
|
Original published in D.F. Merriam, ed., 1964, Symposium on cyclic sedimentation: Kansas Geological Survey, Bulletin 169, pp. 87-106 | |
University of Oklahoma Research Institute, Norman, Oklahoma
New data on the occurrence of ventricose fusulinids (and of similarly shaped modern foraminifera) indicate the possibility of deeper rather than shallower waters for their normal habitat than is currently suggested; but slender fusulinids appear to have inhabited substantially shallower waters than the ventricose forms.
Latest research on stromatolite-forming filamentous micro-organisms induces serious doubt regarding the validity of their current biological understanding and paleoecological interpretations.
Historico-geological and hydrological considerations demand that the Kansas Sea attained substantially greater depths at the climaxes of the late Pennsylvanian-early Permian oceanic invasions than the currently suggested 80 feet; and ventricose fusulinids apparently entered the Kansas Sea at or about the climaxes of the invasions.
Restoration of the Kansas Sea during Beattie time suggests that the southwest is the only possible way for the oceanic transgressions.
The mechanism of "staggering" oceanic transgressions over neighboring Mediterranean basins, such as the eastern Tethyan Black Sea-Caspian Sea-Aral Sea, is here offered as a plausible explanation for the megacyclic sedimentation typical of the Kansas Sea.
[Note: I am indebted to Dr. Helen Duncan for editing the manuscript.]
In 1937 I attempted to reduce the limits of probable maximum depth of the Kansas Sea in Big Blue (Lyonian) time to 180 feet on the basis of the apparent cyclic repetition of the marine biota in the successive Big Blue sediments. The principal evidence for this interpretation was furnished by the centralized position of the fusulinid phase in the cyclic succession of the biota, which apparently reflects periodic invasions of the marine waters during late Pennsylvanian and early Permian time. The maximum depth of the habitat of the modern large Foraminifera, as recorded in the literature of that time, was used in setting the probable maximal depth at which fusulinids lived; neighboring, shallower bottoms, were apparently populated largely by bryozoans and brachiopods.
Mudge and Yochelson (1962, p. 114) have mentioned that, "Since 1937, considerable information has been gathered about the life habit of the larger foraminifers," and they cite recent evidence on the genus Cycloclypeus, occurring at depths of 180 to 1,200 feet and most commonly between 240 and 600 feet, and the occurrence of the abundant Fleterostegina and Operculina "from near low-tide lines to at least 240 feet (Ruth Todd, oral communication) ."
However, more significant paleoecologically are the findings by Bandy (1960), that the:
. . . relationships between environment and the composition, structure, and form of Foraminifera shows striking correlations in all of the major groups . . . [and that their evolutionary] convergence provides one important explanation of many of the correlations between foraminiferal morphology and environment. Spindle-shaped types are characteristic of depths between 20 and 80 meters [60 and 240 feet] in modern tropical and subtropical oceans. This same association is suggested with respect to the fusulinids of the later Paleozoic. . . .
These data may be profitably combined with the conclusions by Rauser-Chernousova (1951; review by Elias, 1959, p. 55) regarding the habitat of the fusulinids in the Ishimbaevo submarine plateau:
The submarine plateau (with the exception of its fringes, where reefs predominate) has been a seat of a facies of shallow water banks. This facies is subdivided into two sub-facies: (a) mizzia (dasyclads)-bryozoan submarine banks with depths less than 40-50 meters (120-150 feet), and (b) algae submarine banks deeper than 40-50 meters (120-150 feet).
The first subfacies is usually encountered in the lithotopes formed by the interstratified organic-detrital limestones of very diversified kinds with thick biostromes: hydractinoid, mizzia (siphonaceous algae) and bryozoans, and corals. Small foraminifera are fairly diversified, and fusulinids are frequent, being represented largely by elongate forms; bryozoans are moderately distributed; crinoids are not infrequent . . .
The second subfacies, at depths greater than 40-50 meters typifies quieter and more uniform conditions of sedimentation and of life. Possibly there were some periodic fluctuations toward colder temperature, which are indicated by several layers with gregarious Glomospira. Sediments are usually biomorphic, complete skeletons prevailing over detritus. Spotty, reef, and encrusting structures are not infrequent. Limestones are distinguished by their massif character, absence of stratification and sorting of grain, and by purity and considerable thickness. Reef-forming hydractinoids, bryozoans, and corals are very rare, but algae have flourished, the white tubules of shamovella (=Tubiphytes) occasionally encrust a rock known as wormy (cherviachnyi) limestone, or even forming lenses with sod-like constructions and showing beautiful encrusting structure. Besides the extremely diversified calcareous algae, this facies not infrequently contains small Foraminifera, with prevailing Glomospira, Tolipammina, small Ammodiscus, and Nodosaria. Fusulines are poorly represented, and are quite absent in the beds rich with shamovells (=Tubiphytes). But in the neighboring layers, with richer detritus, fusulines are richly represented, mostly by inflated to spherical forms (in the middle zone of the Schwagerina horizon), and among the small Foraminifera are found the large, also inflated thick-walled forms of Climacammina. The presence of such thick-shelled, shortened forms may be an indicator of conditions approaching those in reefs. Characteristic of this subfacies is its widespread distribution, and building of expansive submarine banks with fairly uniform and stabilized lithologic and paleontologic character.
It is important to add, that this facies of shallow-water banks, with its two sub facies just described, is set apart by Rauser-Chernousova from the facies of shallow banks to which
. . . belong the sectors of the reefs and submarine banks that are shallower, thus suffering noticeable influence of strong movements of water. Typical sediments of this facies are calcareous sands composed of well-rounded particles of organic nature (not infrequently hydractinoid sand), which are conspicuous by the light color of their particles. . . . Fine gravels, composed of either organic remains (frequently of fusulines rounded into flat or round pebbles), or made of ooze with organic remains embedded in it. Coprolite-nodular ooze is very common in this facies. . . . Characteristic for this ooze are two elements: (1) well delineated oval to subspherical coprolites about 0.5 to 1 mm in diameter, made of dark fine-grained matrix mixed with fine organic particles, and (2) nodules of blue-green algae, which have a similar fine-grained structure, but whose contours are less sharply defined. In the coprolite-nodular oozes are encountered Glomospira, Tolypammina, Ammodiscus, and Hemidiscus, and various encrusting and adnate blue-green algae, whose entangled thin filaments preserve their form well. Fusulines are also characteristic for this facies, with a predominance of more inflated forms with reinforced shell, the same kind as encountered in the reef facies. It is possible, however, that these latter forms were merely transported and buried in this facies, which is not very favorable for their habitat (Rauser-Chernousova, 1951; review by Elias, 1959, p. 55-56).
Rauser-Chernousova did not indicate approximate depth of this facies of shallow banks, but her statement that its "banks are shallower, thus suffering noticable influence of strong movements of water," clearly contrasts with her preceeding description of the first subfacies of the shallow-water banks with depths of less than 40 to 50 meters (120 to 150 feet); and her description seems sufficient for an evaluation of the depth for her facies of shallow banks, with strong water movement, as hardly more than 80 to 90 feet.
