
|
Original published in D.F. Merriam, ed., 1964, Symposium on cyclic sedimentation: Kansas Geological Survey, Bulletin 169, pp. 481-495 | |
Wichita State University, Wichita, Kansas
The formation is generally viewed as a unit which was deposited in epeiric seas. However, as the evaporitic sequence (Hutchinson Salt, Annelly Gypsum) indicates, for a period of time Wellington waters were hyper-saline. Furthermore, as this paper will show, from base to top of the formation, there is considerable evidence to support an interpretation of an alternation of conditions from limited evaporative to brackish to fresh.
The general picture that has emerged from the research to be discussed is one of a coastal flat with relict puddles, ponds, and even small-scale lakes that gradually freshened as the epeiric sea regressed. Marine encroachments over this coastal area occurred between times of recurrent fresh-water biofacies.
Thickness of the Wellington Formation is about 700 feet. The base of the formation is the contact with the Herington Limestone and can be seen exposed in Cowley County, Kansas (SE NE sec. 21, T. 34 S., R. 3 E.). One can proceed in an almost straight line to the top of the formation, a distance of 23 miles, west of the Cowley County exposure to where the upper member of the Wellington, the Milan Limestone, is exposed in Sumner County (NW NW sec. 23, T. 34 S., R. 2 W.). Along and near this traverse line, as well as south (Kay and Noble Counties, Oklahoma) and north of it (Sedgwick, Harvey, Marion, and Dickinson Counties, Kansas) are numerous but discontinuous exposures.
Field exploration for the past four years and subsequent laboratory study of samples have brought to light new information on the Wellington Formation. Because material is still being studied, however, field notes will be used for sections still to be processed.
Acknowledgments--Field and laboratory research on the Wellington Formation has been supported by National Science Foundation grants (G-4150, G-7320, G-14141). The Kansas Geological Survey prepared the illustrations for this paper.
Thus, there is clear evidence of a post-Herington regressing, epeiric sea. The blocky, waxy shales represent shallow-marine deposits; the Lingula bed, a nearshore deposit. With further regression of the sea, a coastal flat bearing relict puddles and ponds remained. The snail-ostracode coquina seemingly accumulated in such relict brackish-water bodies. Thereafter, at this locality (see, Table 1), minor oscillations of the sea-cover on and off the coastal flat can account for the snbsequent shales and fresh-water biota found in them.
Table 1--Basal Wellington in Cowley County, Kansas. Fresh and brackish-water biofacies (SE NE sec. 21, T. 34 S., R. 3 E.).
| Height above top of Herrington, feet |
Lithology | Fauna or faunal element (Macroscopic)1 |
|---|---|---|
| 138 | Gray, waxy shale |
Reptilian fragments;2 snail-ostracode coquina |
| 247 | Buff-gray argillite |
Reptilian fragments (scales, etc.) |
| 306 | Green, waxy shale |
Conchostracans; reptilian fragments |
| 387 | Green, platy shale |
Reptilian fragments; carbonaceous plant fragments |
| 403 | Massive, reticulated argillaceous limestone |
Reptilian fragments; worm burrow? |
1. Microscopic floral elements are discussed in the section on "Carbonized wood/plant beds."
2. Objects referred to as reptillian fragments throughout this paper, properly belong under fishes. This correction is important because fish are generally absent in modern conchostracan-bearing ponds in Kansas and Oklahoma, a condition that markedly contrasts with their frequent association with fossil conchostracans in the Wellington Formation (Permian).
A snail-ostracode coquina, including reptilian fragments, also occurs in Dickinson County, Kansas (Elmo VIII-B; Tasch, 1963a), at the approximate horizon of the "Carlton Insect Beds." Preliminary study shows that the two coquinas appear to have similar species.
Two things are striking about this recurrence of a biofacies: first, the Cowley County occurrence (Table 1) is 12.3 feet above the top of the Herington, whereas the Dickinson County coquina is approximately 250 to 300 feet above that top; second, Myalina meeki Dunbar, a brackish-water pelecypod, occurs in the Dickinson County coquina (Tasch, 1961, 1963a).
Thus, the interpretation derived from study of the transition from the marine Herington to the snail-ostracode coquina in Cowley County is sustained by the appearance of a brackish-water mollusk in the almost identical type of coquina in Dickinson County. It is such evidence taken from different localities in contemporaneous beds and in beds at different elevations above the datum that have led to the environmental interpretations given in this paper.
At the Cowley County section we also are afforded the earliest glimpse of the primary periodicity in the Wellington, i.e. recurrence of fresh-water, brackish-water deposits containing conchostracans, fish remains, and other fossils (Table 1).
Whereas the snail-ostracode coquinas are strictly limited in recurrence, one or another of the biofacies indicated in Table 1 recur intermittently to the top of the Wellington (Milan Member). We thus have unmistakable evidence of periodic fresh- to brackish-water conditions throughout Wellington time.
