, concentrically wrinkled base; rarely zoarium attached to brachiopod valves; basal diameter and maximum thickness of largest specimen 56 mm and 20 mm, respectively. Zooecial apertures circular or subcircular, not in contact, and having conspicuously elevated lunaria generally forming one-third to nearly one-half of the zooecial periphery. Interzooecial vesicles angular or subangular, usually one or two between adjacent zooecia, but clearly observable only in local areas. Monticules nearly flush with surface or moderately elevated, commonly 6 to 9 mm apart measuring from center to center, 2 or 3 mm in diameter, and having centers composed of vesicles flanked by zooecia somewhat above average in size and displaying lunaria directed toward monticular center.
Tangential section--Zooecia broadly ovate to nearly circular, usually thin walled, and never in contact; typically (67 of 150 measurements on 15 specimens) three entire zooecia and part of a fourth in a 2-mm distance; commonly (51 of 150 measurements) four complete zooecia in this distance; less commonly (17 measurements) three entire zooecia or (14 measurements) four whole zooecia and part of a fifth in 2 mm; rarely (1 measurement) five entire zooecia in this distance; average maximum zooecial diameter, including lunarium, 0.40 mm, ranging from 0.32 to 0.49 mm for 150 measurements on 15 specimens; mode of maximum zooecial diameter 0.37 mm; average interzooecial distance 0.20 mm, ranging from 0.08 to 0.32 mm for 154 measurements on 15 specimens; mode of interzooecial distance 0.16 mm. Lunarium moderately conspicuous, usually somewhat thickened and broadly arched, and occupying one-quarter to nearly one-half of zooecial periphery; lunaria poorly defined on some zooecia and not discernible on others. Interzooecial tissue consisting of angular to subangular vesicles smaller than a typical zooecium and usually one or two, rarely three, between adjacent zooecia. Monticular centers are composed of one or two unusually large vesicles and intercalated smaller vesicles; zooecia on monticular flanks somewhat larger than those of intermonticular areas and having lunaria directed toward monticular center.
Longitudinal section--Zooecia thin walled throughout their length and erect through most of zoarium, somewhat recumbent near base of colony, and radiating outward from central basal growth area. Diaphragms thin, horizontal or obliquely inclined, and usually spaced from somewhat less than one-half a zooecial diameter apart to about one and one-half zooecial diameters apart. Interzooecial tissue consisting dominantly of superposed subrectangular vesicles forming laterally imbricating columns; elsewhere, interzooecial tissue of crescentic imbricating vesicles convex outward and nearly rectangular superposed vesicles.
Remarks--Fistulipora decora is the most abundant species in our collections of Beil bryozoans with the exception of Rhombopora lepidodendroides. This fistuliporoid is found at all collecting localities but is especially abundant at localities 1 and 4. The foregoing description is based on examination of thin sections prepared from 15 specimens.
Microstructural features of our material correspond well with those cited by Moore and Dudley (1944, p. 276). Mean zooecial diameter given by Moore and Dudley is 0.40 mm, which coincides precisely with our value of this taxonomic character. Average interzooecial distance recorded by Moore and Dudley is 0.16 mm as compared with 0.20 mm in our specimens. Further, the number of zooecia in a 2-mm distance, calculated from a figure (pl. 18, fig. 1) of the type, compares closely with our material. Our specimens have the same general zoarial form as the type material of F. decora but differ in displaying monticules that commonly are nearly flush with the surface of the colony; conceivably, because of their low elevation these structures could have been termed maculae by Moore and Dudley, who record these structures, but not monticules, in this species.
We assign the species to Fistulipora because the majority of the zooecia exhibit moderately conspicuous broadly arched lunaria that in thin sections commonly occupy a significant part of the zooecial periphery.
Locality--1, 2, 3, 4.
Types--Hypotypes: figured, IU 6780-6783; unfigured, IU 6784-6799.
FISTULIPORA? PECULIARIS Perkins & Perry, n. sp.
Pl. 2, fig. 1-6.
External features--Zoarium solid, ramose, robust; the largest incomplete colony 71 mm long and 22 mm in diameter between two areas of bifurcation. Zooecia thick walled, round or broadly ovate, and commonly displaying elevated lunaria. Monticules sparse, irregularly distributed, nipplelike, rising about 1 mm above zoarial surface, approximately 3 mm in diameter, and lacking zooecia in their centers.