It is important to note, that the facies of shallow banks is considered by Rauser-Chernousova "not very favorable" for the inflated fusulinids with reinforced shell, which predominate among the fusulinids encountered; and that she considers them as possibly transported to the facies from elsewhere. If so, the facies may be considered to contain only rare fusulinids of its own, if any. Furthermore, even if she prominently mentions the frequent occurrence of the rounded fusulinid pebbles in the fine gravels of the facies, she obviously does not consider the encountered intact fusulinids to be necessarily a part of its live population. Because her observations and conclusions are based on material from a broad and densely core-drilled area (which became the principal producer of petroleum in the USSR), her data seem to be particularly important in considering the paleoecological significance of the fusulinids in sediments of similar nature and age in America.
Important observations on the West Texas fusulinids were published by Ross (1963, p. 53):
The shape of the fusulinid shell corresponds closely with the kind of rock in which it occurs. The long subcylindrical shells of such species as Triticites ohioensis Thompson, T. comptus Ross, T. primarius Merchant and Keroher, Schwagerina gracilitatis Dunbar and Skinner [closely related to and similar in shapes to S. emaciata (Beede)], and Monodiexodina linearis (Dunbar and Skinner), are common in sandstone with a high percentage of clay minerals. Stratigraphic relations of these rocks suggest near-shore, shallow-water deposition. . . These subcylindrical species were probably benthonic, and their shapes prevented transportation by currents for any great distance.
The subventricose species, such as Triticites cullomensis Dunbar and Condra, T. beedei Dunbar and Condra [both from the Upper Pennsylvanian of Kansas and Texas], and T. milleri Thompson, on the other hand, are most common as local accumulations in fine-grained calcareous strata. These species probably preferred less agitated water than the subcylindrical species. . . .
Large globose species, such as in the genera Pseudoschwagerina and Paraschwagerina, are common in and adjacent to bioherms of the Neal Ranch [lower Wolfcamp] and Lenox Hills [upper Wolfcamp] formations. Many. . . are crushed or fragmentary, and they must have been transported with other shell fragments away from the bioherms into the debris aprons. . . .
Ross considers that the globose species "may not have been bottom dwellers. Their thin walls and large volume per shell weight may have permitted them to climb and crawl on the sessile crinoid stems, bryozoan colonies, and productid brachiopod spines"--a challenging suggestion.
Other observations by Ross indicate a high selectivity of environments displayed by some ventricose species, particularly among the diversified Permian forms.
Although Ross does not suggest an approximate depth of bottom for the three principal categories of the fusuline shapes, which he calls subcylindrical, subventricose, and globose, it is apparent from his description of their environments that they preferably occupied the bottoms of about the same depths determined by Rauser-Chernousova for the contemporaneous Russian fusulinids of the corresponding shapes: elongate, inflated, and spherical.
It seems useful to add here the following complementary remarks selected from the annotations in Dunbar's bibliography (1957) on fusulinids. According to Kahler and Kahler (1947), who published:
Fusulines are rare where pelecypods, gastropods, brachiopods, or crinoids abound, but may be plentiful in association with certain algae, with scattered brachiopods, and scattered corals, and with sponges. It is inferred that they avoided areas that were strongly colonized by larger organisms that used much calcium carbonate. Their preferred habitat was warm, shallow and quiet near-shore water, even where the oxygen content might be somewhat deficient (Kahler and Kahler, 1947; from Dunbar, 1957, p.754).
On the evidence of the fusulines from the Permian of Indochina, Gubler (1935) concludes that: "Fusulinidae and Neoschwagerinidae did not live together, the latter dwelling farther from shore and perhaps in deeper waters than the former" (from Dunbar, 1957, p. 754).
Thompson (1948) concludes that fusulines were restricted to offshore open-water enviroments, and penetrated farthest into the continental basins when seas reached their farthest advance (Dunbar, 1957, p. 754). Dunbar himself states that:
Where fusulines occur in abundance, . . . the majority of shells are adult, unbroken, and free of abrasion or wear. Where the shells are mostly adult those of each species are similar in size, but commonly two or more species of very unequal size occur abundantly together. From these facts it is inferred that they lived and accumulated on a quiet sea floor free from active agitation by waves and free from bottom currents capable of transporting and size grading the empty shells. Exceptions are known to this generalization, notably in the Permian limestones where in places the long cylindrical shells of Parafusulina and Polydiexodina lie parallel as though rolled by gentle bottom currents. . . . (Dunbar, 1957, p. 753.)
Parallel arrangement of a few adjacent individuals occurs, even if rarely, among the profusely accumulated shells of the very slender Triticites cf. irregularis Dunbar and Condra in the upper half of the Toronto Limestone (lower Oread) east of Lawrence, Kansas.
Concurrently with the accumulation of information on the environment and depth at which the Recent and Paleozoic spindle-shaped foraminifers are encountered, considerable progress has been made in the very difficult subject of the biology and ecology of the primitive filamentous algae which are found in various calcareous encrustations in modern and ancient seas.
A good summary on the latter subject, "based in part on fossil and recent specimens, but mostly on the literature" was published by Cloud (1942). Cloud's review was an important supplement to my use of Edward Forbe's conclusion, "concerning predominance of leathery seaweeds to a depth of 90 feet, and the common occurrence of calcareous seaweeds (encrusting Lithothamnium and branching Corallina) below this depth" (Elias, 1937, p. 419-420). However, I have shown in the restoration of the sea bottom zones in Big Blue time in Kansas (1937, Fig. 3, p. 410), that the leathery seaweeds of that time rather occupied the bottoms of 30 to 75 feet depth, and the calcareous algae 75 to 110 feet depth, as determined on the evidence of the associated invertebrates. I also remarked in the text (1937, p. 420) that the encrusting structures suggestive of lime-secreting red algae of the Lithothamnium type are occasionally found in the Big Blue series and are common in the underlying Pennsylvanian rocks of Kansas. They are found in association with pelecypods, brachiopods, and some even with fusulinids. Actually, there was no evidence at that time, and none at the present, that any of the algaelike filamentous structures in the late Paleozoic encrustations known under the form-names Osagia, Ottonosia, and Cryptozoon, have a definite affinity to red algae. They are now generally believed to belong probably to blue-green and partly to green algae, but this also is not certain. In his discussion Cloud wisely calls them stromatolite-forming algae (1942, p. 371), and concludes that the "stromatolites in most instances probably indicate growth at a depth of less than 30 meters (90 feet) in salt water or 10 meters in fresh water" (1942, p. 363), a conclusion which is still generally acceptable.
In a special paper devoted to the Permian and Pennsylvanian algae of Kansas, Johnson (1946, p. 1103) demonstrates that the:
. . . colonies classed as Osagia consist of intimate intergrowth of algal filaments of different sizes presumably representing algae of several types. Permian specimens consist of intergrowth of such algae with coarse tubular foraminifer belonging to the genus "Nubecularia". The nature of assemblage varies in different formations. . . .