Evaporites (hopper crystals, salt casts, and gypsum), generally in small quantities, are found in recurring conchostracan-bearing beds. These evaporites provide a further criterion that these creatures lived in freshening relict ponds and puddles on a coastal flat. The oldest hopper crystal-bearing bed (Wellington XVIII) occurs an estimated 50 to 70 feet above the top of the Herington. (Note: A tabulation of all localities is appended at the end of this paper.) The youngest occurrence of evaporties was found two-tenths of a foot below the upper Milan Limestone which caps the Wellington Formation (Wellington XI).
A large-scale periodicity in the Wellington is the recently discovered cyclical recurrence of insect beds at approximately lOO-foot intervals above the top of the Herington. Exclusive of the fourth 100-foot interval (for which data are lacking), insect beds were found in the first, second, third, fifth, and sixth 100-foot intervals above the top of the Herington (Tasch, 1962, Table 1). Of these insect beds, five contained conchostracans as well as insects, and other fresh-water biota.
The conchostracan-bearing beds represent the most persistent periodicity in the Wellington. These are found intermittently through out the formation starting with the basal 40 feet and extending to the Milan Member. Two conchostracan-bearing beds also were found in the interbeds of the Annelly Gypsum, the top of which is an estimated 210 to 260 feet above the Herington.
A brief consideration of the conchostracan-bearing beds is necessary at this point to clarify the way in which a microscale periodicity was deciphered. It has been shown that in Leonardian time conchostracans intermittently occupied the same pond sites (themselves intermittent) in Sedgwick, Harvey, and other counties in Kansas. Such on and off occupancy ranged from 40 to 150 years, and frequently conchostracan occupancy lasted for a single season only. This determination was made for conchostracan-bearing beds 0.12 to 0.45 feet thick (Tasch, 1961).
As an example, we can examine one slab of an argillaceous limestone 0.32 feet thick. Eighteen distinct conchostracan generations were found in this slab. These creatures were and are seasonal animals that still persist in ponds in the same general area as the Wellington outcrop belt (Tasch and Zimmerman, 1961a). The time span calculated as necessary for this thickness to have been deposited was about 106.6 years. Because the slab is lithologically uniform, no periodicity could have been detected if one viewed the entire thickness as the result of a single unchanging depositional event.
However, separating this slab into distinct conchostracan-bearing surfaces, along which it readily parted, revealed a periodicity. The surfaces bearing clam-shrimp fossils represented times when pond conditions permitted clam-shrimp eggs to hatch and pass from naupliid to adult stage in a normal growth cycle covering a few months. By contrast, between these fossiliferous surfaces, the rock was barren of fresh-water biotic elements indicating that pond conditions represented could not sustain clam-shrimp populations. If the barren intervals were thicker, they would represent destruction of a given pond site by a marine influx and subsequent restoration of the pond site at a later time.
Many variations of this type of periodicity were found. Successive events in thin slabs of Wellington shale or limestone might be marked by other fresh- to brackish-water biotic elements such as mollusks, fish fragments, carbonized wood and plants, xiphosura, insects, and occasionally other items in the complete absence of conchostracans. In turn, these permitted the reconstruction of microcycles at a given pond site.
Thus, within the larger context of recurring fresh- to brackish-water conditions during Wellington time, it is necessary to abstract microscale periodicities in thin lithologic units. These permit interpretations of paleolimnological events on a basis heretofore unthinkable for a Paleozoic deposit, that is, a season-by-season account of successive events.
Thus, three major types of periodicities differing in scale are indicated:
Large scale--Insect beds recurring at approximately lOO-foot intervals.
Small scale--Recurrent conchostracan beds, and multiple insect beds in 5 to 10-foot intervals, and the usual biotic associates of clam shrimps in beds lacking clam-shrimp fossils.M
Microscale--Seasonal events separated by sediment intervals of 0.02 to 0.04 inch within insect and conchostracan-bearing beds.
All three of these periodicities are part of what is here designated as the "paleolimnological cycle," which will be discussed more fully in another section.
Events during Wellington time also include some acyclical deposits, i. e., those that are restricted in the formation. Among these are: the Lingula bed (discussed earlier), marine algal reefs, Hutchinson Salt Member and the Annelly Gypsum, a hystrichsphaerid-dinoflagellate assemblage at a single locality and horizon (Tasch, 1963b), and several alga belonging to the same species at a single locality and single horizon (Wellington II-C, bed3). (Note: The well log of the salt at a mine in Hutchinson shows cyclical deposition of salt, anhydrite, and shale. However, relative to the entire Wellington, the evaporite event may be viewed as acyclical. The same holds for the Annelly Gypsum.)