Tangential section--Zooecia nearly circular or broadly ovate, less commonly pyriform, generally not in contact, and having transversely fibrous walls usually ranging in thickness from 0.03 to 0.06 mm; typically (31 of 45 measurements) three entire zooecia and part of a fourth in a 2-mm distance; commonly (11 of 45 measurements) four complete zooecia or rarely (3 measurements) three entire zooecia in this distance; average maximum diameter of zooecial apertures, including lunaria but excluding wall thickness, 0.38 mm, ranging from 0.31 to 0.46 mm for 75 measurements; mode of maximum zooecial apertural diameter 0.37 mm; average interzooecial distance 0.14 mm, ranging from 0.05 to 0.30 mm for 72 measurements; mode of interzooecial distance 0.15 mm. Lunaria poorly developed to moderately conspicuous, not significantly thicker than remainder of zooecial wall, broadly conical, and usually not occupying more than one-quarter of the zooecial periphery; lunaria virtually riot discernible on some zooecia. Interzooecial tissue consisting of angular or subangular vesicles smaller than a typical zooecium and usually one, rarely two, between adjacent zooecia. Monticular centers lacking zooecia and composed of a mosaic of polygonal vesicles of variable size; zooecia on monticular flanks somewhat larger than those in intermonticular areas and having lunaria crudely aligned toward monticular center.
Longitudinal section--Zooecia in axial region thin walled, erect, and containing diaphragms usually spaced one-half to nearly two zooecial diameters apart; in outer part of axial region, approximately 5 mm below zoarial surface, zooecia bend outward, become thicker walled, and display diaphragms spaced one-quarter to three-quarters of a zooecial diameter apart after having assumed position normal to zoarial surface, except in region extending about 2 mm below surface of colony where diaphragms commonly lacking or rare. Interzooecial vesicles in axis loosely arranged, vertically elongated, and commonly approaching or exceeding in height a zooecial diameter; vesicles in outer part of zoarium compressed, almost invariably wider than high, crudely rectangular or crescentic, commonly arranged in mutually imbricating columns, and very gently convex outward.
Remarks--Fistulipora? peculiaris is represented in our collections by three large zoarial fragments from which 11 thin sections were prepared; fragments of each zoarium remain in the collections. One specimen (IU 6828) is assigned to this species with some hesitancy because of difficulty in obtaining properly oriented thin sections.
In its internal microscopic features F.? peculiaris does not resemble structurally any previously described Pennsylvanian or Permian species of the genus. In zoarial form F.? peculiaris is somewhat reminiscent of F. amsdenensis described by Perry and Gutschick (1959, p. 315-316) from the Amsden Formation of southwest Montana, which may be either Pennsylvanian in age or include the Mississippian-Pennsylvanian boundary. F.? peculiaris and F. amsdenensis differ most strikingly in the nature of the interzooecial tissue, especially as observed in longitudinal section.
The most conspicuous structural feature of F.? peculiaris is the change in character of the interzooecial tissue from the central axial region to the surface of the zoarium. In the axis the interzooecial tissue consists of loosely arranged vertically elongate vesicles, but in the outer part of the colony the vesicles are compressed, wider than high, and densely packed. The transitional zone between these two very contrasting tissue patterns is narrow, usually not exceeding 2 or 3 mm in width. An axial (transverse) section of F.? peculiaris reveals thin-walled zooecia, usually slightly polygonal in outline, that are interspersed between angular vesicles that vary greatly in size and shape.
Because of the nature of the interzooecial tissue observed in longitudinal section, we have some reservation in assigning this species to Fistulipora, as suggested by its specific name. An argument probably could be advanced to erect a new genus to receive these specimens, differing chiefly from Fistulipora in having compressed tightly packed vesicles in the outer part of the zoarium and loosely arranged vertically elongate vesicles in the axial region. This structural feature is not to our knowledge observed in any species of Fistulipora. Had our collections contained more forms displaying this structural character, particularly if such specimens could be assigned to more than one species, a new fistuliporoid genus would have been proposed to include them. We are reluctant, however, to erect a new genus based on three specimens that belong to a single species.
Locality--2.
Types--Holotype, IU 6800; paratypes, IU 6827 and 6828.
Genus ERIDOPORA Ulrich, 1882
ERIDOPORA BEILENSIS Perkins & Perry, n. sp.
Pl. 3, fig. 1-4.
External features--Zoarium a sheet-like growth, commonly encrusting, and slightly exceeding 1 mm in thickness. Zooecia pyriform or broadly ovate, not in physical contact, and having their maximum apertural diameters crudely aligned over considerable areas. Lunaria prominent, thickened, elevated, and consistently occupying analogous positions on narrower extremities of zooecial apertures. Irregularly distributed areas, approximately 1 mm in diameter and devoid of zooecia, represent maculae.
Tangential section--Zooecia pyriform or crudely subtriangular, less commonly ovate, never in physical contact, and in most areas having their axes of maximum zooecial diameter subparallel. Most commonly (19 of 44 measurements) three entire zooecia and part of a fourth in a 2-mm distance, measured nearly parallel to maximum zooecial diameter direction; commonly (12 of 44 measurements) four entire zooecia or (11 measurements) four complete zooecia and part of a fifth in this distance; and rarely (2 observations) three entire zooecia in 2 mm. Average maximum apertural diameter of zooecia, excluding wall thickness, 0.34 mm, ranging from 0.24 to 0.46 mm for 50 measurements and having modes (7 measurements each) of 0.34 and 0.37 mm. Lunarium thickened where well preserved, typically at narrower extremity of zooecial aperture, and usually occupying from one-third to nearly one-half of the entire apertural periphery. Average interzooecial distance, based on 50 measurements, 0.11 mm, ranging from 0.03 to 0.21 mm and having a mode (10 measurements) of 0.10 mm. Vesicles conspicuously smaller than zooecia, angular or subangular, and generally one or two, rarely three, between adjacent zooecia. Subcircular aggregations, irregular in outline and not exceeding 0.8 mm in maximum dimension, of angular vesicles represent maculae.