He shows also (1946, p. 1114) that the microstructure of Ottonosia laminata Twenhofel consists of the laminae "composed of a mat of branching algae filaments," and he (1946, p. 1106) describes Cryptozoon Hall as having "laminae composed of fine, mostly branching threads of simply organized algae, probably Cyanophyceae [blue-green algae] ." Johnson describes also small lumps and crusts made of tubular filaments and classifies these in the genus Girvanella Nicholson and Etheridge. Johnson (1946, p. 1108) summarizes his ecological observations on algae thus:
. . . (1) Some algae were present in all the marine environments involved; (2) the majority of lime-secreting algae are associated with molluscan and mixed faunas [Elias, 1937, p. 411]; (3) some algae occur with the presumably deeper-water fusulinid faunal group; [and then adds that] . . . most of the algal limestones occur in the bottom and top limestone members of a limestone formation (Figs. 2-5). This confirms the observation of Elias (1937) on the Permian Big Blue series. The exceptions are certain phases of the fusuline.bearing limestones which may develop highly algal facies.
Johnson describes six species of the dasycladacean green algae Epimastopora Pia, nearly all from the basal members of the limestones in the cyclothems; and five species of his genus Anchicodium, which he believes to be related to the modern Codium, also a green alga. However, a closely related, if not identical genus Ivanovia Khvorova (1946), described in a paper published 4 months prior to Johnson's Anchicodium, is considered by Khvorova to be "most like a brown or red alga," but Maslov places it in Siphonales of green algae. On the other hand, a very similar Eugonophyllum Konishi and Wray (1961, p. 662) is described as having presumably fertile structures which "closely resemble the conceptacles of the Fucales (Phaeophyceae), and their growth form similar to that of some Phaeophyceae, e. g. Thalassiophyllum clathrus (Gmelin) ." Eugonophyllum also resembles Thalassiophyllum clathrus in having a perforated thallus.
The possibility that Ivanovia as well as Anchicodium and Eugonophyllum (which may be considered its subgenera) are not green, but rather brown algae (Phaeophyceae) has some support in their apparent general occurrence in waters somewhat deeper than those favored by green algae. Johnson's reported occurrence of the species of Anchicodium places them about equally in the lower, middle, and upper parts of the limestones in the megacyclothems. In the book on the Middle and Upper Carboniferous of the Moscow Basin, Ivanova (1955) considers the development of the extensive bioherms built by Ivanovia tenuissima Khvorova, "the platy brown ( ?) calcareous alga," as belonging to the waters "at a fairly great distance from shore, beyond the reach of terrigenous mud, and at a comparatively great depth" (Ivanova, 1955, p. 57). The meaning of the expression "comparatively great depth" is elucidated in a sketch that shows the schematic facies position of the algae in the Middle Carboniferous sea of the western part of the Moscow Basin (1955, Fig. 29). It shows a zone of brown algae (Ivanovia) mixed with some siphonaceous green alga Dvinella Khvorova, between the sediments of the open sea shoals and their slopes, and the comparatively deep water zone, the deepest in the local Mid-Carboniferous sea. This deepest zone is characterized by small-to-fine-grained calcareous ooze, with a fauna mostly well preserved and fairly diversified, but poor in number of species representing each phylum: brachiopods, echinoids, crinoids, and comparatively rare horn corals, bryozoans (mostly fenestrate forms), gastropods and other molluscs, fishes, and sponges. Foraminifers are abundant, with a "dominance of small forms among the fusulinids and other families" (Ivanova, 1955, p. 51).
Similar environments are postulated for Eugonophyllum, whose "bioherms probably originated as broad, moderately low relief submarine structures, their top possibly extending to but generally occurring below the vigorous wave action" (Konishi and Wray, 1961, p. 664).
The opinions expressed by Ivanova (1955), and by Konishi and Wray (1961) do not seem to harmonize with the belief by Imbrie and others (1959) that the Anchicodium facies of the lower half of the Cottonwood Limestone in southeastern Kansas has developed in very shallow waters of the postulated Greenwood Shoal, shown as only about 30 feet below surface water level (Imbrie and others, 1959, p. 71). This same profile with the same interpretation is also shown by Laporte (1962, p. 524), who describes the Anchicodium platy algae facies as formed in "an area of moderately turbulent shallow, well-lit water, with some restriction of circulation" (Laporte, 1962, p. 540).
Most interesting is Laporte's statement (1962, p. 525) that "the concept of the Greenwood Shoal is critical in understanding Cottonwood facies." It seems that the evidence favoring the existence of this shoal, as discussed by Laporte (on the same page), and also by Imbrie and others (1959, p. 74-76) is quite substantial, except it does not supply the evidence for as shallow waters over the shoal as shown on the quoted profile. It is sufficient, however, for the conclusion that the widely developed but low structures, built by the platy alga in the Greenwood Shoal stood higher than the adjacent bottom to the north, where the Cottonwood fusulinids flourished. As will be discussed subsequently, the whole concept of the extremely shallow Kansas Strait, which supposedly was never deeper than 80 feet even at a culmination of the Beattie marine invasion (as shown in the quoted profile, and implied in the text), is out of harmony not only with the evidence of the inflated (ventricose) fusulinids ecologically comparable to those of the Cottonwood and Morrill Limestones, but is also irreconcilable with the hydraulics of modern epeirogenic seas, as well as with the direct evidence of the repeated channeling cutting deep into the marine sediments of the Kansas Sea up to early Lyonian time. The conflicts just enumerated do not negate the validity of nearly all observations made by Imbrie and his associates, but necessitate a re-evaluation of their significance. It appears particularly necessary to review critically the much too broadly applied term Osagia, which has been used indiscriminately for all kinds of organically induced encrustations, including those around the Cottonwood fusulinids.
In 1958 Ginsburg and Lowenstam made the statement (1958, p. 316):
Fossil algal mats--stromatolites--have long been known. . . There is at present little doubt that their structures were produced by blue-green algae. However, many paleontologists interpret these structures as produced by algal precipitation, and the physical sediment-fixing aspect first described by Black (1933) has often been overlooked. . . .
They also summarized the following important data on the blue-green algae (Cyanophyceae) which should be considered in their identification (1958, p. 311):
Blue-green algae. . . are extremely abundant in the tropical oceans both as plankton, which is the first link in the food chain, and as sessile forms on and within the bottom sediments and rocks. Furthermore, these ubiquitous organisms do not require [for their growth] constant submergence or a specific salinity. . . . These algae have a thick external capsule or sheath of mucilaginous cellulose. In the colonial growths that form the film, the mutual adherence of these sheaths and the intertwining habit of the filamentous forms produce a tough rubbery mat that is not easily broken.
