
|
Original published in D.F. Merriam, ed., 1964, Symposium on cyclic sedimentation: Kansas Geological Survey, Bulletin 169, pp. 541-563 | |
Peabody Museum, Yale University, New Haven, Connecticut
Geographically the assemblages occupied elongate northeast-trending areas in what was apparently a northern embayment of the waning interior Cretaceous sea. Groups of assemblages show progressive southward shift concurrent with encroachment of littoral facies from the northeast. Finer patterns of distribution resulting from local abundances of nondominant species and from differing local associations of species are characteristic of the individual layers.
The distribution patterns, together with the characteristic dominance of a few species and the fact that dominant associations are repeated in the sequence both locally and regionally, contribute to the marked general resemblance between the fossil assemblages and modern marine bottom communities. The preservation of successive accumulations of shells showing patterned distribution over much the same large area is taken as strong indication that the assemblages resulted from recurrent mass mortalities with relatively little disturbance by current action before burial. The cause of the mortalities is not known. Sparse evidence seems to favor burial under conditions of excessive turbidity and lowered salinity, possibly brought about by repeated influx of sediment-charged fresh water from rivers in flood.
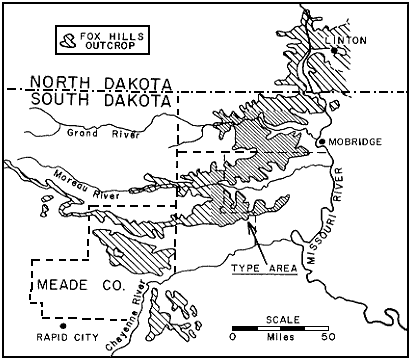
Figure 1--Principal outcrop of Fox Hills Formation in Dakota showing location of type areas.
The richly fossiliferous concretion layers in the marine part of the formation have been the most intriguing but also the most perplexing aspect of the work. With the details of the stratigraphy known and a start made on distributional studies of the fossils, the idea that the fossil assemblages resulted from recurrent mass killings grew from an impression to a strong working hypothesis. This hypothesis permits explanation of distributional peculiarities as vestiges of natural ecological arrangements, and in this context a number of facts make sense that are otherwise exceedingly difficult to explain. The reason for presenting the idea at this stage of the work is to emphasize that it can be substantiated or disproved only by detailed studies on the distributional and ecological aspects of the paleontology of similar interior Cretaceous terrains--an approach still much neglected despite the apparent popularity of paleoecological studies.
The basal Trail City Member of the Fox Hills averages about 90 feet in thickness and consists chiefly of clayey silt that becomes more sandy upward. Its two most conspicuous characteristics are the general, but not universal, lack of bedding structures and the prominent layers of limestone concretions, many of which are abundantly fossiliferous. Blebs and stringers of silt and clay and bored and contorted laminae indicate post-depositional working of the sediment by organisms; details of the structure are identical to those of recent sediments that have been so worked. The presence of distinctive assemblages of great numbers of fossils in successive layers, or groups of layers, is an outstanding feature of the fossiliferous concretions. The successively different fossiliferous layers, together with certain persistent layers of dominantly barren concretions, thin layers of yellow-weathering jarositic silt, and glauconitic silt or sand layers form a framework of key beds that can be carried throughout the extent of the member and for some distance into equivalent lateral facies.
Figure 2--Subdivisions of Fox Hills Formation in type area.
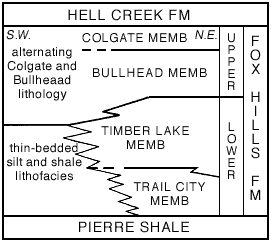
Together the Trail City and Timber Lake Members form a distinctive lower marine part of the Fox Hills Formation. Northeastward in the Missouri Valley area the sandy lithofacies thickens at the expense of the underlying clayey silt; the contact between the two occurs progressively lower in the section. In the northeasternmost outcrops in North Dakota east of the Missouri River, only a few feet of Trail City lithology remain. Southwestward in the type area both members grade laterally into a succession of sparsely fossiliferous, thin-bedded silty shale and clayey sand and silt. This lithofacies superficially resembles the underlying Pierre Shale, but is considerably more silty.
Succeeding the Timber Lake Member is a conspicuous unit, between 35 and 110 feet thick, of thinly interbedded shale, silt, and sand that was long referred to as the "banded beds," but more recently named the Bullhead Member. A sparse and restricted marine fauna is locally present in the lower part of the member; but higher in the section fossils are rare except for otoliths.
The Colgate Member overlies the Bullhead throughout much of the area. It consists of fine- to medium-grained, dirty, friable sandstone with considerable interstitial white clay. It weathers to characteristic gray-white fluted outcrops. Generally the Colgate is about 20 feet thick, but may be considerably thicker or entirely absent from the sequence. Large spheroidal, brown-weathering concretions of lime-cemented sand are characteristic features. The most common fossils in the member are reefs and coquinas of oysters and a brackish-water fauna dominated by the bivalve Corbicula. Locally this fauna contains a few marine elements and, where its usually gradational contact with the Bullhead Member is channeled, there are local accumulations of plant and dinosaur remains.
The Bullhead and Colgate Members form a distinctive upper part of the Fox Hills Formation characterized by restricted marine and brackish-water faunas. The two lithofacies are so closely interrelated that it is not practical to separate them over all of the type area. The upper contact of the Fox Hills with the Hell Creek Formation is arbitrarily taken at the first bed of lignite or lignitic shale. This contact is more or less traditional and generally satisfactory, as it marks the beginning of dominantly continental sediments.
The rich molluscan faunas of the type Fox Hills and equivalent beds in the Missouri Valley area of the central Dakotas are the youngest marine Cretaceous faunas known in the interior region of the United States and Canada. These faunas disappear westward and southwestward within the type area. As the formation is traced along the outcrop to the southwest from the type area, a major change in facies takes place within about 50 miles which completely transforms the Fox Hills into a heterogeneous sequence of dominantly sandy nonmarine beds containing such atypical sediments as lignite beds and variegated silty clays. This complex and little known Fox Hills terrain occupies a large area of outcrop in and south of Meade County, between the type area and the Black Hills (Fig. 1) . Here the formation rests about 250 feet lower in the section relative to the top of the range zone of Baculites clinolobatus than does the base of the Fox Hills in the type area. Although the details are unknown, the gross relationships indicate that this brackish and continental phase of the Fox Hills was a large delta or series of coalescing deltas formed by major drainage into the Cretaceous sea from the north or northwest. The delta complex was established while Pierre Shale was deposited in the area of the type Fox Hills, and it marks the westward limit of the sea in this part of the interior region during the deposition of the type Fox Hills. There is also evidence of regional shoaling to the north and northeast within the Fox Hills of the Missouri Valley region, and it appears that sediments of the type Fox Hills were deposited in a northern embayment of the Late Cretaceous sea which lay between deltaic areas to the west and east or northeast.