Longitudinal section--Zooecia subprostrate near base of zoarial growth, later becoming erect and meeting zoarial surface at right angles, and either lacking diaphragms or, less commonly, containing only one of these structures per zooecium. Interzooecial tissue composed of small vesicles, crescentic or subrectangular in outline, convex outward, and either haphazardly arranged or locally forming laterally imbricating columns. Maximum observed thickness of zoarial growth 1.1 mm.
Remarks--Eridopora beilensis, represented in our collections by five specimens, is related more closely to E. macrostoma described by Ulrich (1882, p. 137, pl. 6, fig. 2, 2a) than to any other species of the genus. The most conspicuous difference between these two species of Eridopora is in the size of the zooecia; average maximum zooecial diameter of E. beilensis is 0.34 mm as compared with 0.45 mm for E. macrostoma (Perry and Horowitz, in press). Further, interzooecial vesicles, especially as viewed in tangential section, are somewhat smaller in E. beilensis than in E. macrostoma.
The specific name alludes to the stratigraphic unit that furnished the original type material.
Locality--1.
Types--Holotype, IU 6801; figured paratype, IU 6802; unfigured paratypes, IU 6803-6805.
Family HEXAGONELLIDAE Crockford, 1947
Genus MEEKOPORA Ulrich, 1889
MEEKOPORA sp. indet.
Pl. 1, fig. 1.
Remarks--The collection includes a fragmentary zoarium that clearly can be assigned to the genus Meekopora, but excessive silicification has obliterated structural features in tangential section and in the outer part of the longitudinal section so that specific identification cannot be determined.
The zoarium is plainly bifoliate in longitudinal section, which clearly shows zooecia diverging outward from the mesotheca. Zooecia are nearly recumbent adjacent to the mesotheca and gradually assume a more erect attitude as they approach the zoarial surface. Vesicles, convex outward and crudely rectangular or crescentic in outline, separate adjacent zooecia a short distance outward from the mesotheca but are obliterated in the outer part of the zoarium by excessive silicification.
Locality--1.
Described specimen--IU 6806.
Order TREPOSTOMATA Ulrich, 1882
Family STENOPORIDAE Waagen and Wentzel, 1886
Genus STENOPORA Lonsdale, 1844
STENOPORA PAUCA Perkins & Perry, n. sp.
Pl. 5, fig. 5-7.
External features--Zoarium ramose, solid, bifurcating, the larger fragmentary specimen having length and width of 16 mm and 7 mm, respectively. Zooecia angular and displaying subangular to broadly ovate apertures. Acanthopores locally prominent but not observed elsewhere. Maculae sparse, irregularly distributed, approximately 1.5 mm in diameter, and composed of zooecia slightly above average in size and smaller cells, presumably mesopores.
Tangential section--Zooecia subangular to nearly angular and having broadly ovate apertures; most commonly (5 of 13 measurements) four entire zooecia in a 2-mm distance, measured nearly parallel to the length of the zoarium; number of zooecia in a 2-mm distance highly variable, ranging from three complete zooecia and part of a fourth (1 measurement) to six entire zooecia and a small part of a seventh (1 observation); zooecial walls homogeneously fibrous, lacking any evidence of a divisional line except in very local areas, and having an average thickness of 0.05 mm, ranging from 0.03 to 0.10 mm for 20 measurements but only two observations greater than 0.07 mm; mode (10 measurements) of zooecial wall thickness 0.05 mm; average maximum apertural diameter, based on 20 measurements, 0.38 mm, ranging from 0.30 to 0.46 mm and having a mode (5 measurements) of 0.34 mm. Acanthopores,prominent, as much as 0.07 mm in diameter and inflecting zooecia, as many as three associated with a single zooecium but elsewhere zooecia commonly devoid of acanthopores and having comparatively thin walls of concentrically arranged laminae surrounding a small central lumen. Except in maculae, mesopores very rare or absent. Maculae composed of zooecia somewhat above average in size and intercalated smaller structures, presumably mesopores, about one-half to two-thirds the size of intermonticular zooecia.