According to observations by Black (1933), the sediment particles are securely bound by the intertwining of the filamentous algae, some of which are mobile and can move over and around the particles. This binding can be very rapid and in the laboratory "a lamina of sediment, a few millimeters in thickness, could be permeated and bound fast by filaments of Schizothrix, Symploca, or Phormidium in a few days" (Black, 1933, p. 178). Similar observations of a culture of Phormidium were made by Ginsburg and others (1954, p. 27). Illing (1954) calls attention to a considerable similarity between the algae threads in the modern algally encrusted lumps of the Bahama Banks and the similar threads in the Permian and Pennsylvanian limestone of Kansas. These Bahamian lumps (Illing, 1954, p. 31-32):
. . . reveal their algal encrustation by a faint green or brown discoloration. . . . Clear cyanophyceous threads may be seen growing on them, and entrapping small grains and silt particles. Isolation of the algae by decalcification was not very satisfactory, but they appear to ramify throughout the lumps from the rather friable surface to the more densely cemented material beneath. . . . They are about 5 microns in diameter, and are closely packed together, as in Plate 4.2 (. . . very similar to illustrations of algal threads from Pennsylvanian and Permian limestones of Kansas by Harlan Johnson--1946, p. 1107, Pl. 9, Fig. 3, 4. . .). It seems likely that they play an important part in the biochemical extraction of the calcium carbonate which surrounds them. As the distinction between this type and the broader filamentous boring form is not clear, there is no sharp division between "algally encrusted lumps" and the featureless type here described. They have been separated in the tabulations of sediment contents, in order to avoid classifying lumps as being formed by algal action in the absence of external visible evidence.
Attempts to grow cultures of the algae in the laboratory were not successful, and it has not been possible to proceed with their identification. Probably several different genera and species are present.
In view of Illing's statement and in the light of the preceding observations by Black (1933) and by Ginsburg and Lowenstam (1958), one may question whether the clear cyanophyceous threads mentioned in the first part of Illing's statement are truly referable to the blue-green algae. Still more difficult to determine is the provisional classification of the threads illustrated by Johnson (1946, Pl. 9, Fig. 3, 4).
Even the long standing belief, that the best known Paleozoic filamentous alga Girvanella belongs to blue-green algae is no longer maintained by the most enlightened authorities on the primitive filamentous-tubular forms.
Thus, Alan Wood (1963) in his latest revision of Girvanella and related genera, refers to them merely as calcareous algae, whereas Henbest (1963, p. 22) in his taxonomic understanding of Osagia and Ottonosia speaks about Girvanella and similar filaments in these form genera with even greater reticence; these form genera are "colonies of algae or of algae and Cornuspirinae, assuming that Girvanella and such are algae." Indeed, Henbest has good reason to say so, as his superior technique in staining microscopic organic structures, and in photographing them at x 40 magnification, and their extra thin sections at x 1000, enables him to reveal a much greater diversity of such minute organisms and their structure, than ever known before.
Our progress in understanding of true nature of Paleozoic encrustations, originally given generic dignity under the names Osagia and Ottonosia, may justly be called spectacular. What was at first considered a single even if peculiarly protean alga (Twenhofel, 1919), became known as a combination, perhaps symbiotic, of an irregularly multicellular foraminifer identified with the Cretaceous genus Nubecularia (Johnson, 1946, p. 1103), and filamentous alga classified in the broadly interpreted genus Girvanella, supposedly a blue-green alga. Now "Nubecularia" of Johnson (as well as its Russian equivalent, Paleonubecularia Reitlinger, 1950) is shown to be, similarly a combination of irregularly squirming sessile tubular genera of Cornuspirinae, associated with finer tubules or threads (known as Girvanella sensu lato), which do not seem to be truly blue-green algae, and even may not be algae at all. Hence the argument that these accumulations required good illumination for algae growth, in as far as applied to Osagia and the Osagia facies, is no longer valid.
I (1962, p. 114) have already commented on the statement by Lane (1958), that:
. . . Gentle wave or current action, easily developed at a depth of 60 feet, is necessary in that the shell particles must have been turned over from time to time to allow algal growth on all sides.
I can now add to my previous comment, that stronger and quite constant turning over can be provided by crawling and burrowing invertebrates at and beyond wave and current disturbances. An example of the magnitude of such disturbances is provided by the following direct observation cordially supplied by Dr. Jean M. Berdan (written communication, 1962) of the U. S. Geological Survey, concerning the burrowing habits of Limulus in captivity:
Four specimens of Limulus, the largest of which is now 8 cm long, have been in a 25 gallon tank in the U. S. National Museum since September, 1960. The water in the tank has not been changed during this time, and as a result, the sand at the bottom is contaminated with organic matter, and is dark in color. However, the upper part of the sand, to a depth of 1 cm, is much lighter in color, because the animals plow back and forth across the bottom searching for food and constantly turn it over. Although the same sand is agitated more often by one animal in confinement, presumably large numbers of them might produce the same effect in a state of nature.
Laporte (1962, p. 530) describes a "fusuline-rich layer, 1.3 inches thick, which occurs at the top of the Cottonwood from central Kansas to northern Oklahoma, almost to the southern limit of its exposure. Each of the Cottonwood facies found in this region--platy algae, shelly, and silty Osagia--has thin layers; except for the abundance of fusulines, the lithologic and biologic character of the layer in each facies is typical for that facies. In this same area, the lower part of the overlying FIorena Shale has a well developed fusulinerich horizon, which is a continuation of the fusuline-rich layer at the top of the Cottonwood."
I have pointed out already (1962, p. 100) that in the diagram of the taxonomic composition of biota (Laporte, 1962, Fig. 4) in locality 15 of the Cottonwood Limestone, which is about 30 miles south of the southern edge of the platy algae facies development, a prominent fusuline biota is shown well developed in the uppermost 1 foot (or more) of the 6 foot section, whereas the same section of locality 15 is interpreted in Laporte's Figure 3 (1962, p. 526-527) as fully in the shelly facies. This topmost one foot thick layer, with abundant to dominant fusulinids in the southern province Cottonwood Limestone, is substantially thicker than, and cannot be the same as the fusuline-rich layers, 1-3 inches thick, which Laporte observed at the top of the Cottonwood; and therefore it appears rather to correspond to the same fusulinid phase of the invading sea, which is indicated by the 3 foot thick occurrence of the fusulinids in the northern province, as portrayed in the section of the localities 5, 7, and 10 in the diagram of Laporte's Figure 4.
This fusulinid phase of the Beattie marine transgression is by no means a nearshore development, as it stretches for at least 200 miles across the Nebraska Shelf from the restored northern boundary of the Kansas Strait (Imbrie and others, 1959, p. 70-71; Laporte, 1962, p. 523-524). It is, of course, a deeper phase in the Beattie marine transgression than the bioclastic facies of the lower half of the Cottonwood Limestone, an opinion apparently no one disputes.
Thompson (1954) describes from the upper part of the Cottonwood Limestone Schwagerina jewetti Thompson, S. kingi Dunbar and Skinner, and "Triticites sp.," all typically ventricose forms. This is also the shape of the single axially sectioned fusulinid in the slide illustrating the fusulinid facies from the same upper part of the limestone (Laporte 1962, Pl. 2, Fig. 2, left upper corner). The fusulinids that I observed in the Cottonwood Limestone were invariably ventricose, and I see no reason to change my 1937 opinion that this fusulinid phase of the Beattie cyclothem represents the culmination of marine invasion.