Marine fossils in the Fox Hills Formation occur chiefly as accumulations in limestone concretions, a mode of occurrence that is restricted to the lower marine portion of the formation, in particular to the Trail City Member and the marine biofacies of the Timber Lake Member. In the Trail City Member the abundantly fossiliferous concretions are concentrated in the lower 50 feet of the member; both here and elsewhere in the member, fossils are exceedingly rare in the surrounding clayey silt and clayey sand. In the Timber Lake Member abundantly fossiliferous concretions are limited to the lower 60 feet of its marine biofacies, but scattered fossils are not uncommon in the surrounding sandy sediments. Fossils in the Tancredia-Ophiomorpha biofacies of the Timber Lake and those of the restricted faunas of the upper part of the Fox Hills rarely occur in limestone concretions.
The concretions are commonly distributed in continuous layers. In some the individual concretions are closely spaced; in others they are widely spaced. Locally concretions are scattered through an interval of several feet which when traced laterally thins to a single concretion layer. The extent of the individual concretion layers ranges greatly. Some persist throughout the fossiliferous Trail City Member and extend many miles beyond it into the relatively barren, thin-bedded lithofacies to the southwest. Other layers are of local extent within a very small part of the area.
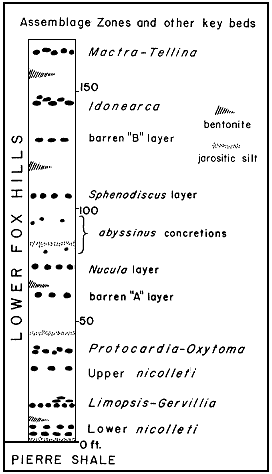
Although none of the concretion layers in the lower part of the Fox Hills appears to be completely barren, several are commonly unfossiliferous. Of the layers shown in Figure 3, those indicated as barren A, Nucula, and barren B, commonly lack fossils. The Nucula layer carries a very sparse but distinctive faunule, but the barren A and barren B layers are even less fossiliferous and have yielded only a very few scattered ammonoids.
Figure 3--Generalized composite succession of as semblage zones and other key beds in marine biofacies of lower Fox Hills Formation.
The layers of richly fossiliferous limestone concretions in the marine lower Fox Hills sequence of the type area are one of its most conspicuous and interesting features; the same can be said for a number of other Upper Cretaceous terrains in which similar fossiliferous concretions occur. Many geologists have described these layers, collected their fossils, and speculated on the origin of the concretions, but James Todd appears to have been the first of very few to be impressed by the fact that each fossiliferous layer commonly contains a distinctive faunal assemblage. His appreciation of the biological implications of this peculiar distribution is apparent in the following excerpt from his remarks on the origin of the fossiliferous concretions in the type Fox Hills (Todd, 1910, p. 31).
Very commonly they are crowded with fossils, which in the same concretions are usually only one or two species, as though they had been isolated colonies in the surrounding mud flat. Some of the shallow masses are barely covered with the calcareous rock. In other cases they are very thickly invested. The colony theory seems as probable as any, but why should they have a thickness nearly equal to their breadth? No shells are found in the surrounding clay or sand. The shells do not seem to be horizontally laid as in stratified rocks. Is it possible that some strong currents broke them out of stratified patches and rolled them away? This does not seem likely when we remember the fine character of the material in which they lie; nor does it agree with the uniformity of individuals in the same concretion; nor with the difference of kinds in different concretions. That they have grown by concretion is evident, but the original blocking out and dispersion is the puzzle.No progress toward the solution of Todd's "puzzle" has been made in the more than half century since these observations. His clear separation of concretion formation from the origin of the fossil accumulations and his implication that the latter is the more significant problem are points that deserve re-emphasis; investigators commonly make speculative remarks about the concretions, but usually take the fossil masses for granted. However, the fossil assemblages and concretions are too closely associated to ignore one completely in discussing the other. A more realistic perspective is to regard concretion formation as a related, post-depositional aspect of the broader problem of the origin and preservation of the fossil accumulations.
This is not the place for a thorough discussion of what is known about the origin of the Fox Hills concretions, but certain aspects of the subject are pertinent to the pattern of fossil distribution. The principal role of the concretions relative to the fossil accumulations has been that of a preserving medium. The fact that fossils do occur, though rarely, in the sediments outside of concretions indicates that the peculiar distribution of the fossils in localized masses or clusters is not merely the result of localized preservation resulting from selective calcification of parts of widespread shell beds. The scattering of fossiliferous concretions among unfossiliferous concretions, which is a pattern prevalent in all layers, supports this contention, as do the rare occurrences of single fossils and small clusters of shells about which no concretion has formed.
Thin sections of the concretions show that they consist of calcium-carbonate cemented sediment preserving structures similar to those in the surrounding sediment. All features point to their formation after the burial of the organisms but before compaction. Fossils in the concretions show no flattening in the horizontal plane, but those extending beyond the concretion and the few that occur entirely outside in the clayey silt are generally crushed.
As there is no way of making a reasonable estimate of the rate of sedimentation, the knowledge that the concretion formed before compaction cannot be used to determine how long it took the concretion to form. Preservation of perishable structures such as the ligaments of bivalves, occasional leaves, and thin carbonaceous aptychi of scaphitid ammonoids suggests that concretion formation took place within a relatively short time after burial. Calcareous concretions in marine sediments of relatively recent origin are of little help in estimating the time involved in concretion formation. In a study, now in progress, of fossiliferous limestone concretions from Pleistocene deposits in Long Island Sound, the radiocarbon age of included scallop shells is 4600 ± 90 years B. P. Although this is, to my knowledge, one of the youngest marine concretions yet dated, the figure probably has little bearing, even as an outside limit, on the rate of formation of concretions of this type. If the precipitation of calcium carbonate is brought about by the products of decomposition of the organisms as is commonly held (Burt, 1932; Weeks, 1953; and others), but as yet incompletely explained, concretion formation must have taken place at most within tens rather than thousands of years.
The barren concretions do not necessarily weaken the hypothesis that the decomposition of organic matter created local environments chemically favorable to the formation of calcium carbonate. Shelled molluscs, which make up practically the entire macrofossil content of the fossiliferous concretions, were only a part of the biomass and most likely were appreciably less in aggregate volume than the combined marine plants and soft-bodied animals that lacked easily preservable parts. Throughout the marine portion of the type Fox Hills it is a conspicuous fact that the area of most abundant concretions coincides geographically with the area of most abundant fossils. Only preliminary geochemical work has been done on the Fox Hills concretions, and although this indicates as strong a concentration of amino acids in the "barren" limestone concretions as in the fossiliferous ones, it is obvious from lateral changes in the physical properties and calcareous content of concretions and from peculiarities in both their stratigraphic and geographic distribution that their mode of origin is far more complex than can be explained by any simple relationship to accumulations of organic matter alone. (Note: As part of a senior project in 1962-63 at Yale University by Edward Gilfillan.) The point pertinent to this paper, however, is that the original distribution of organic matter apparently had a great deal to do with concretion formation; but concretion formation was subsequent to fossil distribution and did not influence it other than to insure local preservation of the fossils in an uncompacted state.