Longitudinal section--Zooecia erect in axis and having thin, straight or loosely flexuous walls of indiscernible microstructure; zooecia bend gradually and uniformly to zoarial surface, meeting it at nearly right angles. Mature zone virtually lacking in holotype (IU 6810) and thin (less than 0.08 mm) in paratype (IU 6811). In thin mature zone zooecial wall structure generally obscure, but in local areas composed of laminae wall structure moderately to broadly convex outward. Diaphragms and mesopores lacking throughout. Monilae questionably present in holotype (IU 6810) as a single horizon of wall thickenings located a short distance above plane of rejuvenation and extending obscurely through width of zoarium.
Remarks--Distinguishing features of Stenopora pauca are the prominent but sporadically developed acanthopores, the sparse development of monilae, and the considerable range in the number of zooecia observed in a 2-mm distance. The species is represented in our collections by two specimens.
Probably the closest form to this species is S. angularis, described by Ulrich (1890, p. 439-440, pl. 74, fig. 6-6b) from Keokuk rocks (Mississippian) at La Grange, Missouri. S. angularis has smaller zooecia and acanthopores, is a more robust form, and has better developed monilae than S. pauca.
The specific name alludes to the paucity of the monilae.
Locality--1, 3.
Types--Holotype, IU 6810; paratype, IU 6811.
Genus TABULIPORA Young, 1883
TABULIPORA GROVERENSIS Perkins & Perry, n. sp.
Pl. 5, fig. 1-4.
External features--Zoarium ramose, solid, the fragment having length and width of 15 mm and 9 mm, respectively. Acanthopores conspicuous, protuberant, and located at junction angles of zooecia. Mesopores not observed. Two areas, each approximately 1 mm in diameter, composed of zooecia about one-quarter above average size, may be maculae.
Tangential section--Zooecia angular to subangular and having homogeneously fibrous walls ranging in thickness from approximately 0.02 to 0.08 mm; most commonly (5 of 10 measurements) six zooecia and part of a seventh in a 2-mm distance, measured nearly parallel to length of zoarium; commonly (3 of 10 measurements) seven entire zooecia in this distance; and rarely (1 measurement) five complete zooecia and part of a sixth or (1 measurement) six entire zooecia in a 2-mm distance. Acanthopores very conspicuous, commonly inflecting zooecia, almost invariably at junction angles of adjoining zooecia, and consisting of a hollow lumen surrounded by dark-colored concentrically fibrous tissue clearly segregated from zooecial wall material; average acanthopore diameter 0.08 mm, ranging from 0.06 to 0.11 mm for 15 measurements. Mesopores lacking but small zooecia simulating mesopores very rarely observed. Evidence of monticules or maculae not seen.
Longitudinal section--Zooecia erect in axis and in this region having thin flexuous walls lacking discernible microstructure; in outer part of axial region zooecia arch gradually and intersect zoarial surface at right angles. In mature zone, averaging 1.4 mm in width, zooecial walls composed mainly of coalesced or isolated monilae clearly consisting of laminae moderately convex outward; locally laminae of monilae laterally continuous into centrally perforate diaphragms; four to eight monilae in a 0.5-mm distance measured along zooecial wall; markedly thinner segments of zooecial wall, separating isolated monilae, devoid of distinctive microstructure. Diaphragms centrally perforate, lacking in axial region, and usually spaced one to nearly two zooecial diameters apart in mature zone.
Remarks--The closest species to Tabulipora groverensis, which is represented in our collections by a single specimen, is probably T. carbonaria (Worthen) whose features are best described and illustrated by Ulrich (1890, p. 445, pl. 73, fig. 8, 8a) inasmuch as Worthen (Worthen and Meek, 1875, p. 526, pl. 32, fig. 5) did not illustrate the microstructural features of the species. Thus, our concept of T. carbonaria stems from Ulrich's work, although Foerste (1887, p. 85-86, pl. 7, fig. 8-8c) gave a brief description of the species.
T. groverensis differs from T. carbonaria in having somewhat larger zooecia, in lacking any suggestion of "a series of minute dark spots" (Ulrich, 1890, p. 445) in the divisional line between adjacent zooecia, in having more monilae per unit length measured along the zooecial wars in the mature region, and in having somewhat thicker zooecial walls as observed in tangential section.
The specific name alludes to the locality which furnished the type specimen of the species.
Locality--1.
Type--Holotype, IU 6809.
TABULIPORA REGULARIS Perkins & Perry, n. sp.
Pl. 4, fig. 1-6.
External features--Zoarium hemispherical in gross form, 15 mm in maximum width and 5 mm in greatest thickness, and having a concave base and a convex upper surface. Zooecia sharply angular, showing moderately thick walls, and displaying subangular to broadly ovate apertures. Acanthopores prominent, somewhat protuberant, numerous, and located at most junction angles of zooecia. Evidence of maculae or monticules wanting.