Quite different is the shape of Schwagerina emaciata (Beede), the first species described from the Beattie Limestone and traditionally attributed to the Cottonwood Limestone. Actually, it is this elongate to subcylindrical form that is frequently encountered (in my observation) at the top of the Cottonwood Limestone and the base of the overlying FIorena Shale, where fusulinid specimens are easily obtained from the matrix. There is hardly any doubt that this form is a predominant, if not exclusive, fossil in what Laporte differentiated as a "very thin fusuline-rich layer at the top of the Cottonwood Limestone . . . from central Kansas to northern Oklahoma, almost to the southern limit of its exposure" (Laporte, 1962, p. 530). In the light of the previously mentioned observations by Rauser-Chernousova the fusulinids of this elongate shape belong to bottoms less than 120 to 150 feet (Rauser-Chernousova, 1951; review by Elias, 1959, p. 55). It is therefore reasonable to suggest, that the fusuline-rich layer at the top of the Cottonwood and the base of the FIorena, which is properly not included by Laporte in the underlying fusulinid facies (Laporte, 1962, p. 530), provides evidence of the retreat of the sea, subsequent to the culmination of its inundation at late Cottonwood time.
Further retreat appears to be indicated by the richly fossiliferous, but fusulinid-free, middle and upper FIorena Shale, where the mass occurrence of Chonetes granulifer inspired the useful quantitative research by Imbrie (1955). Perhaps it is the central position of the Chonetes facies, between the fusulinid occurrences below (in the Cottonwood) and above (in the southern exposures of the Morrill), that prompted Imbrie to decide that the "Chonetes facies, rather than the fusuline limestone, should be taken as representing the climax of marine transgression" (Imbrie and others, 1959, p. 78). However, in other Lyonian marine invasions, notably in the Foraker Limestone cycle, the fusulinid and brachiopod phases are repeatedly interstratified, thus indicating, in my opinion, minor fluctuations of the sea level during a prolonged major marine invasion; and it seems that the recurrence of the single ventricose fusulinid species in the southern facies of the Morrill Limestone, and described by Thompson as Schwagerina vervillei Thompson (1954, p. 62-63), a probable descendent of the S. jewetti from the Cottonwood, indicates a second, short duration sea inundation of the Beattie megacyclothem.
If I am correct in this interpretation, the Kansas Sea tended to become deeper toward the south as compared with the north even in early FIorena time. This understanding is suggested by the northern limit of the fusulinid-rich layer in central Kansas, whereas the underlying fusulinid facies of the Cottonwood extended far into Nebraska, but was poorly developed south of Greenwood Shoal. Then, at Morrill time, the last fusulinid appearance was limited to the southern edge of Greenwood Shoal and to the Arkansas Embayment. Such evidence may be considered to indicate further lowering of this area in comparison with that in the north. The suggested interpretation is harmonious with the postulated differential uplift and subsidence suggested by Imbrie and others (1959, Fig. 2, p. 71; see, also Laporte, 1962, Fig. 2).
As I have already commented (1962, p. 110): "Chonetids generally tend to be gregarious and when occurring in great number and not mixed with other invertebrates, they seem to indicate waters shallower than normal for the majority of articulate brachiopods."
I may add in support of this conclusion a discovery of three nested chonetid valves, with the convex sides facing downward and the size of the valves diminishing upward. This valve-in-valve arrangement came from the mass occurrence of Chonetes in the basal part of the Plattsmouth Limestone 1 mile south of Plattsmouth, Nebraska. I failed to understand the significance of the arrangement until recently; in the tide zone of Galveston Bay at Galveston, Texas, I observed a similar valve-in-valve arrangement of two, and occasionally three valves of various pelecypods, with convex sides invariably facing downward. Whereas most of the disarticulated valves were resting in their normal position of stability, with convex sides facing upward, the few that managed to land in the bowls of larger valves (usually together with some sand), added weight to the latter, and hence helped them to maintain their position with the convex side facing downward; and made it possible for a still smaller valve to land within the bowl of the second valve. In order to achieve this valve-in-valve arrangement, gently agitated water appears essential, in order to lift one valve above the edge of another, so that it could come to rest in the hollow of a larger valve. In the light of this observation the Plattsmouth chonetid valve-in-valve phenomenon appears to indicate deposition in water that was gently agitated, not necessarily within the tidal zone, but at a depth where wave action affected the sea bottom.
The problem of depth of the Kansas Sea is a part of general problems of its hydrology, and should be viewed in a historical retrospect of the late Paleozoic Mediterranean of America, comparable to the Tethys of the Old World. The attempt at reconstruction of the hydrology of the Kansas Sea at Beattie time by Imbrie and associates is commendable, but perhaps they have not reckoned with the hydrological and general geological implications, entailed in postulating an extremely shallow maximal depth at the culmination of the Beattie marine invasion.
In a profile across the Nebraska Shelf and Arkansas Embayment (Imbrie and others, 1959, Fig. 2, p. 71; Laporte, 1962, Fig. 2, p. 524), the greatest depth is shown to be about 80 feet, in the middle of the Arkansas Embayment. If this were so, the average southern slope, for a distance of about 100 miles to the postulated shore, would be about 1 1/4 foot per mile, and the northern slope across the Nebraska Shelf, about 1/2 foot per mile, for a distance of about 180 miles to northern edge of the Greenwood Shoal.
There are only very few existing slopes of similar average gentleness for comparable long distances of scores of miles. Such are the slopes in the northeastern and southeastern corners of Great Salt Lake (1/2 to 3/4 foot per mile), which are the sites of discharge of Bear River and Jordan River respectively; in the mangrove swamps (1 foot per mile) of Gulf of Mexico; in the great deltas of Mississippi and Volga Rivers, and in the drowned limestone plateau off the west coast of Florida (1 foot per mile).
If the maximum depth of Kansas Sea, as reconstructed, were to be estimated at about 180 feet, as postulated by me in 1937, the slopes would be about 3 feet and 1 1/2 feet per mile respectively. These figures are more nearly comparable to those of the gentlest slopes on either side of Volga River delta in the northern Caspian Sea, and in the northwestern part of Gulf of Mexico, and to some slopes in the western part of Great Salt Lake.
Direct evidence in favor of the greater depth for the Kansas Sea in Lyonian time, and of still greater depth (at least 200 feet, and perhaps as much as 250 feet) in the Late Pennsylvanian Sea, is the existence of some deep channeling across the marine sediments of the Kansas Sea. The deep channeling at the beginning of Missourian time was described by Hinds and Greene (1915); and channeling that occurred repeatedly during Wabaunsee and early Lyonian time was described by Mudge (1956; also Mudge and Yochelson, 1962). The great Pennsylvanian channeling, as indicated by the Warrensburg and Moberly channel sandstones in the basin of the Missouri River in Missouri, was 2 to 3 miles wide and had a maximum depth of 175 to 200 feet (Hinds and Greene, 1915, p. 95-102) and belonged to a single drainage.