Table 2--Composite of all clam shrimp-bearing beds and other biotic horizons in a 4-county area (Sedgwick, Harvey, Marion, and Dickinson Connties, Kansas).
| Clam shrimp C-horizons |
Other faunal horizons |
|
|---|---|---|
| C17-56.01 | ||
| 10' cover | ||
| C16-47.5 | ||
| C15-47.4 | ||
| C14-45.4 | ||
| C13-45.1 | ||
| C12-45.0 | ||
| 44.8' log bed | 42'-44' snail-ostracode coquina |
|
| C11-43.1 | ||
| C10-42.1 | ||
| 42.0'insects2 | ||
| C9-41.5 | ||
| 41.0' insects | ||
| C8-40.4 | ||
| C7-39.9 | ||
| 37.2' insects | ||
| C6-28.2 | ||
| C5-27.7 | ||
| C4-26.O | ||
| 22.4'-24.2' plants3 | ||
| C3-21-22 | ||
| 16.5' mollusks | ||
| C2-11.5 | ||
| C1-1O-11 | insects | |
| C0-3.5BG | ||
| C00-5.78BG | ||
1. All evaluations are in feet above the Annelly Gypsum except C0 and C00 which are given in feet below the top (BG). Total thickness: about 65 feet.
2. The three insect beds constitute the "Carlton Insect Bed" and taken together are one of the large-scale periodicities discussed in this paper.
3. Plants and mollusks are also found in many clam-shrimp beds in addition to the indicated horizons. This also applies to other biotic elements.
Table 3--Fresh-water biofacies in Wellington Formation of Noble County, Oklahoma (Noble I A-D).1
| Elevation above base of section, feet |
Clam-shrimp fossils |
Insects | Plants and carbonaceous beds |
Fish fragments and xiphosurans |
|---|---|---|---|---|
| 9.2 | X | |||
| 21.32 | X | X | ||
| 21.4 | X | X | X | X |
| 22.0 | X | |||
| 22.73 | X | X | ||
| 22.8 | X | |||
| 24.14 | X | X | ||
| 31.45 | X | X | ||
| 31.5 | X | |||
| 31.9 | X | |||
| 32.4 | X | |||
| 33.8 | X | |||
| 34.9 | X | |||
| 37.7 | X | |||
| 44.6 | X | |||
| 45.9 | X | |||
| 46.0 | X | X | X | |
| 47.0 | X |
1. total thickness of composite section is about 77 feet.
2, 3, 4. Respectively, the lower, middle, and upper insect beds of the Midco.
5. The Asthenohymen-Delopterum Insect Bed (Tasch and Zimmerman, 1962).
Table 4--Fresh-water biofacies in Wellington Formation of Kay County, Oklahoma (Wellington XIX).1
| Elevation above base of section, feet |
Clam-shrimp fossils |
Insects | Plants | Xiphosurans? |
|---|---|---|---|---|
| 19.9 | X | |||
| 21.6 | X | |||
| 21.7 | X | |||
| 28.1 | X | |||
| 28.6 | X | |||
| 30.0 | X | |||
| 30.5 | X | X | ||
| 31.12 | X | X | X |
1. Total thickness of section is 41.3 feet.
2. This bed has been tentatively designated as the "Youngest Insect Bed" (YIB; Tasch, 1962).
The sediment interval between any two successive fossiliferous beds varies between very short intervals of tenths of a foot to short intervals of 3 feet or less, and finally to intervals of greater than 5 feet. Study of these tables indicates that in the Kansas section (Table 2), 83 percent of all sediment intervals are 3 feet or less; in the Noble County section (Table 3), this interval occurs 75 percent of the time, whereas in the Kay County section (Table 4), it occurs 86 percent of the time. Since these three sections represent different portions of Wellington time, it is apparent that the fresh-water, brackish-water biofacies followed a similar rhythm in recurrence.
The basal Wellington, by contrast (Table 1), had only thicker sediment intervals of 5 feet or greater between successive biofacies. At this locality it appears that the epeiric sea covered the coastal area for longer periods of time. Only when this sea retreated from the coastal area were relict brackish- to fresh-water bodies available.
Very short sediment intervals between successive biofacies separated by a few hundredths of an inch are unlikely to be related to marine transgression. Rather, these represent persistent pond conditions through several seasons or years. Approximately 50 percent of all sediment intervals are less than 1 foot in thickness. In most of these intervals decades of time separated any two successive conchostracan-bearing beds.
Figure 1A--Small-scale algal (biscuit) structure.