Assemblage Zones
The fossiliferous concretion layers, either individually or in groups of several layers, contain distinctive assemblages of fossils. In the Trail City Member one or two species of pelecypods commonly dominate a given assemblage, locally constituting as much as 95 percent of the total number of specimens in a concretion. Less commonly the dominant species is an ammonoid or a gastropod. The assemblages in the Timber Lake Member are usually characterized by more variety within the individual concretions, but the dominance of one or two species is still marked. For the lower Fox Hills as a whole, the pelecypod fauna is relatively uniform in composition and the majority of species range throughout the sequence. The conspicuous faunal difference from one fossiliferous concretion layer or group of layers to the next results from differences in the abundant species, the majority of which are long-ranging. Accumulations of this type are assemblage zones (Am. Comm. Strat. Nomenclature, 1961, Art. 21) and are named for one or more of the dominant elements in the assemblage.
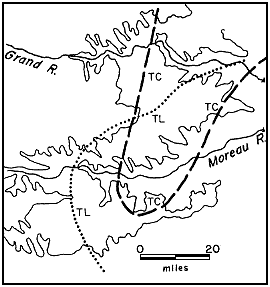
The sequence of assemblage zones in the lower Fox Hills is shown diagrammatically in Figure 3. Not all of the assemblages shown in this composite illustration are necessarily found at any one place. The lower Trail City assemblages all have approximately the same general pattern of distribution occupying, to somewhat different degrees, a northeast-trending lobate part of the type area that terminates in the vicinity of the Moreau River (Fig. 4, 5, 6, 7). The Timber Lake assemblages overlap those of the Trail City in the eastern part of the Moreau-Grand divide, but they extend farther to the south and southwest be yond the limits of the Trail City assemblages (Fig. 4).
Figure 4--Approximate limits of abundantly fossiliferous marine assemblages in Trail City (dashed line) and Timber Lake (dotted line) Members of Fox Hills Formation.
Trail City Assemblages
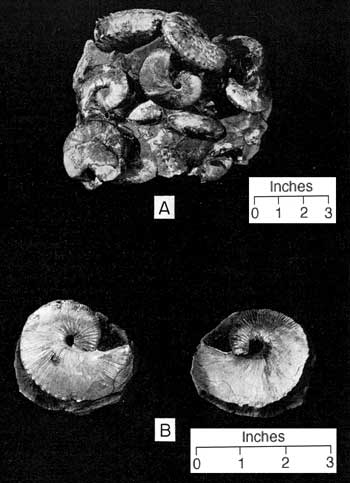
For some 200 feet beneath the widespread and abundantly fossiliferous succession of assemblage zones in the lower 50 feet of the Trail City Member the drab gray, finely silty Pierre Shale is practically devoid of macrofossils except for scattered linguloid brachiopods and rare ammonoids. The base of the Fox Hills Formation is marked by both an appreciable and fairly abrupt increase in silt and by the appearance of the concretions of the basal assemblage zone. Within the individual concretions of the Trail City assemblages, the fossils occur in great numbers oriented at random (Fig. 8, 9, 10). Preservation is excellent. Pelecypods commonly are preserved with both valves in position or agape, some retaining ligament, and complete specimens of ammonoids are a commonplace. In fresh concretions the bivalve shells usually show their original luster and the ammonoids a beautiful iridescent coloration.
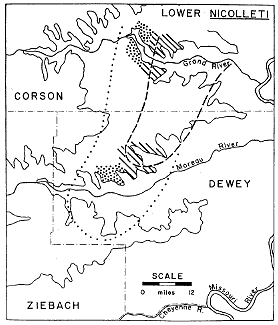
Figure 5--Distribution of Lower nicolleti assemblages. Dashed line encloses, area of abundant large accumulations of nicolleti, dotted line scattered small concretions. Stippled areas indicate Drepanochilus accumulations, cross-hatched areas abundant large Gervillia.
Light-gray weathering, ovoid, to flat-ovoid, calcareous concretions consisting of thick, punky jackets of calcareous silt and harder, silty, gray limestone cores up to 18 inches in diameter are characteristic of this zone (Fig. 8). Beginning at or a short distance above the base of the formation, the concretions are distributed in two to five layers spread through an interval that ranges from 2 to 15 feet in thickness. Concretions containing large numbers of the ammonoid Scaphites (Hoploscaphites) nicolleti, and a less conspicuous fauna, chiefly other molluscs, are scattered throughout the concretion layers. The number of concretion layers, concretion density in the layers and relative number of fossiliferous concretions varies from place to place. The fossiliferous concretions are substantially outnumbered by the unfossiliferous, but exposures adequate for counts are too few in relation to the size of the area, and the counts themselves too variable to permit a meaningful estimate of the ratio.
The Lower nicolleti zone exhibits two principal phases, a thinner eastern phase characterized by several layers of jacketed concretions, and a thicker western phase characterized by numerous small spherical concretions scattered among some of the larger jacketed kind. The abundantly fossiliferous nicolleti concretions disappear westward about where the latter phase begins; nicolleti and a few other ammonoids occur rarely as single specimens in the small concretions. Westward beyond this latter phase concretions at the horizon of the Lower nicolleti zone are few and barren.
The principal features of faunal distribution in the Lower nicolleti zone are shown in Fig. 5. Certain conspicuous local variations indicate a more complex distribution pattern of molluscs other than the ubiquitous nicolleti. In the uppermost concretion layer of the zone in restricted areas of outcrop along both the Moreau and Grand Rivers, small concretions dominated by the gastropod Drepanochilus americanus occur. Just to the east of both of these areas large specimens of the bivalve Gervillia are noticeably more abundant than elsewhere in the zone.
In addition to these more obvious distributional features the dominant bivalves associated with the masses of nicolleti in individual concretions differ from place to place. In many concretions the chief bivalve accompanying nicolleti is Inoceramus fibrosus, which is not very common in the Fox Hills except in association with nicolleti. Speden (oral communication) has found that at least four different associations of bivalves occur in the Lower nicolleti Assemblage Zone but, as is true of this type of association at all levels, not enough is yet known about the probable life habits of the fossil bivalves to judge the ecological validity of the associations.
The total geographic extent of the abundant Lower nicolleti accumulations is unknown, but they have been found as far to the northeast as Linton, North Dakota, 90 miles from their southwesternmost occurrence in the type area of the Fox Hills Formation (Fig. 1). A conservative estimate of the area they cover is 1,500 square miles. Fossiliferous concretions have been calculated to occur locally with a frequency of as high as one in every 100 square feet; this probably approaches maximum local density. There is no quantitative basis for arriving at an average density figure for the entire area. From collecting experience in the type area the frequency of one in every 2,500 square feet would be a conservative estimate. Individual fossiliferous concretions may yield from as few as 10 to as many as 45 specimens of ammonoids. In a count of approximately 1,800 specimens, 14 out of every 15 were the species nicolleti. Even using the most conservative figures the number of specimens of nicolleti in the area covered by the assemblages is in the tens of millions. In one 20 square-mile area along the Grand River, all outcrops sampled indicate a density of appreciably more than one fossiliferous concretion in every 2,500 square feet, but even taking this figure and an average of 20 nicolleti specimens per concretion the estimated number of specimens in the 20 square-mile area is more than 20 million. Consequently, if the buried area between outcrops is as prolific as the outcrops of the Lower nicolleti zone, which we must presume is so, the number of ammonoids can be measured in the hundred millions. No matter how one manipulates the figures available, the accumulation is impressive; yet of all the principal assemblage zones, the Lower nicolleti zone has the fewer fossiliferous concretions.