. . . the Warrensburg channel was made by water flowing from higher country on the Ozark dome, bringing with it sands and muds derived largely from early Pennsylvanian [Cherokee] sediments. The Warrensburg stream was joined when it reached the present site of Missouri River by the [ancient] Moberly River descending westward from an Ozark peninsula in northeastern Missouri, and the united streams continued northward or northwestward to the open sea. This condition did not last long, for the valleys have all the characteristics of a topography as youthful as that of the Red River of the North--deep trenches in level plains with very little lateral drainage. Shortly before the land was again submerged the channels were choked with sediments, swamps with thin peat beds were formed on the flood plains, until finally the sea again swept in and covered them. . . .
The writers feel justified in tentatively referring the channel deposits to the Pleasanton epoch (Hinds and Greene, 1915, p. 93-94).
The origin of the channel sandstones of Virgil-Big Blue time is probably generally similar to that of the Warrensburg-Moberly channel sandstones, that is subaerial channeling followed by sedimentary choking. However, the emergent areas were apparently lower, and the ensuing erosion probably was generally less. Just how much less may be judged by the decrease of the maximal depths of the sandstone channeling, estimated by Mudge and Yochelson (1962, p. 16-17) to be about 140 feet for the Pony Creek (Wabaunsee) Shale channel, and to about 85 feet for the Towle Shale (basal Big Blue) channel.
The gradual diminution of the maximal depth in the Kansas Sea toward the end of Lyonian time is indicated by the orderly disappearance of the major groups of marine invertebrates in the last invasions of the sea; no fusulinids subsequent to the Barneston megacyclothem (the miniature fusulinid named Oketoella, was discovered by M. L. Thompson in the Oketo Shale, but its occurrence hardly indicates renewal of marine invasion at this time); no brachiopods and bryozoans after the Nolans cyclothem; and no large molluscs after the Donegal cyclothem (Elias, 1937, p. 405-408). It seems that the maximum depth of the Kansas Sea approached 80 feet at this latter time, which was followed by an epoch of still shallower waters, with the major development of redbeds and the accumulation of evaporates in isolated lagoons.
The published paleogeography for the Beattie Limestone Sea does not seem harmonious with some pertinent geological data, particularly with respect to the southeastern and southern parts of the sea. The original paleogeography by Imbrie and others (1959, p. 70, Fig. 2, p. 71) already has been somewhat modified by Laporte (1962, Fig. 2, p. 524) , who states that whereas (p. 523): "The Kansas Strait had its principal connection with the open ocean through the Arkansas Embayment on the southeast; a lesser connection may also have existed to the south-west. . . ."
However, Laporte's roughly indicated southwestern connection for the Kansas Strait has but little resemblance to any of the progressively retreating shorelines illustrated by Tanner (1959, Fig. 5, p. 334), and acceptance of anyone of them effectively eliminates the supposed southwestern shore of the Arkansas Embayment. The postulated northeastern shore of this same embayment, when superimposed over the geologic map of Missouri, runs across the west-central part of the stable Ozark Dome. Furthermore, there is no direct evidence that any sediments of Permian, or even medial and late Pennsylvanian age, ever covered northwestern Arkansas and the adjacent territory. Hence not only the postulated shores, but even the very existence of Arkansas Embayment lack geological support; whereas the isopachous patterns of the Florena Shale east of Cottonwood outcrop are admittedly conjectural (Imbrie and others, 1959, Fig. 4).
The elimination of the Arkansas Embayment and the acceptance in its place of the shorelines suggested by Tanner (1959), with a logical northeastern prolongation for same, puts the platy algae facies of Greenwood Shoal close to the presently suggested southeastern shore of the Kansas Sea, instead of across the supposed deeper bottom of the Arkansas Embayment (Fig. 1). As the result of this change in the paleogeography, the southwestern connection with the open ocean remains the only one possible.
Figure 1--Alternative restoration of Kansas Sea at Beattie time with superimposed major channeling (across marine sediments). Central and northern parts motstly after Imbrie and others (1959; see, also Laporte, 1962); southern part modified from Tanner (1959); southeastern part original. Channels modified after Hinds and Greene (1915), and Mudge (1956; also Mudge and Yochelson, 1962). Late Paleozoic ridges of Oklahoma and Texas Panhandle modified from King and others (1942).
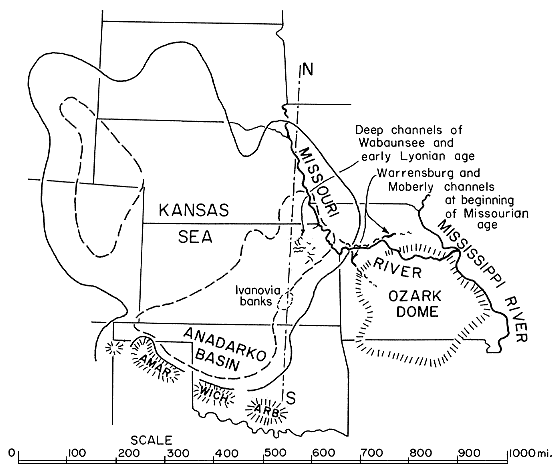
According to a regional tectonic map of the southern Permian Basin in Texas and New Mexico (King and others, 1942), the most probable connection would be across the Dalhart Basin in northwestern corner of the Texas Panhandle, west of the buried Amarillo Mountains (see, Eardley, 1951; Moore and Jewett, 1942); another possible connection would be a saddle between the Amarillo and Wichita Mountain ranges.
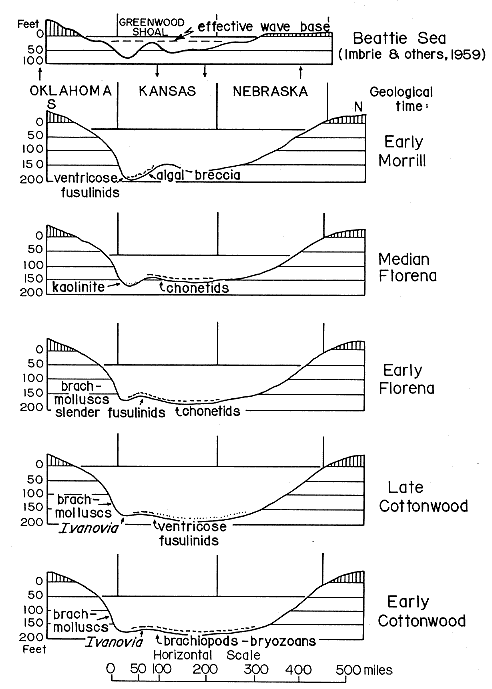
A series of profiles (Fig. 2) to illustrate successive phases of the Beattie oceanic invasion is drawn along the line of the Cottonwood Limestone outcrop, instead of the profile line illustrated by Imbrie, and others, (1959); and Laporte, (1962). The latter profile is shown for comparison, modified to the greater vertical scale used for all profiles.
Figure 2--North-south profile of Kansas Sea at five successive stages of Beattie marine transgression. Distribution of selected biotas and facies modified from Imbrie and others (1959) and Laporte (1962). Profile of "Beattie Sea" (modified to greater vertical exaggeration) after Imbrie and others (1959; see, also Laporte, 1962), is shown for comparison. Arrows indicate postulated uplifts and subsidences.