Tangential section--Zooecia commonly angular to subpolygonal, having irregularly rounded, broadly ovate, or subpolygonal apertures, and displaying moderately thick (usually 0.03 or 0.04 mm) homogeneously laminated zooecial walls; locally and rarely dark-colored, poorly defined irregularly and concentrically fibrous material appearing as small patches in zooecial walls; typically (10 of 30 measurements) six entire zooecia and part of a seventh or (9 of 30 measurements) seven whole zooecia in a 2-mm distance; less commonly (7 measurements) six complete zooecia or (4 measurements) five entire zooecia and a portion of a sixth in this distance. Acanthopores conspicuous, large (commonly 0.06 to 0.09 mm in diameter), usually circular but less commonly subquadrate, and typically three (14 of 28 measurements), but as many as five and as few as two, associated with each zooecium, almost invariably located at junction angles of adjoining zooecia, and commonly inflecting; acanthopore walls composed of distinct concentrically arranged fibers surrounding a hollow lumen. Mesopores not observed, but small subpolygonal immature zooecia relatively common. Neither monticules nor maculae noted.
Longitudinal section--Zooecia erect throughout zoarium, except near zoarial periphery where zooecia subprostrate near base and emanating outward from central basal region of growth. Zooecial walls moniliform except immediately above base of zoarium where walls thin and virtually devoid of definable microstructure; commonly five or six complete monilae in a 0.5-mm distance but as few as four and as many as seven in this distance; intermonilae wall segments distinctly thin; monilae clearly separate or partially coalesced within the same zoarium; monilae composed of distinct laminae gently convex outward, with outer laminae possibly continuous into diaphragms. Centrally perforate diaphragms usually spaced one-half to one and one-half zooecial diameters apart in holotype (IU 6807) but from one and one-half to nearly three zooecial diameters apart in paratype (IU 6808). Mesopores not observed.
Remarks--This species is based on two specimens. The paratype (IU 6808) seems to have a hemispherical zoarial form but the longitudinal section of the paratype indicates that the zoarium actually is a laminar expansion, having a maximum growth thickness of approximately 3 mm, that during its development has enveloped a foreign object, possibly a brachiopod shell. The colony of the holotype (IU 6807) has undergone local rejuvenation. For a short distance above the plane of rejuvenation the zooecia are inclined or subprostrate, and their thin walls lack monilae.
Distinguishing features of Tabulipora regularis are the large conspicuous acanthopores occupying many junction angles of zooecia, the distinctly moniliform zooecial walls as observed in longitudinal section, and the lack of mesopores.
The closest allies of T. regularis are T. plummeri, described by Moore (1929, p. 8, 10, pl. 1, fig. 21, 22) from the Wayland Shale (Pennsylvanian) of north-central Texas, and T. heteropora, originally reported by Condra (1902, p. 340-341, pl. 20, fig. 1, 2) from the "Coal Measures" near South Bend, Nebraska, and later redescribed by him (Condra, 1903, p. 43, pl. 4, fig. 7, 8) from the same horizon and locality.
T. regularis differs from T. plummeri Moore as the latter species lacks distinctly moniliform walls and possesses mesopores. T. regularis has more numerous acanthopores than T. heteropora (Condra) and further has more strikingly moniliform walls than the latter species. T. regularis differs from T. bullata, described by Coryell (1924, p. 180, pl. 39, fig. 5, 6) from the Wapanucka Limestone (Pennsylvanian) in the Stonewall Quadrangle of Oklahoma, as the latter species has eight or nine maculae in a 1-sq.-cm area and somewhat more numerous monilae.
The specific name directs attention to the uniform distribution or arrangement of the zooecia as observed in tangential section.
Locality--1.
Types--Holotype, IU 6807; paratype, IU 6808.
Order CRYPTOSTOMATA Vine, 1883
Family RHABDOMESIDAE Vine, 1883
Genus RHOMBOPORA Meek, 1872
RHOMBOPORA LEPIDODENDROIDES Meek, 1872
Pl. 3, fig. 5-7.
Rhombopora lepidodendroides Meek, 1872, Paleont. Eastern Nebraska, p. 141, pl. 7, fig. 2a-f; Keyes, 1894, Missouri Geol. Survey, v. 5, p. 35, pl. 33, fig. 4a, b; Condra, 1903, Nebraska Geol. Survey, v. 2, pt. 1, p. 99, pl. 6, fig. 2-4; pl. 7, fig. 1-12; Condra, 1903, Am. Geologist, v. 31, p. 22, pl. 2, fig. 1-11; Girty, 1908, U.S. Geol. Survey Prof. Paper 58, p. 153, pl. 31, fig. 17; Mather, 1915, Bull. Sci. Lab. Denison Univ., v. 18, p. 132, pl. 6, fig. 8, 9; Plummer and Moore, 1922, Texas Univ. Bull. 2132, p. 169, pl. 23, fig. 20, 20a, 21, 21a, 22; Coryell in Morgan, Okla. Geol. Survey Bull. 2, pl. 38, fig. 3-5; Sayre, 1931, Kansas Univ. Sci. Bull. 17, p. 92, pl. 1, fig. 6-8; Bassler in Moore, ed., 1953, Treatise on Invert. Paleontology, pt. G, Bryozoa, Geol. Soc. America and Univ. Kansas Press, p. G134, fig. 95, 4a-c.