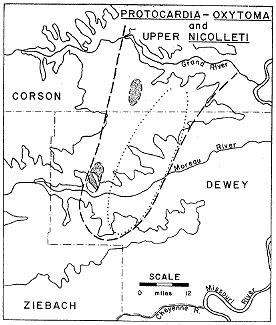
Figure 7--Distribution of Protocardia-Oxytoma (within dashed line) and Upper nicolleti (within dotted line) assemblages. Stippled areas indicate Lucina concentrations in Protocardia-Oxytoma zone.
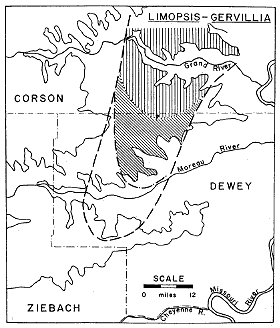
From as little as two to as much as 13 feet of barren clayey silt lie between the highest concretion layer of the Lower nicolleti zone and the basal concretion layer of the Limopsis-Gervillia Assemblage Zone. This persistent basal layer is in some places the only concretion layer in the zone, but in other places it is one of several layers in a zone ranging from 3 to 10 feet in thickness.
The basal layer consists of hard, blue-gray, spherical to ovoid concretions that commonly weather a characteristic rusty brown color. Many of these are crowded with the small bivalve, Limopsis striatopunctata Evans and Shumard (Fig. 9) and an assortment of less numerous molluscan species the more common of which are Gervillia recta? Meek and Hayden and a small bivalve that Meek called Nemodon sulcatinus. Ammonoids, though few in number relative to bivalves, are numerous and considerably richer in variety than in the Lower nicolleti Assemblage Zone.
Figure 9--Concretions from Limopsis-Gervillia Assemblage Zone. A, Weathered core of a Limopsis concretion. B, Fresh fragment of a Limopsis concretion (approx. x 2/3). C, Part of a Gervillia concretion oriented as found in place. A larger version of this figure is available.

Distributional features of the Limopsis-Gervillia Assemblage Zone in the type area of the Fox Hills are shown in Figure 6. It covers approximately the same area as the Lower nicolleti zone, although its fossil accumulations disappear several miles beyond the western limit of the large accumulations in the nicolleti zone. Outside of the type area it too extends northeastward to the vicinity of Linton, North Dakota.
The number of individual specimens of Limopsis in the lower concretion layer throughout its area of extent must number in the billions for there are not only a greater number of fossiliferous concretions per unit area than in the nicolleti zone, but also many times the number of individual specimens in a single concretion. The fossiliferous Gervillia concretions do not cover as large an area as the Limopsis layer, but they are much more heavily concentrated and more numerous than Limopsis concretions. At one locality where a layer is exposed at the surface of a small bench about 300 square feet in area, 17 fossiliferous concretions were counted, each of which contained over 100 specimens, counting two valves per specimen. This same concretion density in one layer over 1 square mile would amount to well over a hundred million specimens.
Upper nicolleti Assemblage Zone
The interval of beds between the Limopsis-Gervillia Assemblage Zone and the next major zone above it--the Protocardia-Oxytoma Assemblage Zone--ranges from 5 to 30 feet in thickness. Throughout the western part of the outcrop area of the Fox Hills in the Grand River Valley the interval is less than 15 feet in thickness and lacks persistent concretion layers. Throughout the eastern part of the type area it is over 15 feet thick and contains a rather poorly defined layer of widely spaced, ovoid, hard, dark-blue limestone concretions some of which contain accumulations of Scaphites (Hoploscaphites) nicolleti. These concretions, which make up the Upper nicolleti Assemblage Zone vary in position from 2 to 12 feet below the lowest concretions in the Protocardia-Oxytoma Assemblage Zone.
As the distribution map indicates (Fig. 7), the Upper nicolleti zone is more restricted geographically than the other zones in the succession; it has not been found outside the type area of the Fox Hills. In the southwestern part of the type area it changes laterally into a sparsely fossiliferous zone with small scattered concretions containing a few ammonoids, pelecypods (chiefly Oxytoma), and less commonly but in unusual numbers considering their scarcity in the interior Cretaceous, the gladii of coleoids.
The Upper nicolleti assemblages contain other ammonoids including representatives of the Discoscaphites conradi (s.l.) complex. The most common bivalve is Inoceramus fibrosus. Fossiliferous concretions of the Lower and Upper nicolleti zones are readily distinguished by their stratigraphic position, marked difference in the lithology of concretions, and absence or presence, respectively, of certain members of the conradi complex. Nevertheless, fossils of the two assemblages are remarkably similar and in dealing with weathered concretions in float it can be very difficult to tell them apart. This is particularly true where both zones locally have the nicolleti-fibrosus association, and no ammonites other than nicolleti are present.
Protocardia-Oxytoma Assemblage Zone
This uppermost zone in the succession of highly fossiliferous assemblage zones in the lower Trail City ranges from a single layer of concretions to an interval locally as much as 10 feet thick, containing scattered concretions. It lies approximately in the middle of the Trail City Member varying locally in position from 30 to 55 feet above its base.
The Protocardia-Oxytoma Assemblage Zone is associated with the first appearance of an appreciable amount of fine-grained sand in the Fox Hills Formation. Within the type area the sand thickens to the east and northeast and grades into clayey silt to the south and west, its maximum thickness in the area is about 12 feet. The concretions are hard, dark blue-gray limestone commonly surrounded by semi-indurated rusty-weathering jackets where the matrix is sandy; the cores range from 6 to 15 inches in diameter and are spherical to flat-ovoid in shape.
Two species of bivalves, Protocardia sub-quadrata (Evans and Shumard) and Oxytoma nebrascensis (E. and S.) commonly occur in about equal quantity, each far outnumbering individuals of the other species in the varied fauna (Fig. 10). Their common mode of occurrence is in clusters of one or the other species, and a single concretion may have a mass of Protocardia in one part and of Oxytoma in another. Oxytoma, an epifaunal bivalve, is the more gregarious and commonly monopolizes individual concretions. The fauna of the Protocardia-Oxytoma assemblage is the most varied in the succession of assemblage zones; ammonoids are plentiful and concretions in which they are a numerically significant part of the assemblage are not uncommon.
Figure 10--Concretions from Protocardia-Oxytoma and Idonearca Assemblage Zones. A, Fragment with Protocardia. B, Fragment with Oxytoma. C, Part of a Idonearca concretion. A larger version of this figure is available.

Although the Protocardia-Oxytoma Assemblage Zone is more abundantly fossiliferous than any other in the sequence, excepting the layers with Gervillia, it would be exceedingly difficult to estimate abundances of the dominant species because the fauna is more varied and the dominants do not generally monopolize the fossiliferous concretions.