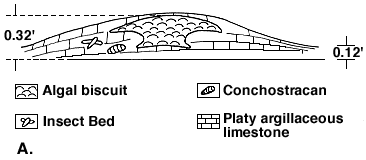
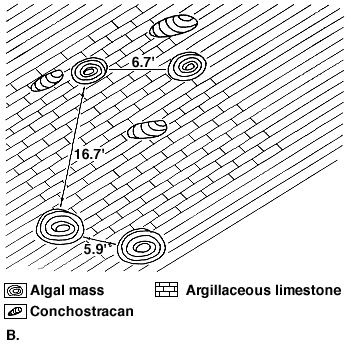
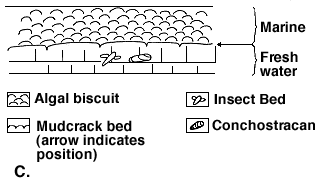
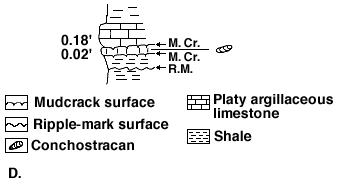
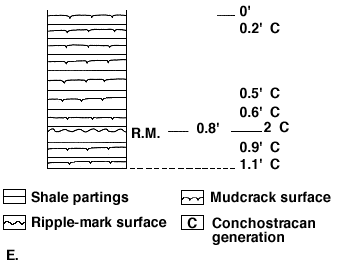
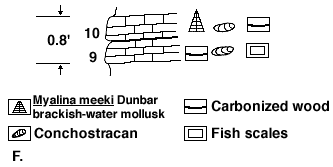
Figure 1A shows a small-scale algal (biscuit) structure. This was secreted by marine algae when the epeiric sea washed over a portion of the coastal area dotted with ponds and puddles in which conchotracans thrived. Insects are found on the flanks of these algal structures suggesting freshening pools as the sea receded.
Figure 1B shows such algal masses and their areal disposition. These structures created a shallow-water topography. Pools that formed between and around such structures freshened and bore conchostracans.
Figure 1C indicates that after the insect-conchostracan bed was deposited, the bottom muds became desiccated (as shown by the mudcrack bed). That, in turn, was followed by a marine transgression as evidenced by the algal bed.
A few tenths of a foot of section is shown in Figure 1D that represents a portion of the Milan Member of the Wellington. The ripplemark surface, which denotes very shallow water, was succeeded by a mudcrack surface that denotes a regression of the sea. A clam shrimp-bearing, argillaceous limestone shows that relict ponds had freshened only to be followed by a period of desiccation (upper mudcrack surface). The sea once again covered the area and deposited a barren limestone.
Ten distinct sets of mudcrack surfaces are shown in Figure 1E; these occur within a thickness of 1.1 foot. There is also a ripple mark surface in the lower third of this sequence. In and between several mudcrack surfaces, there are four conchostracan generations represented. Two additional conchostracan generations occur on and directly below the ripple-mark surface. The mudcrack surfaces are separated from each other by very thin shale partings. Such surfaces and the recurrent fresh-water fauna clearly represent alternate wetting and drying conditions in tbe area. A tongue of the sea could wash over the coastal area briefly then retreat during which time the mudflat dried up except for isolated puddles, then the sea could wash over the area again, etc.
The association of conchostracans on and below the ripple-mark surface suggests that they thrived in nearshore puddles, probably brackish, and when current ripples developed in the shallows and the sea retreated again, they thrived in narrow lenses of brackish water in the troughs between ripple crests.
Figure 1F illustrates the type of fresh- to brackish-water, clayey-lime mud deposition. Carbonized plants and conchostracans occur throughout the 0.8 foot of limestone. The upper of the two units contained the brackish-water pelecypod Myalina meeki Dunbar. These limestones clearly were deposited in a brackish-water swamp environment which was marginal to a coast. The presence of conchostracans indicates freshened isolated puddles on this clay-carbonate mud bottom.
Interpretation of the depositional environments throughout Wellington time is derived from evidence of the type discussed above. Multiple examples of the types mentioned and a wide range of other examples lend support to the conclusion that, exclusive of the major evaporitic deposition in the Wellington, the absence of the fresh- to brackish-water biofacies in any portion of the Wellington denotes shallow-marine deposition.
Carbonized wood, etc., occurs 38.7 feet (Table 1) above the Herington in Cowley County (Cowley I). A series of slides of all beds of this section processed for spores, pollen, and carbonized debris adds information to this picture. Carbonized material also occurs 6.1 and 36.1 feet above the Herington. At Wellington XVIII carbonized material was found in the insect bed which is an estimated 50 to 70 feet above the top of the Herington Limestone.
Carbonized plant facies occur in the Noble County sections ("Midco Insect Beds" are approximately 550 feet above the Herington) as follows: 538, 551, 567.9 and 575.8 feet; these may be broadly interpreted as filling the 540-, 550-, 560-, and 570-foot intervals above the datum.
In the estimated 595 to 668-foot interval above the Herington there is negligible macroscopic carbonized wood/plant material (Wellington XIX; elevation of the youngest insect bed in the Wellington at this locality is an estimated 630 to 668 feet; see, Tasch, 1962, Table 1). Palynological analysis however, shows considerable carbonized wood, spores, and pollen in this interval. These occur at 6 to 9-foot intervals with several tenths of a foot recurrences between some of the larger intervals.