Rhombipora lepidodendroides White in Wheeler, 1877, U.S. Geol. Survey, v. 4, Paleont., p. 99, pl. 6, fig. 5a-d.
Rhombopora lepidodendroidea Ulrich, 1884, Jour. Cincinnati Soc. Nat. Hist., v. 7, p. 27, pl. 1, fig. 1-1b; Foerste, 1887, Bull. Sci. Lab. Denison Univ., v. 2, pl. 7, fig. 3a, b.
External features--Zoarium ramose, solid, branching irregularly, the fragments typically 2 to 4 mm in diameter and as much as 20 mm in length. Zooecial apertures crudely aligned, usually ovate with greatest diameter parallel to length of zoarium, and surrounded by thick walls. Acanthopores large, black, somewhat protuberant, and usually observed at extremities of zooecial apertures. Definite evidence of maculae or monticules not observed.
Tangential section--Zooecial apertures broadly ovate and surrounded by concentrically fibrous peristome usually 0.02 to 0.03 mm in width; typically (27 of 84 measurements on 10 specimens) five zooecia and part of a sixth in a 2-mm distance, measured parallel to branch length; commonly (16 of 84 measurements) six entire zooecia or (16 of 84 measurements) five complete zooecia in this distance; somewhat less commonly (15 measurements) four complete zooecia and part of a fifth in 2 mm; and rarely (4 measurements) six entire zooecia and part of a seventh or (6 measurements) four complete zooecia in this distance; average maximum apertural diameter of zooecia, based on 100 measurements on 10 specimens, 0.24 mm, ranging from 0.20 to 0.29 mm; mode of maximum apertural diameter 0.24 mm; average interapertural distance, measured normal to long axes of contiguous zooecia and based on 100 measurements on 10 specimens, 0.17 mm, ranging from 0.09 to 0.25 mm; mode of interapertural distance 0.18 mm. Megacanthopores prominent, usually ranging in diameter from 0.06 to 0.10 mm, and consisting of a small lumen surrounded by obscurely fibrous concentric laminae; a megacanthopore commonly noted between successively aligned zooecia along branch length. Micracanthopores approximately seven to ten times more abundant than megacanthopores, dark in color, and consisting of a minute, poorly preserved lumen surrounded by dark, concentrically arranged laminae. Neither monticules, maculae, nor mesopores observed.
Longitudinal section--Zooecia thin walled in axis, straight in central axis, and diverging gently to zoarial surface in outer axial region; at base of well-marked mature region, zooecial walls thicken abruptly, and zooecia assume position normal to zoarial surface. Zooecial wall material composed of laminae moderately to gently convex outward. Diaphragms sparse, generally two or three per zooecium, one located near base of mature zone; diaphragms not observed in some zooecia. Mesopores not seen.
Remarks--As judged from our collections, Rhombopora lepidodendroides is the most abundant species in the Beil bryozoan fauna; it is especially common at localities 1 and 4. This species is widely distributed in rocks of Pennsylvanian age and has been reported from several stratigraphic horizons.
Locality--1, 2, 3, 4.
Types--Hypotypes: figured, IU 6812 and 6813; unfigured, IU 6814-6826.
Brown, W. G., 1956, Stratigraphy of the Beil Limestone, Virgilian, of eastern Kansas: Unpub. master's thesis, Kansas Univ., 180 p.
Condra, G. E., 1902, New Bryozoa from the Coal Measures of Nebraska: Am. Geologist, v. 30, no. 6, p. 337-359, pl. 18-25.
Condra, G. E., 1903, The Coal Measure Bryozoa of Nebraska: Nebraska Geol. Survey, v. 31 pt. 1, 162 p., 21 pl., 6 tables.
Coryell, H. N., 1924, Bryozoa, in Morgan, G. D., Geology of the Stonewall Quadrangle, Oklahoma: Oklahoma Bureau Geol., Bull. 2, p. 176-184.
Foerste, A. F., 1887, Flint Ridge Bryozoa: Denison Univ. Sci. Lab. Bull., v. 2, pt. 1, p. 71-88, 1 pl.
Moore, R. C., 1929, A bryozoan faunule from the upper Graham Formation, Pennsylvanian, of north-central Texas: Jour. Paleontology, v. 3, p. 1-27, 3 pl., 3 fig.
Moore, R. C., 1936, Stratigraphic classification of the Pennsylvanian rocks of Kansas: Kansas Geol. Survey, Bull. 22, 256 p., 12 fig. [available online]
Moore, R. C., and Dudley, R. M., 1944, Cheilotrypid bryozoans from Pennsylvanian and Permian rocks of the Midcontinent Region: Kansas Geol. Survey, Bull. 52, pt. 6, p. 229-408, 48 pl.