Stratigraphic Constancy
The individual assemblage zones and their component layers hold the same stratigraphic position relative to one another throughout their geographic extent. The observer mindful of ecology is easily persuaded by the faunal distinctiveness of the individual assemblages, their lateral continuity without break, and their relatively limited geographic distribution within the region, that each layer of accumulation was formed simultaneously over its entire area of distribution. The areas of accumulation in both the Trail City and Timber Lake Members appear to follow the environmental strike, and there is no reason to suspect that any of the individual layers might be time-transgressive.
Other key beds in the lower 50 feet of the Trail City conform to the fixed stratigraphic pattern of the assemblage zones. Two persistent jarositic silt layers, one at the base of the Trail City, the other a short distance above the Protocardia-Oxytoma Assemblage Zone, maintain their positions relative to the concretion layers, but their relation to time planes cannot be demonstrated as nothing is known about their origin.
Consistent maintenance of position of the assemblage zones relative to bentonite beds would be strong indication of the contemporaneity of individual layers; the evidence of such a relationship is suggestive but incomplete. Although several bentonite layers are present in the Fox Hills, they are preserved chiefly in the relatively unfossiliferous lithofacies of interbedded thin layers of silt and silty clay into which the Trail City and Timber Lake Members grade to the west and southwest. Traced eastward from this thin-bedded lithofacies, the bentonite layers begin to disappear where the thin bedding gives way laterally to predominantly mixed sediment in the marginal parts of the area rich in fossil accumulations. Here, there is abundant evidence that the ash falls were dispersed in the sediment during or soon after deposition by the mixing action of burrowing organisms.
Some overlap of bentonite layers and fossil accumulations exists in areas of gradation between the two lithofacies, and the bentonites consistently hold their position relative to the layers of fossil accumulations. Moreover, within the thin-bedded lithofacies the bentonites lie parallel to layers of barren concretions which can be traced laterally into specific assemblage zones. Together, these parallel relationships can be traced for over 12 miles along the Moreau River, obliquely across the environmental strike of the lower Trail City Member.
The thickness of beds between individual assemblage zones and concretion layers varies within the type area of the Fox Hills. The variations, all of which are very gradual lateral changes, consist of both broad areal trends and more local fluctuations within areas 30 to 40 square miles in extent. For the latter, a vertical difference of 10 feet is about the maximum fluctuation in thickness between any of the layers. The most marked areal trend is a general northwestward thinning of the intervals between all of the lower Trail City assemblage zones. The reduction in thickness of the lower Trail City observed in this trend is about 30 feet, the assemblage zones becoming more closely spaced as the member thins northwestward. The Upper nicolleti Assemblage Zone disappears where the interval between the Limopsis-Gervillia and Protocardia-Oxytoma zones becomes less than 15 feet in thickness. Nowhere in the succession, however, is there any evidence of erosional unconformity or subaerial exposure of the sediments, and the thinning must have resulted from decreased sediment supply or relatively stronger current action.
An apparently unique occurrence of the convergence of two concretion layers is found locally within the Limopsis-Gervillia zone in the area of reduced thickness in the western part of the Trail City outcrop along the Grand River. Here large barrel-shaped concretions, with their long axis perpendicular to bedding, contain masses of Limopsis in the lower part and Gervillia in the upper part. It is evident from the field relationships that the Limopsis masses were either still exposed on the sea floor or had been laid bare by currents when the area became inhabited by the Gervillia association. The local coincidence of the Gervillia masses with Limopsis masses, ultimately to form together a single concretion, poses a problem that cannot be reasonably explained by agencies of sedimentation. It seems more likely that at least the initial localization of Gervillia masses in areas of Limopsis accumulation was controlled by ecology rather than sedimentation-the exposed clusters of Limopsis shells affording the best settling place for the epifaunal Gervillia.
In summary, the stratigraphic relationships of the fossiliferous concretion layers, in combination with their internal characteristics, indicate that individual accumulations were formed simultaneously. The resultant fossilif. erous concretion layers then represent time planes. Relatively level sea bottoms continuously submerged were the site of the successive accumulations.
Distribution Patterns
Three different kinds of distributional features are encountered in examining the geographical aspects of an assemblage zone: the total distribution of the fossil accumulations, distribution of specific associations of animals within the zone, and distribution of specimens in scattered, dense, clusters. In regard to total distribution, only the western limits of the Fox Hills assemblages are known for certain because of the pattern of outcrop. The southern and eastern limits shown for the Trail City assemblages (Fig. 4, 5, 6, 7) are approximations which will probably need some revision if drought ever lays bare the grassed-over slopes. Enough is known, however, to show that the Trail City assemblages have approximately the same geographic limits within the type area and that they define a lobate area trending, and broadening, northeastward and terminating southwestward within the Cheyenne-Moreau divide. The limits of this area of prolific fossil accumulations do not coincide with any pronounced stratigraphic change.
Distribution of the assemblages outside of the type area has not been studied in detail. It is evident, however, that the Lower nicolleti and Limopsis-Gervillia assemblages are the most widespread, and as far as is known, co-extensive. The Upper nicolleti and Protocardia-Oxytoma assemblages do not extend as far to the northeast as the preceding, apparently because of the gradual southward encroachment of more marginal environments. The distribution of the assemblages in the lower part of of the Timber Lake Member shows a pronounced southward shift or spread (Fig. 4). At this stratigraphic level the distribution of the rich assemblages appears to coincide with the clayey marginal areas of the Timber Lake sand body, the remainder of which has in some places a sparse marine fauna similar to that in the rich assemblages, but more commonly has the restricted Tancredia-Ophiomorpha biofacies.
Total distribution patterns of the Fox Hills assemblage zones show that all the areas of maximum fossil accumulation overlap to some extent, and southward shift took place apparently in response to a southward encroachment of more restricted marginal environments. As recorded in the successive zones of fossil accumulations, this environmental shift appears episodic rather than gradual. Instead of the succeeding assemblage zones showing a progressive southward restriction, three groups of zones indicate three positions at which the environment must have remained stable long enough to permit the accumulation of two or more layers of abundant marine mollusc shells and from 15 to 30 feet of sediment. The Lower nicolleti and Limopsis-Gervillia zones mark the initial spread of the Fox Hills fauna in the Missouri Valley region; the Upper nicolleti and Protocardia-Oxytoma zones mark a stage of restriction from the northern part of the original spread; the abyssinus-mandanensis and Idonearca assemblage zones mark a stage of more radical geographic shift southward, and the Mactra-Tellina assemblage represents a much restricted final stage in which the last prolific marine faunas of the type Fox Hills are found.
The second kind of distribution pattern has to do with the occurrence of particular associations of species of fossils within individual assemblage zones. Such associations are present in all of the zones, but they are not commonly conspicuous because of the masking effect of the great numbers of the one or two dominant species. Study of these associations is barely begun but it is evident that they are numerous, that some occur more frequently than others, and that some recur at a number of levels. A few of the more obvious associations such as the large Gervillia and the Drepanochilus masses noted in the Lower nicolleti Assemblage Zone and the Lucina in the Protocardia-Oxytoma zone, appear to have a preferred distribution (Fig. 5, 7).