Another bit of evidence may be cited here. Elsewhere (Tasch, 1960), an occurrence of carbonized wood in shale interbedded with Hutchinson Salt in the Carey salt mine at Hutchinson was reported. Since then, the writer has found carbonized floral debris (gymnosperm tracheid cell, not unknown in other Wellington beds) in shale in-fills in this mine. The writer's former field assistant, Bernard Shaffer, demonstrated that a microfloral suite of spores and pollen occurred in the salt both at the Hutchinson and Lyons, Kansas, mines (Shaffer,1961).
If, as Dunbar suggested (1924, p. 194), the upper portions of the Hutchinson Salt lie at or slightly below the level of the "Carlton Insect Beds," then the carbonized wood and microfloral suite found in the salt should correspond to one or another of the recurrent carbonaceous facies in the 220 to 290-foot interval above the Herington.
The picture that emerges at present indicates that small-scale rhythmicity of carbonaceous facies in the Wellington can be demonstrated on the macroscopic level for the following interval, above the Herington: 220 to 290, and 540 to 570 feet. On the microscopic level, comparable rhythmicity can be shown for the 595 to 668-foot interval. The record of the basal one hundred feet as noted, is sparse. This is true even when implemented by palynological study of all exposed beds in the sequence. However, the same type of recurrence of carbonized wood facies is apparent although the sediment interval between any two successive carbonized wood facies in this portion of the column is generally greater than in other sections. With these restrictions in mind, one can also include the 6 to 70-foot interval.
This record might be less spotty if more and better exposures were available for sampling. Nevertheless, it does establish the recurrence of both macroscopic or microscopic carbonaceous facies in the Wellington.
A distinct floral absence is indicated for the upper 32 feet of the Wellington (above 668 feet), that persisted through all of the overlying Ninnescah Shale. (Note: Palynological study of the Milan type section and all Ninnescah beds failed to reveal any palynomorphs of any kind. Several Ninnescah samples were rerun as a check also with negative results.)
It has already been suggested (Tasch, 1962) that this floral absence could "reflect some important environmental change." However, whatever change did occur, affected only the insects which are also absent above 668 feet and all through the Ninnescah Shale. (Note: The striking biotic change that occurs above the 668-foot level, suggests that this level should logically be the top of the Wellington.)
The Wellington paleolimnological cycle has two phases: Phase I--the coming into existence of temporary ponds. Two different periods are associated with this phase, namely, the large-scale period, recurrent, insect beds, and small-scale period, recurrent, conchostracan beeds. Phase II--seasonal events in ponds that come into being (recur) in Phase I. Here, the only period is microscale, i.e., event, separated by a few millimeters of sediment.
Phase I generally involves a transition from normal marine to brackish- to fresh-water conditions. The step from normal marine to brackish is first achieved by regression of the sea. The step from brackish to fresh is achieved by evaporation that acts to fractionate the brackish zone into isolated ponds and puddles.
Presence of large-size hopper crystals and considerable numbers of salt casts and gypsum in a fresh-water biofacies can be explained by omitting the brackish-water step. The complete absence of evaporites in any quantity suggests that the brackish-water step wa, a prelude to the fresh-water biofacies.
Thus the two types of events would be:
A: Normal marine --> regression --> fractionation into relict pond. --> evaporation --> fresh-water ponds. . .
B: Normal marine --> regression --> brackish inner margin --> evaporation --> fresh-water ponds. . .
Where the fresh-water biofacies terminates and is succeeded by nonfossiliferous shales, a slight normal marine transgression over the area is indicated (Fig. 1C).
There are, of course, variants of A and B. During Annelly time, conditions were obviously evaporative, and a cutoff margin of an inland sea gave rise to the gyp,iferous depo,it. Because conchostracans occur in some shale interbeds of the gypsum, it is apparent that the same evaporatic conditions that caused shrinkage of the Annelly basin also created temporary fresh-water conditions on its margins (with a one-to-two inch cover of water) . That would define a modified cycle.
C: Normal marine --> cutoff --> evaporation leading to evaporitic deposits --> marginal fresh-water ponds --> marine influx --> cutoff --> evaporation leading to evaporitic deposits. . .
There is another variant of C that does not generally involve evaporites (Fig. 1E).
D: Evaporation or shrinkage of brackish-water zone --> relict puddles on mudcrack surface. bearing fresh-water biota --> brackish-water cover --> evaporalion or shrinkage, etc.
Figure 2 illustrates the biotic spectra of the insect beds, which, as noted previously, constitute the large-scale periodicity in the Wellington. The recurrent features of these beds are: (1) same faunas (frequently on the species level, although particular insect genera show changes in ratios of abundance); (2) evaporites (salt casts and hopper crystal,); and (3) carbonaceous material (wood and plant debris).