The chain of basins in the eastern part of the ancient Tethys--Black Sea, Caspian Sea, Aral Sea--provides a modern example of a hydrographic situation that is likely to result in staggering rather than uninterrupted advance of oceanic waters, inasmuch as these basins are isolated at present.
It has been assumed by some that periods of rising oceanic waters (in the course of geologic history) could have been a result of substantial gradual rise of some part of the oceanic bottoms. Such periodic rising of oceanic waters would be a natural cause of periodic marine transgressions over continental shelves and deep into the continents through existing inland seas.
The following stages may be visualized in gradual advance of marine waters through the Tethyan chain of inland basins in a course of a single oceanic transgression.
(1) As water level in the Black Sea began to rise, the waves along its shores would expand their erosional activity farther and farther inland. This activity would be particularly important in the plains north of Caucasian Mountains, where there exists unmistakable evidence of an ancient Black Sea-Caspian Sea connection, in the form of the wide, nearly waterless valley of Manych.
(2) The wave activity of the rising waters eventually would break the land barrier between the Black Sea and Caspian Sea, and the Black Sea waters would start to spill into the Caspian Basin.
The level of Caspian Sea is at the present about 25 meters (75 feet) below oceanic water level, and the divide between the Caspian and Black Seas is 20 to 25 meters high. The total vertical dimension of the volume of water needed to fill the Caspian Basin to the rim of the divide is therefore 40 to 50 meters (135 to 150 feet); and the area to be covered by water is approximately 2 1/2 times the area of the present Caspian Sea. Such was the area of the greater Caspian Sea in its maximal post-Pliocene expansion as shown on the evidence of modern topography by Kovalevsky (1933; Zavoico's review, 1935, Fig. 2, p. 122). The area is equivalent to half the present Mediterranean Sea, and about as large as the Gulf of Mexico.
In view of the great difference of water level in the two adjacent basins, one might expect that the initial rate of flow of water over the broken divide would be as great as, if not greater than, the flow of water over gigantic modern river dams.
(3) However, in the course of time, cumulative loss of water from the ocean to the Caspian Basin would tend to lower the oceanic water level just as would the spreading of the oceanic waters over the newly inundated beaches and low lands elsewhere in the world. Because of all this, the original rate of rise of the oceanic water level is bound to slow down, providing the cause of rise remains the same.
The slackening of the rate of rise of oceanic level would, in its turn, tend to slow down the rush of waters from the expanded Black Sea to the Caspian Basin, and flow could even be temporarily discontinued.
(4) As soon as the water level of the Caspian Sea began to rise rapidly, through mentioned (2) rushing of the waters from Black Sea, the waves along the expanding shores of Caspian Sea would accelerate their erosional activity.
The effect of this activity would be felt particularly in the area southeast of Caspian Sea, the site of an ancient connection between the Caspian and Aral Basins--through the broad east-west valley of Uzboi. This Uzboi area is shown covered by water in Kovalevsky's reconstruction of the greater Aral Sea in post-Pliocene time (Fig. 3; review by Zovoico, 1935, Fig. 2, p. 122).
Figure 3--Apparent staggered oceanic transgression over Caspian Sea-Aral Sea region. A, Map of Mediterranean Sea and seas to east, with superimposed maximal expansion of Caspian Sea and Aral Sea reconstructed by Kovalevsky (1933; reviewed by Zavoico, 1935). B, West-east profile of Azov Sea-Caspian Sea divide, and of northern Caspian Sea. Based mostly on data in Baranov (1954). C, Diagram showing changes in level of Caspian Sea; modified from Kovalevsky (1933; review by Zavoico, 1935). D, Alternative restoration of Kansas Sea at Beattie time, shown for comparison.
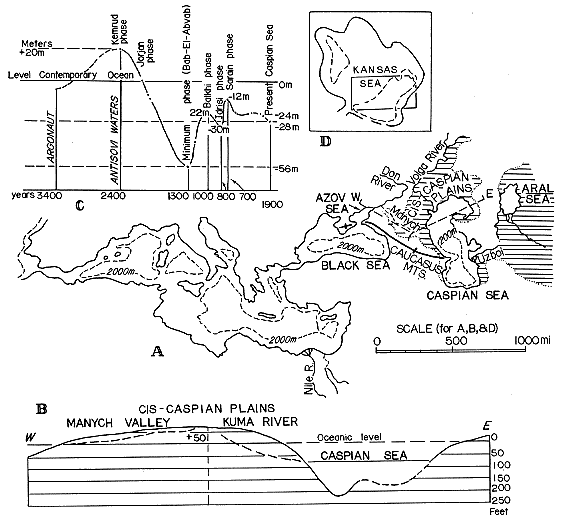
(5) The rush of the Caspian waters through Uzboi Strait into the vast Aral Basin would cause an appreciable lowering of water level in the Caspian Basin, especially if the flow of water from the Black Sea (stage 3) simultaneously slowed down or even became temporarily arrested.
(6) As oceanic water level continued to rise, no matter how much slower than it was originally, waters eventually would resume flowing from the expanded Black Sea into the expanded Caspian, whose water level was temporarily lowered when its waters spilled into the Aral Basin; thus water level of Caspian Sea would resume rising.
The postulated hydrological mechanism may explain the "staggering" nature of the curve, which portrays the latest rise of the Caspian water level in the diagram showing changes in level of Caspian Sea by Kovalevsky (review by Zovoico, 1935, Fig. 3, p. 125) . This staggered rise occurred after a roughly indicated major fall of the Caspian water level from 20 meters above the present oceanic water level (some 2,400 years ago) to 56 meters below the present oceanic level (1,300 years ago Bab-EI-Abvab phase), or total fall of 76 meters (228 feet) in the course of 1,100 years. Next the Caspian level rose to 22 meters below the oceanic level (Balkhi phase), a rise of 34 meters (102 feet) in 300 years; it was followed by the lowering of the level to 30 meters mark (Idrisi phase), or total lowering of 8 meters (24 feet), 200 years later; and then came the second rise of the staggered ascent: to 12 meters (Sarain phase) below the same oceanic level, or total second rise of 18 meters (54 feet). Further changes lowered it gradually down to its present level about 25 meters below the present oceanic level, some 750 years later.
[Note: Kovalevsky estimated the changes in water levels in Caspian Sea on the evidence of the ancient historical records of sea travel, from the Mediterranean to the Black and Caspian Basins. Hence this reference to changing Caspian Sea water level in relation to the present oceanic water level obviously does not imply that the Caspian water level was actually continually below the former oceanic water levels since about 2,000 years ago. The historically recorded influx of the waters from the Black Sea to Caspian Sea over Manych Strait has brought with it some marine Black Sea biota into the Caspian Basin, as indicated by the migration of some marine molluscs mentioned by Kovalevsky.]
The staggered change of water level in the Caspian Sea seems to explain the complexity of megacyclic sedimentation in the Kansas Sea, particularly the typical Shawnee megacyclothems.
The suggested hydrological mechanism, postulated for oceanic transgression through an existing chain of inland basins, provides a basis for the following interpretation of the Shawnee megacyclothems.