These examples have in common the fact that a species other than one of the zonal dominants is locally abundant. Less conspicuous associations involving the sparser elements of the accumulations are also present. A good example of this is the nicolleti Inoceramus fibrosus association noted in both the Upper and Lower nicolleti zones. Particularly important to the interpretation of such associations as ecological rather than depositional or chance features is the fact that some 250 feet below the base of the Fox Hills, in the Pierre Shale, an earlier variant of I. fibrosus occurs in association with earlier variants of Scaphites (Hoploscaphites) nicolleti. Whether the nicolleti-fibrosus and other similar associations have recognizable geographic patterns is not yet known. The impression is that they are patchy, with limited areas of distribution.
The prevalence and diversity of the associations within the individual assemblage zones and the preferential geographic distribution and recurrence of some indicate that each assemblage zone is a complexly structured entity. Just how closely these reflect the original ecological distribution of the species involved is not yet known, but their gross resemblance to marine bottom communities is unmistakable even at this preliminary stage of investigation.
The third kind of distribution pattern, clustering, is found in the individual fossil accumulations, the great majority of which are contained in calcareous concretions. That concretion formation most likely took place subsequent to the accumulation and burial of the masses of shells has already been discussed. Distribution of the fossils in scattered clusters that are roughly ovate in shape and show chiefly random orientation is difficult to attribute entirely to sedimentary processes. This is particularly so when one considers not only the preservation of the specimens, which include many complete pelecypods, but also the preservation of patterns of geographic distribution and intricately structured associations. Current action seems to have had very little sorting or redistributing effect on the assemblages; only rarely and very locally do the clusters exhibit layering of fossils or other evidence of bedding that one would expect if the shells had been swept into heaps or into irregularities on the sea floor by currents.
While no completely satisfactory explanation of the clustering of the shells in the assemblages presents itself at this stage of the investigation, the possibility that the masses of shells represent an incipiently clustered distribution of organisms on the sea floor that was accentuated by some current action at the time of burial finds support in the size distribution of specimens. Speden (oral communication) finds that in the bivalve-dominated concretions of the lower Trail City there is marked conformity in the size of specimens of the abundant species in a single concretion. In an adjacent fossiliferous concretion specimens of the same species may be uniformly of a different size. The spotty distribution of sizes does not suggest current action as the size range for specimens of all the species, not just those of the dominant species, is great. Commonly, clusters of tiny bivalves occur in the same concretion with clusters of larger bivalves of similar shape. This intricate and random pattern of size changes among the shell clusters suggests age grouping of the bivalves. Hallam (1961) interpreted clusters of brachiopods from the Jurassic, which have many features in common with the Fox Hills molluscan clusters, as "life assemblages" and suggested the presence of successive annual broods in the clusters on the basis of size-frequency analyses.
On the other hand, the shape of the clusters is difficult to explain as the result of organic agencies alone, and the general lack of either epifaunal or infaunal elements in living position favors at least some influence by sedimentary agencies. Many concretions are as thick in vertical dimension as they are broad and in the principal assemblages most are between 8 and 12 inches thick. The random orientation of specimens, which include both infaunal and epifaunal bivalves as well as elongate elements such as pieces of wood, belemnites, scaphopods and large cephalopods, precludes the growth of the clusters by gradual increment. If, as it appears, the clusters were buried rapidly, they must have accumulated along with about 8 to 12 inches of loose sediment on the sea bottom.
Resemblance to Modern Communities
Two striking characteristics of the successive fossil assemblages are (1) the relatively uniform faunal composition of the individual assemblage zones throughout their extent, and (2) the numerical dominance in each assemblage of one or two species. These same two characteristics are the conspicuous features of present-day marine bottom communities; in fact, as Thorson points out (1957, p. 467), Petersen's concept of marine communities grew out of his repeated observance of these features.
Thorson also notes (1957, p. 468) that one of the more convincing indications of the "reality of Petersen's communities is "... that communities parallel to those described from Danish waters, comprising characteristic animals of the same genera and associated with the same type of substratum, are now known from nearly all parts of the globe." The validity of a natural community thus can be indicated by the recurrence of similar faunally homogeneous associations. Sanders (1960, p. 138), in a recent ecological study of a bottom community, emphasizes this point by using the term community "... to mean a group of species that show a high degree of association by tending to reoccur together." In the study of the Fox Hills assemblages the area involved is too small to test the validity of the individual assemblages as natural communities by the criterion of geographic recurrence. But here the dimension of time, the paleoecologists' one advantage over the ecologist, helps out. Similar fossil assemblages do recur stratigraphically in the interior Cretaceous strata. Within the area of the type Fox Hills this is demonstrated by the Lower and Upper nicolleti assemblages, and particularly by the recurrence of the nicolleti-Inoceramus fibrosus association at these two horizons and in an earlier evolutionary stage in the Pierre Shale some 250 feet lower stratigraphically. Even more convincing evidence comes from preliminary studies of Late Cretaceous (Montanan) assemblages outside the area of the type Fox Hills. The more pertinent recurrent assemblages substantiated to date are the Protocardia-Oxytoma assemblage from older beds in both Montana and Wyoming, and the Limopsis-dominated assemblage from Montana. Even the more restricted assemblage dominated by the gastropod Drepanochilus has been found to recur well down in the Pierre Shale of South Dakota.
In addition to the similarity of the gross features of assemblage zones to those of marine bottom communities, the numerous local faunal associations in the assemblage zones suggest a complex internal structure possibly comparable to the ecological units of varying size and complexity that exist within communities. Any significant comparison of these details must await more complete knowledge of the fossil species, their life habits and their distribution in the assemblages.
In seeking possible explanations for the widespread clustering of fossils within assemblage zones one finds that studies of modern marine bottom populations reveal very little about the finer patterns of animal distribution. This is undoubtedly a consequence of the limitations of the prevalent methods of sampling bottom communities; the dredge does not preserve any evidence of the spatial relations of the recovered animals, and the use of grab and coring devices are generally too spotty to reveal distributional detail. Pratt (1953, p. 62), in a study of the abundance of Venus and Callocardia in relation to type of bottom sediment, makes the following statement which, enhanced by reverse perspective, neatly sums up the factors limiting our knowledge of distribution patterns.
Experience of quahog fisherman has shown that the distribution of individuals in a bed is spotty, the numbers often varying greatly from one square yard to the next; hence a large number of grab samples is required to make an adequate census. In the dredge sample this unevenness is averaged out.A substantial number of scattered references, the majority of which are only brief notations, indicate that clustering is not an uncommon feature in intertidal and sublittoral environments. Observations of clumped distribution of intertidal invertebrates, particularly mussels and oysters, are fairly numerous. Kuenzler (1961, p. 197) in a study of Modiolus demissus in a Georgia salt marsh notes that "mussels are usually found in clumps" and suggests that "since large mussels have very limited motility, the clumps probably result from attraction of small mussels to ones already established and, perhaps, from enhanced growth or lessened mortality in the group." Clustering of more motile molluscs has been less frequently observed. Bradley and Cooke (1959, p. 322-323) attribute the bunched pattern of distribution in Gemma gemma largely to the modification of grouping, in a reproductive pattern, by waves and currents. Moulton (1962) describes clustering in small circular groups of up to a few thousands of individuals in a population of Australian cerithiid gastropods and concludes (p. 176) that the "... clustering is a behavioral adaptation to tropical conditions superimposed perhaps over physiological adaptations..." Loesch (1957), in a study of Donax, observes that living concentrations of this bivalve correspond to beach intercusps. Connell (1955), investigating intertidal distribution of Mya arenaria and Petricola pholadiformis found nothing indicating gregariousness; both random distribution and a tendency to aggregate were evident and he attributed aggregations of the two clams to variations in the substrate.