Figure 2--Biotic spectra of insect beds. A larger version of this figure is available.
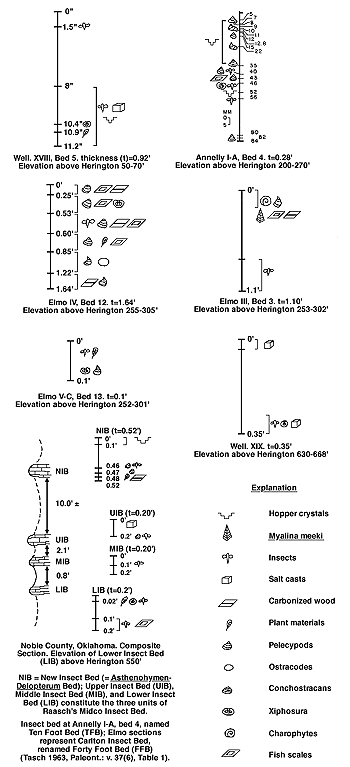
Ten feet above the Asthenohymen-Delopterum insect bed in Noble County, Oklahoma (Noble II B, bed 3), 25 conchostracan generations were found in a thin, fissile shale 3.2 inches thick. These generations occurred below the top of the bed (in millimeters) as follows: 3.5; 5.0; 10.0; 13.0; 15.0; 21.0; 23.0; 24.0; 25.0; 27.5; 32.5; 33.0; 34.5; 36.5; 38.5; 39.5; 41.0; 46.0; 47.5; 49.5; 51.5; 53.5; 56.5; 58.5; and 62.5 (this is bottom surface).
The smallest sedimentary interval between two conchostracan-bearing surfaces was 0.5 mm (= 0.0192 inch = 0.02 inch; Tasch and Zimmerman, 1961b; Tasch, 1961). The time represented by this one fossiliferous bed (3.2"/0.02") is about 160 years. If there had been one generation per year, there should have been 160 conchostracan generations. Actually, about one-fifth of maximum was found to have occurred.
Table 5 graphically illustrates the paleolimnological cycle represented by the Noble County sample. The graph shows the variable time value in seasons for successive sediment intervals between clam shrimp-bearing surfaces. The table is to be read as follows: Generation I died out and the pond remained barren for eight successive seasons (years). Generation II lived a season and for the next four seasons the pond site was barren, etc.
Table 5--The Paleolimnological Cycle, Phase II--Leonardian Wellington of Noble County, Oklahoma (Noble II-B, bed 3, thickness 3.2 inches). Each conchostracan-bearing surface in this rock represents one season (year) with the exception of Generation XIV-XV, which are interpreted to be "same-season" generations. For the total history of the pond-site these seasons must be added to those represented by the sediment intervals between conchostracan-bearing beds.

A similar, yet more complete, cycle, because a more complex biota occurred in the life of the pond site, was found in Harvey County (Annelly IA; see, Tasch, 1961, Fig. 2). This cycle has yet to be evaluated on a seasonal basis but that will be done here (Table 6).
Table 6--Paleolimnological cycle--Phase II, Harvey County, Kansas (Annelly I-A).1

Table 6 indicates the following history during the time represented by 0.28 foot of sediment deposition.
Generation VII then appeared and within the same season, Generation VIII. This is evidenced by the sediment interval which is less than the annual rate of sedimentation in the area. Thereafter, for two successive seasons, while clam shrimps were totally absent, the pond site supported Productae dunbaris.
After two more seasons, Generation IX appeared, followed in the next season by Generation X, and two seasons later by Generation XI.
Thus, the intermittent occupancy of the given pond site by fresh-water biotas covers almost a century of time and is now represented in an argillaceous limestone a few tenths of a foot thick.
Fresh-Water Deposits
The successive clam shrimp-bearing beds are generally units of a few tenths of a foot thick which testifies to the relatively short duration of the water bodies they represent. Blocky, platy, laminated waxy shales and argillaceous limestones are the recurrent lithologies in such beds. The color of the thin limestones is generally gray-blue weathering gray-white. These limestones are hard beds and stand out in outcrop since they are more resistant to weathering than overlying or underlying shales. Equivalent beds in Kansas sections have greater than 50 percent carbonate as determined by insoluble residue studies.
The shales are generally some variation of green, black-green, or gray-green. Red shale is rare in the Kay County section.
From study of the lithologies of the 40 plus clam shrimp-bearing beds of Kay County, certain observations emerge:
(1) Small quantities of carbonate (or the complete absence of carbonates) in the bottom muds at any given time had no effect whatsoever on conchostracan growth and reproduction. These forms are as abundant and sometimes more abundant in the shales as in the limestones. Some of the shales, of course, are calcareous.