Perry, T. G., and Gutschick, R. C., 1959, Bryozoans from the Amsden Formation, southwest Montana: Jour. Paleontology, v. 33, no. 2, p. 313-322, pl. 46-48.
Perry, T. G., and Horowitz, A. S., in press, Bryozoans from the Glen Dean Limestone (middle Chester) of southern Indiana and Kentucky: Indiana Geol. Survey [Bulletin 26, 1963].Rudwick, M. J. S., 1961, The anchorage of articulate brachiopods on soft substrata: Paleontology, v. 4, pt. 3, p. 475-476.
Sverdrup, H. U., Johnson, M. W., and Fleming, R. H., 1942, The Oceans: Prentice-Hall, New York, 1087 p., 265 fig., 121 tables.
Ulrich, E. O., 1882, American Paleozoic Bryozoa: Cincinnati Soc. Nat. Hist. Jour., v. 5, p. 121-177, 5 pl.
Ulrich, E. O., 1890, Paleozoic Bryozoa: Illinois Geol. Survey, v. 8, pt. 2, sec. 6, p. 285-688, pl. 29-78, 18 fig.
Worthen, A. H., and Meek, F. B., 1875, Descriptions of invertebrates: Illinois Geol. Survey, v. 6, sec. 2, p. 491-532, pl. 18-25.
A higher-resolution version of this plate is available as an Acrobat PDF file.
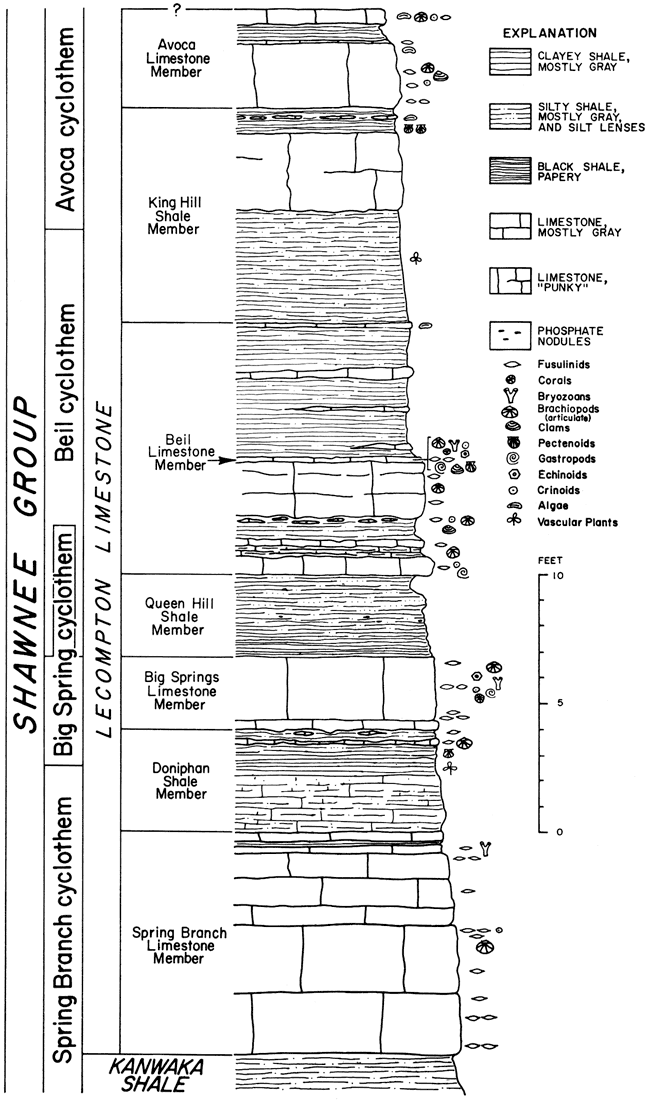
(Figure 2, X60; other figures X20.)
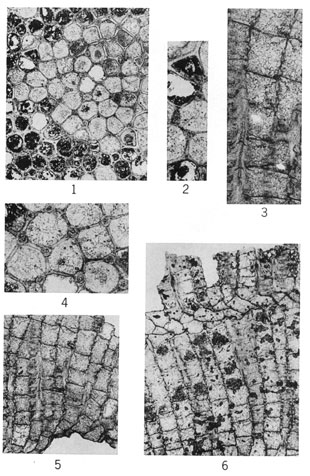
Figure 1--Meekopora sp. indet. Longitudinal section showing zooecia diverging from central mesotheca and adjoining vesicles; outer part of colony excessively silicified (IU 6806).
Figures 2-6--Fistulipora decora (Moore and Dudley); hypotypes. 2, Tangential section showing thickened lunaria (IU 6781). 3, Longitudinal section displaying compresses and vertically elongate vesicles (IU 6782). 4, Tangential section exhibiting monticule (IU 6780). 5, Typical tangential section (IU 6783). 6, Longitudinal section showing normal development of interzooecial tissue (IU 6783).