The examples noted, which are among the few that attempt to explain clustering, serve briefly to suggest the great range and complexity of the modes of clustering, the factors governing it, and the animals involved in it. Beyond indicating that clustering is a fairly common ecological pattern, works on presentday marine communities offer little that is useful in attempting to interpret the fossil clusters of the Fox Hills assemblage zones.
Continuous deposition of sediment, though undoubtedly at varying local rates, is indicated by the nature of the sedimentary record which features uninterrupted and intricate intergrading of facies marginal to and succeeding the beds with the fossil accumulations. No recognizable evidence of marked hiatus or of subaerial exposure yet has been found in the marine lower Fox Hills in the type area, and considering its marginal locus of deposition and position in a gradational series from marine to continental in an area of active sedimentation, it is not likely that significant breaks in deposition occurred. Moreover, both the general uniformity and distinctive distributional patterns of the individual layers of fossil accumulations, as well as the unusually good preservation of the shells themselves, preclude the possibility that the accumulations were formed gradually during long intervals of nondeposition or slight sedimentation. The marked confinement of accumulations to layers separated by unfossiliferous sediments points to abrupt extermination and burial of the existing community and subsequent fresh resettlement of the area, sometimes by the same association and sometimes by different associations of the same basic fauna.
Repeated mass killings, presumably accompanied by rapid burial and relatively little redistribution of the organisms by current action, seems the most satisfactory explanation of the richly fossiliferous layers in the Fox Hills Formation. In her excellent synoptic paper on mass mortality in the sea, which pertains mostly to vertebrates, Brongersma-Sanders (1957, p. 968) states that:
Whether the abundance of invertebrates in certain deposits was caused by catastrophic killing has not been convincingly determined.Ample evidence is given in her paper of both the frequency of mass mortality in the sea and the fact that marine invertebrates are commonly involved in it; consequently, it is reasonable to assume that some accumulations of invertebrate fossils have resulted from such catastrophes, the principal problem is what constitutes convincing evidence.
Abundant vertebrate fossils are so rare in marine sediments that local accumulations are highly suggestive of catastrophe; on the other hand, local abundance of invertebrates is not significant in itself as it could result from a number of causes. But geographically patterned accumulations of abundant invertebrate fossils seem to be convincing evidence of mass mortality, for it is difficult to visualize any alternative way of entombing in marine deposits great numbers of invertebrates that retain both coarse and fine patterns of community structure over sizable areas. The repetition of such patterned accumulations over approximately the same area is strong supplementary indication of mass killing for as Brongersma-Sanders points out (p. 942), certain of the more common phenomena causing mass mortality in the sea are repeated at short intervals.
To what extent the hypothesis of mass mortality can be applied to other interior Cretaceous terrains with similar types of fossil accumulations will have to be determined by detailed study of the individual occurrences in the context of local features of their stratigraphy. Layers of scattered calcareous concretions many of which contain rich associations of fossils dominated by one or two species are a common kind of fossil accumulation in the interior Cretaceous. Two features of these accumulations in particular (1) their apparent preferential occurrence in clayey marine sediments beneath and laterally adjacent to marginal facies of restricted marine or mixed environments, and (2) the repetition in successive layers of particular associations of dominant genera, are, I believe, strong reason to suspect that mass killings of the bottom fauna may have been common occurrences repeated many times in certain environments of the coastal waters of the interior Cretaceous sea.
Possible Causes
A thorough review of the known causes of mass mortality and step by step comparison with the fossil accumulations in question is not warranted here. The range of possibilities is well documented in Brongersma-Sanders paper (1957), which is necessary background for any discussion of the subject. Unfortunately, we do not know enough about the oceanography of epicontinental seas to be able to rely on analogy with mass mortality in modern seas with complete confidence. This is especially true for the interior Cretaceous sea whose faunal peculiarities are well known. Somehow it seems presumptive to speculate about how certain communities of the fauna died when we know so little about how they lived. However, consideration of pertinent features of the assemblages in question may at least provide some idea of the nature of the catastrophes involved.
The repetition of the assemblage zones with similar features over similar areas indicates that the mass killings were recurrent and suggests that the agent was the same for each successive killing. Three causes of modern mass mortality, low temperature, hypersalinity, and noxious waterbloom, recur frequently in local areas (Brongersma-Sanders, 1957, p. 942). For the Cretaceous interior sea, particularly for the shallower restricted phases of it marginal to deltas, reduced salinity might be added to the list. The possibility of recurrent low salinity in coastal areas seems a legitimate assumption considering the great size of the area draining into the sea and the humid warm temperate to subtropical climate indicated by the Late Cretaceous flora (Dorf, 1942, p. 100-103). This does not eliminate the possibility of local or even widespread hypersalinity in the interior sea, although there is no obvious indication of it, and to my knowledge it has been proposed only on theoretical grounds.
The faunal constitution of the Fox Hills assemblages suggests selective killing. Each layer contains the same principal types of animals; infaunal and epifaunal molluscs and ammonoids--chiefly scaphitids. The latter were most likely slow swimmers feeding on or near the bottom. Fish remains, except for scattered scales, are rare; belemnoids are present but not abundant and never found in concentrations. It would appear that the killing, whatever the cause, did not take place rapidly enough to catch the more mobile organisms such as fish and belemnoids. The absence of these more capable swimmers also suggests that the area in which the mortalities took place, though an embayment, was open to the sea.
The floral constitution of the assemblages may be significant. Of the varied vegetable contents of the assemblage zones, wood fragments, commonly infested by the wood-boring bivalve Martesia, are the more abundant. Fragmental to complete leaves, conifer cones and foliage, fragments of palm fronds, nutlike bodies, charcoal fragments and slender trunks of Palmoxylon up to 8 feet long, have all been found in the assemblage zones. While not in abundance, these plant structures are sufficiently common to attest to the proximity of land and of rivers competent to sweep such debris out to sea in times of flood.
Clustering and concretion formation are both features that have a bearing on the killings. As has been noted, the clusters may have resulted from the accentuation of an original ecological clustering by some sort of mild current action, and it was indicated that from 8 to 12 inches of sediment are involved in the deposition of individual layers. If, as seems likely, concretion formation resulted from the decay of buried organic matter, both clustering and concretions call for either the rapid deposition of up to a foot of sediment or the roiling of the bottom sediment to that depth. Strong sweeping currents are ruled out by lack of sorting, by the cluster pattern itself, and by the random orientation of shells.