(2) Despite its thinness, the argillaceous limestone within a given bed frequently shows gradation upward or downward to blocky, platy or laminated shale. That would indicate temporary and limited increase in carbonate in the given water body where the dominant sediment was clay mud.
(3) The top beds of the Kay section are siltstone and sandstone. These are laminated and cross-bedded as well as pelletiferous. Several surfaces contain conchostracan fossils.
Somewhat lower in the section (Fig. 1E), a mudcrack bed composed of argillaceous siltstone showed ripple marks. The lower inch of this bed is a purple orthoquartzite. Five successive conchostracan generations occur in this unit.
The mudcrack surface and ripple marks indicate deposition in the shallowest portion of a brackish-water cover with minor onlap and offiap controlled by slight oscillations of the sea.
(4) Frequently, the fossiliferous argillites are waxy to the touch, suggesting a relationship to the organic content; however, chemical determinations have yet to be made. It should be noted, however, that many waxy shales are barren of fossils.
Observations of ecdysis of living conchostracans in aquaria show that the creatures shed a very thin chitinous duplicature of the entire skeleton. These objects float for a while and then sink to the bottom muds. On the mud, they soon crumple or roll into an indistinguishable mass. Such chitinous material could be reworked by chitin-reducing bacteria although there is no direct evidence of the presence of such forms in Leonardian time. Chitin-reducing enzymes secreted by snails could also reduce such fine duplicatures. However, mollusks were not found in the Kay County sections.
Even explanations of this kind for fossiliferous waxy shales would still leave the barren waxy shales in the Wellington to be explained. Microscopic algae that seasonally cover all or a portion of many extant Kansas ponds may have had Leonardian equivalents. If so, that might account for the waxy shales. A few undetermined yet apparent algal filaments have been found in one or two Wellington sections in Sumner County but not in waxy shales. The only algae that have been found in Kay County, Oklahoma, and Sumner County, Kansas, were carbonate-secreters that formed reefIike and biscuitlike structures. Most of the waxy argillites in Kay County, surprisingly, lack spores or other palynomorphs. Therefore, in most instances, this type of explanation would be inapplicable.
(5) Grading into an argillaceous limestone in one bed of the Kay County section, was a sedimentary conglomerate of clay pebbles. Such objects were probably formed in shallow nearshore depressions that paralleled the shore or in desiccated portions of the mud bottom.
Interbeds and Sedimentary Events Indicated
The second question pertains to the types of sedimentary events that followed a given fresh-water biofacies in the Kay County section. One can take certain relatively closely spaced biotas as an example. The beds range from 62 (older) to 59 (younger). These are field numbers and are reversed for convenience.
Between bed 62 (a hard, argillaceous limestone), that contains insects and conchostracans, and the next overlying conchostracan-bearing bed, there occurs an interval of 0.1 foot of black, waxy shale. In turn, that is overlain by 0.1 foot of a hard, argillaceous limestone bearing conchostracans and xiphosura. That bed is separated from the next higher fossiliferous bed by 0.04 foot of a black, waxy shale. The next three fossiliferous units occur at approximately 0.1-foot intervals of shale or argillaceous limestone. A waxy shale 0.41-foot thick that overlies the upper of these fossiliferous beds is followed by another conchostracan bed.
Measurements indicate that the annual rate of sedimentation is about 0.02 inch for these Leonardian beds. Taking this figure, for the intervals given above that separate conchostracan beds, the time value in years (from older to younger) is: 60, 60, 24, 60, and 240. If the sedimentation rate is doubled, i.e., taken to be 0.04 inch as in the Kansas sections, these rates would be halved.
These intervals should be compared with those given in Table 5 for a single fossiliferous bed with 25 successive conchostracan generations in a sediment thickness of 3.2 inches. If multiple generations had occurred in successive seasons in the same pond, they would have been represented by separations between generations of as little as 0.02 inch. Because in actuality at the locality under discussion, they did not recur after such intervals, it is apparent that each of these fossiliferous beds represent a single season. At the end of one season multiple decades elapsed before a new pond site came into being at the same locality.
The most plausible explanation appears to be that the existence of pond sites, as noted earlier, was a fortuitous result of movements of an epeiric sea on and off a coastal flat.
In turn, this last observation may have value as a climatic indicator. If there were temporary ponds that seasonally dried up and contained evaporites during any of the 60-year intervals, such occurrences would denote a more arid weather cycle. In the absence of such evaporites or of evidence of small-scale wetting and drying during the 60-year intervals, a more temperate weather cycle would be indicated.
The point here is that there was a variation in weather during Wellington time; this can be inferred when thin units are studied that represent decades of time instead of millenia. The concept of deciphering, or at least securing a glimpse of "weather cycles" as distinct from overall climate in selected Paleozoic deposits, seems to merit additional study and may have applicability to Pennsylvanian and Permian cyclothems as well as older Paleozoic evaporite sequences. (Note: It should be noted that conchostracan fossils are present in many Pennsylvanian cyclothems and in such deposits as the Dunkard Group (Permian).)