A higher-resolution version of this plate is available as an Acrobat PDF file.
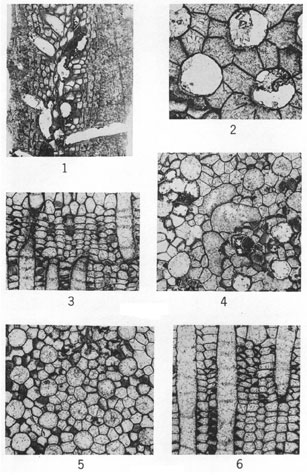
(Figure 2, X60; other figures X20.)
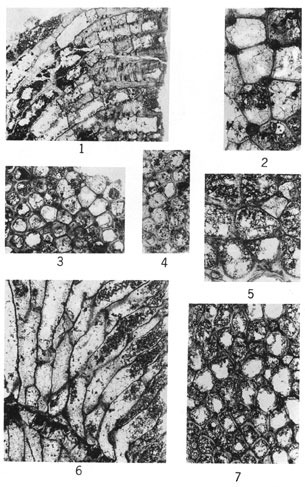
Figures 1-6--Fistulipora? peculiaris Perkins & Perry, n. sp.; holotype, IU 6800. 1, Tangential section. 2, Tangential section showing thick-walled zooecia and adjoining vesicles. 3, Axial (transverse) section displaying thin-walled zooecia interspersed amid irregularly developed vesicles. 4, Tangential section. 5, Longitudinal section near zoarial surface (to right) showing nature of interzooecial tissue and paucity of diaphragms in outer part of zooecia. 6, Longitudinal section in axial region showing zooecia and loosely arranged vertically elongate vesicles.
A higher-resolution version of this plate is available as an Acrobat PDF file.
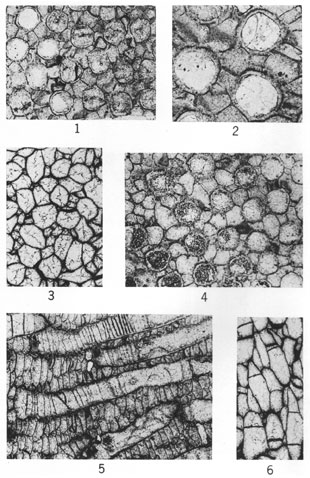
(Figures 1 and 5, X60; other figures X20.)
Figures 1-4--Eridopora beilensis Perkins & Perry, n. sp. 1, Tangential section showing nature of lunaria on pyriform zooecia and interzooecial tissue (holotype, IU 6801). 2, Longitudinal section (holotype, IU 6801). 3, Tangential section (holotype, IU 6801). 4, Tangential section (paratype, IU 6802).
Figures 5-7--Rhombopora lepidodendroides Meek; hypotypes. 5, Tangential section showing megacanthopores, micracanthopores, and character of zooecial wall (IU 6812). 6, Longitudinal section (IU 6813). 7, Typical tangential section (IU 6813).
A higher-resolution version of this plate is available as an Acrobat PDF file.
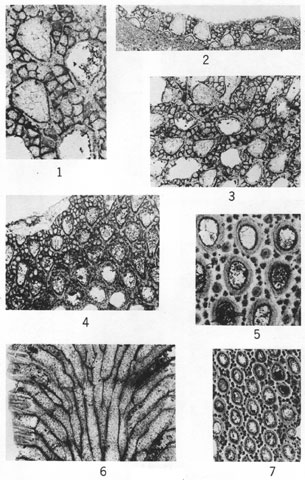
(Figures 2-4, X60; other figures X20.)
Figures 1-6--Tabulipora regularis Perkins & Perry, n. sp.; holotype, IU 6807. 1, Tangential section showing uniform size and arrangement of zooecia. 2, Tangential section. 3, Longitudinal section revealing acanthopore (lower left), a centrally perforate diaphragm (center), and monilae. 4, Tangential section showing nature of acanthopores and zooecial wall structure. 5, Longitudinal section. 6, Longitudinal section showing thin-walled inclined zooecia immediately above plane of rejuvenation.
A higher-resolution version of this plate is available as an Acrobat PDF file.
(Figures 2 and 5, X60; other figures X20.)
Figures 1-4--Tabulipora groverensis Perkins & Perry, n. sp.; holotype, IU 6809. 1, Longitudinal section showing centrally perforate diaphragms and isolated and coalesced monilae. 2, Tangential section displaying prominent acanthopores. 3, Tangential section showing distribution of acanthopores and variation in zooecial wall thickness. 4, Tangential section.
Figures 5-7--Stenopora pauca Perkins & Perry, n. sp.; holotype, IU 6810. 5, Tangential section. 6, Longitudinal section exhibiting thin zooecial walls and poorly developed monilae above plane of rejuvenation. 7, Tangential section.
Kansas Geological Survey, Geology
Placed on web Sept. 11, 2008; originally published in Nov. 1962.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/157_5/index.html