The thin bentonites in the Fox Hills bear no direct relation to the fossil assemblages. Although there was appreciable volcanic activity in the Late Cretaceous of the interior region, there is no evidence that the marine faunas suffered directly from it in any way. In fact, the disappearance of the ash in the areas of abundant invertebrates suggests that the sediment feeders literally ate it up.
Bringing together the scanty direct evidence we can say that the fossil assemblages were associated with a recurrent phenomenon, localized in a fairly large but geographically restricted coastal area, which selectively affected the bottom fauna and slow-swimming invertebrates and accomplished the deposition and redeposition of approximately a foot of bottom sediment without the aid of currents strong enough to redistribute and sort the organisms. This suggests that turbidity was an important factor at the time the accumulations were buried, but if the beds in which the accumulations are found were ever graded, the evidence was subsequently destroyed by burrowing organisms. Some small-scale graded bedding is at least locally associated with the thin-bedded silt and silty shale facies into which the fossiliferous sequence grades to the west; its frequency of occurrence is not known.
Recurrent phenomena that could conceivably have caused turbid bottom conditions include severe storms, which could both disturb bottom sediment and bring about the introduction of sediment-laden currents from rivers in flood; flooding of rivers in periods of excessive rainfall not associated with severe storms; and stirring of the bottom by tsunamis. Tsunamis cannot be ruled out as a frequently recurrent phenomenon considering that quakes may have been common during the Laramide mountain building then in progress nearby in the interior region, but the effects of such disturbance should be fairly widespread and not restricted to the limited area of fossil accumulation. If evidence for disturbances of the bottom exists lateral to the horizons of fossil accumulations, it is not apparent. It is also questionable whether burial in bottom sediment alone could have destroyed the dominant faunal elements found in the associations. Certainly the ammonoids and epifaunal bivalves could have been killed in this manner, but we do not yet know enough about the life habits of the infaunal bivalves involved to determine whether they are types that would not have been able to re-orient themselves in the sediment and continue to live. If the study of the bivalves in progress indicates that the dominant infaunal species present are exclusively types that could not have readjusted to burial, the selectivity of the mortality will add considerable support to the possibility of smothering.
If turbidity resulted from the discharge of sediment-laden rivers during storms or periods of torrential rainfall, reduction of salinity may have acted together with smothering to cause mortality. In this situation even bivalves that could adjust to burial would have suffered. The chief obstacle to this possibility is that reduction of salinity by freshets usually affects only very shallow bottoms, generally not below 3 meters, as the fresh water being less dense than the salt, tends to remain on the surface. Conceivably, highly turbid fresh water because of its greater density, could affect bottoms to greater depths, but I know of no recent studies offering quantitative data on this subject. The salinity of the interior sea, particularly in coastal areas, is also an important consideration here, and the possibility that it was considerably less than the salinity of normal oceanic waters has yet to be either substantiated or disproven. The scarcity of such common invertebrates as echinoderms, corals, and bryozoa in the interior Cretaceous sea could result from low salinity, excessive turbidity, or even hypersalinity. Whatever the salinity of the sea, the inclusion of plant remains in the marine assemblages points to the recurrent local influx of highly turbid fresh water from rivers in flood.
None of the foregoing speculations are supported by sufficient evidence to rule out other possible causes of mortality. Periodic low temperatures could have occurred, as they do today in the Gulf of Mexico, in the warm temperate to subtropical climate of the Cretaceous. Noxious waterblooms may also have been common. However, the absence of fish in the assemblages, unless explainable in some other way, does not favor either of these common kinds of recurrent mortality. The actual cause or causes of mortality may not be determinable from what evidence is left in the record. Even in some present day mortalities the multiplicity of factors involved obscures the principal causes of killings. In hurricanes, for example, killing may locally result from stranding, hypersalinity in brackish waters, lowered salinity in coastal waters, burial and deoxygenation, as well as other direct physical effects. Possibly the prevailing bottom conditions in much of the Cretaceous sea were inhospitable to life and became the agent of mortality following brief episodes of hospitable conditions brought about by some recurrent environmental factor. If so it is the existence of the benthic faunas rather than their extermination that requires special explanation. But until more is known of the life habits of the species in the peculiarly restricted interior Cretaceous faunas, we are not likely to understand the factors involved in either their great local abundance or their repeated local extermination.
Bradly, W. H., and Cooke, Peter, 1959, Living and ancient populations of the clam Gemma gemma in a Maine coast tidal flat: U. S. Fish and Wildlife Serv., Fish Bull., v. 58, no. 137, p. 304-334.
Brongersma-Sanders, Margaretha, 1957, Mass mortality in the sea; in Treatise on marine ecology and paleoecology: Geol. Soc. America Mem. 67, v. 1, p. 941-1010.
Burt, F. A., 1932, Formative processes in concretions formed about fossils as nuclei: Jour. Sed. Pet., v. 2, no. 1, p. 38-45.
Connell, J. H., 1955, Spatial distribution of two species of clams Mya arenaria L. and Petricola pholadiformis Lamarck, in an intertidal area: Mass. Shellfisheries Inv., Rept. no. 8, p. 15-25.
Dorf, Erling, 1942, Upper Cretaceous floras of the Rocky Mountain region; 2. Flora of the Lance formation at its type locality, Niobrara County, Wyoming: Carnegie Inst. Washington Pub. 508, p. 83-168.
Hallam, A., 1961, Brachiopod life assemblages from the Marlstone rock-bed of Leicestershire: Palaeontology, v. 4, pt. 4, p. 653-659.
Kuenzler, E. J., 1961, Structure and energy flow of a mussel population in a Georgia salt marsh: Limnol. Oceanogr., v. 6, no. 2, p. 191-204.
Loesch, H. C., 1957, Studies of the ecology of two species of Donax on Mustang Island, Texas: Texas Inst. Marine Science, v. 4, no. 2, p. 201-227.
Moulton, J. A., 1962, Intertidal clustering of an Australian gastropod: Biological Bull, v. 123, no. 1, p. 170-178.
Pratt, D. M., 1953, Abundance and growth of Venus mercenaria and Gallacardia morrhuana in relation to the character of bottom sediments: Jour. Marine Research, v. 12, no. 1, p. 60-74.
Sanders, H. L., 1960, Benthic studies in Buzzards Bay. III; The structure of the soft-bottomed community: Limnol. Oceanogr., v. 5, no. 2, p. 138.153.
Thorson, Gunnar, 1957, Bottom communities (sublittoral or shallow shelf), in Treatise on marine ecology and paleoecology: Geol. Soc. America Mem. 67, v. 1, p. 461.534.
Todd, J. E., 1910, Preliminary report on the geology of the northwest-central portion of South Dakota, in Report of the State Geologist for 1908: South Dakota Geol. Survey Bull. 4, p. 13-76, 193-207.
Waage, K. M., 1961, The Fox Hills Formation in its type area, central South Dakota, in Symposium on Late Cretaceous Rocks; Wyoming Geol. Assoc. 16th Ann. Field Conf. Guidebook, p. 229-240.
Weeks, L. G., 1957, Origin of carbonate concretions in shales, Magdalena Valley, Columbia: Geol. Soc. America Bull., v. 68, p. 95-102.