In view of these considerations, the question asked earlier can be answered. The pond sites ended after a single summer. Decades to centuries of time separated successive pond sites at the same locality. The control appears to have been the minor fluctuations of an epeiric sea. Influx of occasional carbonized material indicates a vegetal cover in the swampy inland environs, and such debris was probably transported by small nonpersistent feeder streams.
Certain similarities to the Dunkard cyclothem (Beerbower, 1961) were suggested to the writer by the recurrence of carbonaceous beds at approximately 10-foot intervals above the gypsum in the Kansas Wellington sections (Tasch, 1963). Occurrences of conchostracans, fresh-water ostracodes and vertebrates, as well, in the Dunkard and Wellington seemed to strengthen this idea.
However, it remains to be seen by processing of all Wellington and Ninnescah samples on hand, and detailed comparisons to the Dunkard, whether the paleolimnological cycle can be made to fit into a modified Dunkard-type cyclothem. More importantly, it remains to be seen whether any new insights into environments, biotas, and sediments are thereby achieved. It may turn out that the concept of the "paleolimnological cycle" is a more productive approach.
Dunbar, C. O., 1924, Kansas Permian insects, pt. 1: Am. Jour. Sci., 5th ser., v. 7, no. 39, p. 175-208.
Raasch, G. O., 1946, The Wellington Formation in Oklahoma: Unpub. doctoral dissertation, Wisconsin Univ., 157 p.
Shaffer, B. L., 1961, Microfloral successions in Permian evaporites: Unpub. master's thesis, Missouri Univ., 89 p.
Tasch, Paul, 1960, Paleoecological observations on the Wellington Salt (Hutchinson Member): Kansas Acad. Sci. Trans., v. 63, p. 24-30.
Tasch, Paul, 1961, Paleolimnology: part 2--Harvey and Sedgwick Counties, Kansas: Stratigraphy and biota: Jour. Paleontology, v. 35, p. 836-865.
Tasch, Paul, 1962, Vertical extension of Mid-continent Leonardian insect occurrences: Science, v. 135, no. 3501, p. 378-379.
Tasch, Paul, 1963a, Paleolimnology: part 3--Marion and Dickinson Counties, Kansas, with additional sections in Harvey and Sedgwick Counties: Stratigraphy and biota: Jour. Paleontology, v. 37, p. 1233-1251.
Tasch, Paul, 1963b, Hystrichosphaerids and dinoflagellates from the Permian of Kansas and their paleoecologic significance: Micropaleontology, v. 9, no. 3, p. 332-336.
Tasch, Paul, and Zimmerman, J. R., 1961a, Fossil and living conchostracan distribution in Kansas-Oklahoma across a 200-Inillion-year time gap: Science, v. 133, no. 3452, p. 584-586.
Tasch, Paul, and Zimmerman, J. R., 1961b, Comparative ecology of living and fossil conchostracans in a seven-county area of Kansas and Oklahoma: Wichita Univ. Bull., Univ. Studies, no. 47, no. 36, 1, p. 3-14.
Tasch, Paul, and Zimmerman, J. R., 1962, The Asthenohymen-Delopterum bed--a new Leonardian insect horizon in the Wellington of Kansas and Oklahoma: Jour. Paleontology, v. 36, p. 1319-1333.
Dickinson County, Kansas
Elmo VIII-B-SW SW sec. 36, T. 16 S., R. 3 E., and offset.
Elmo III-C SE sec. 21, T. 10 S., R. 2 E.
Elmo Y-C-NE SE sec. 28, T. 16 S., R. 2 E.
Noble County, Oklahoma
Noble IA-D-SW NE sec. 34, T. 22 N., R. 1 W., and offsets.
Noble III-SE SW sec. 28, T. 24 N., R. 1 W.
Noble IV-SW SW sec. 23, T. 24 N., R. 1 W.
Noble II-A-NW NW sec. 4, T. 23 N., R. 1 W.
Noble II-A-SW SW sec. 23, T. 24 N., R. 1 W.
Kay County, Oklahoma
Wellington XIX-SE NW sec. 20, T. 29 N., R. 2 W.
Kay I-NW SW sec. 21, T. 25 N., R. 1 W.
Sumner County, Kansas
Wellington II-C-NW NE sec. 18, T. 32 S., R. 1 W., offset.
Wellington XVlII-NW NE sec. 23, T. 34 S., R. 2 E.
Wellington III-SE SE sec. 30, T. 32 S., R. 3 W.
Wellington XI-NW NW sec. 23, T. 34 S., R. 2 W.
Harvey County, Kansas
Annelly 1-A-NW NE sec. 21, T. 23 S., R. 2 E.