The first of the several fusulinid phases in each megacyclothem may be regarded as a culmination of the first cycle in a megacyclic or staggered oceanic invasion, but the first culmination does not necessarily indicate as great a depth of Kansas Sea as was attained in the third or the main cycle of this same megacyclic invasion (Fig. 4).
Figure 4--Generalized columnar sections of two typical megacyclothems of Kansas Sea. A, Beattie megacyclothem, Big Blue Series ("Lower Permian"). Based mostly on data of Imbrie and others (1959). B, Oread megacyclothem, Shawnee Group, Virgilian Series (Upper Pennsylvanian). Based on personal observations, mostly near Lawrence and Lecompton, Kansas, and few places in Nebraska. Curves to left of columnar sections indicate approximate fluctuations in water depth during corresponding marine transgressions.
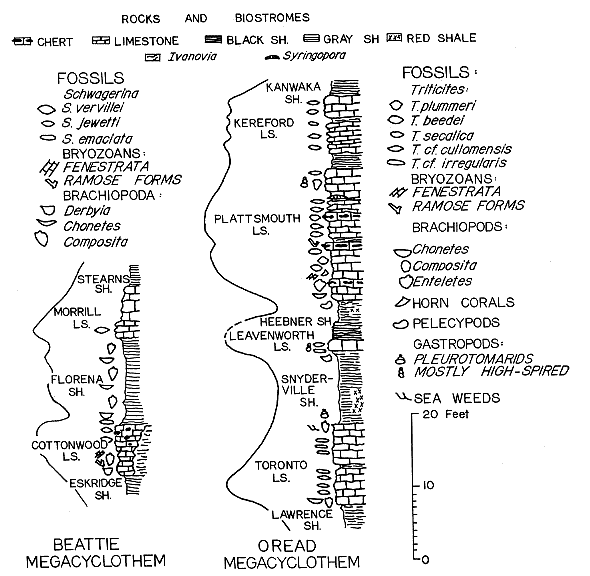
Lower Toronto Limestone--In the most typical, Oread megacyclothem, the deepest waters in the first cycle seem to be indicated by the uppermost 2 feet of the lower, 5 foot bed of the Toronto (Weeping Water) Limestone. The basal 1/2 foot part of this bed is characterized by an abundance of Chonetes granulifer and Composita. It is followed by a 2 1/2 foot layer containing a prolific and varied brachiopod fauna, with scattered fusiform Triticites cf. cullomensis. Next is a 1 1/2 foot layer characterized by great abundance of this same fusulinid accompanied by the rare brachiopods. In the uppermost 1/2 foot there are only a few specimens of T. cf. cullomensis and of the slender T. cf. irregularis, an apparent indication of a slight shallowing of the waters.
Upper Toronto Limestone--One half foot of practically unfossiliferous shale, underlies a 5 1/2 foot upper bed with a basal 1 1/2 foot ledge containing a rich brachiopod fauna, accompanied by a lophophyllid coral and a few molluscs. The basal ledge is overlain by a 2 foot ledge with a great abundance of T. cf. irregularis accompanied by the same invertebrates as in the underlying ledge. The uppermost 2 1/2 feet of the upper bed are usually solid and unfossiliferous below, but it gradually changes upward into a cavernous mudstone with an impoverished biota. It contains poorly preserved molds of a bivalve, some shells of Composita in natural position, and an abundnce of irregularly branching molds, probably of leathery seaweed.
The sequence of the biota in the upper bed of the Toronto may be interpreted to indicate the more or less continually regressing half of the first cycle of the Oread megacycle.
Snyderville Shale--I have found only a few specimens of a poorly preserved pleurotomarid gastropod about 2 feet above the base of the succeeding 10 foot thick Snyderville Shale. In the calcareous uppermost 1/2 foot of the Snyderville, there is a good "mixed" fauna of brachiopods, including abundant Chonetes, and various pelecypods.
Leavenworth Limestone--The overlying Leavenworth Limestone (1 1/2 feet thick) contains a moderate number of smaller brachiopods, such as Marginifera sp., Crurithyris planoconvexa, and rare Chonetes, as well as a few pelecypods and several genera of small gastropods. Fusulinids, mostly scattered T. cf. cullomensis, and rarely T. cf. irregularis, are not rare in the upper part; but in Nebraska west of St. Joseph, the latter species is abundant in the basal part of the limestone. The Leavenworth may be considered to indicate a second, comparatively brief cycle of the staggering Oread oceanic invasion, resulting in maximal depth of waters hardly exceeding the depth at the culmination of the first (Toronto) cycle.
Heebner Shale--The occurrence of black fissile shale in the lower part of the Heebner Shale, is interpreted to indicate a suppression of the abundant biota of the Leavenworth cycle, as a result of pollution by toxic black mud. It seems that this sudden appearance of the black shale (or merely of carbonaceous material, which takes the place of black-fissile shale in the middle of some megacyclothems) is indicative of an advance of oceanic waters into some previously isolated basin or less accessible area where large supplies of black mud had accumulated (see, Zangerl and Richardson, 1963). In the calcareous, uppermost 1/2 foot thick part of the Heebner Shale, there are some pelecypods, Crurithyris planoconvexa, and Orbiculoidea.
Plattsmouth Limestone--ln the exposures about 1 mile south of Plattsmouth, at the base of the Plattsmouth Limestone is a layer filled with gregarious Chonetes. Here, I have observed the previously mentioned peculiar valve-in-valve arrangement of the three chonetid valves. The next 4 to 4 1/2 feet of the Plattsmouth Limestone contain abundant fauna of small- and medium-sized brachiopods including Enteletes plattsmouthensis, and here occurs also a fairly large horn coral.
About 5 feet above the base of the Plattsmouth occur its most interesting fusulinids: the subrhombic Triticites beedei and the subspherical T. plummeri, accompanied by the abundant Rhipidomella pecosi and few other small brachiopods. At some localities these same fossils are scattered in a zone up to 4 feet thick but never become gregarious. On the contrary, the fusiform Triticites secalicus is invariably gregarious in the many zones of the upper 10 feet of the Plattsmouth Limestone. These fusulinid layers are interstratified with layers in which calcareous brachiopods predominate. Small gastropods become locally abundant in the uppermost 3 feet of the Plattsmouth Limestone, where Composita is the only common brachiopod. Fusulinids are scarce, and they seem to be smaller than typical Triticites secalicus.
Kereford Limestone--This same, smaller than normal T. cf. secalicus appears also in the Kereford Limestone, which is separated from the Plattsmouth Limestone by an apparently unfossiliferous shale. The fauna of the uppermost Plattsmouth and the Kereford seems to indicate shallowing of the Oread Sea, after it had attained its maximum depth (probably about 200 feet) apparently in medial and late Plattsmouth time.
Zangerl and Richardson (1963) investigated an area where black muds are accumulating in the modern Mississippi Delta as representing a similar environment under which the Pennsylvanian black shales of Illinois accumulated (p. 8):
The over-all character of the . . . [black] shales suggested a rather unusual environment of deposition. modern situations where black muds (consisting largely of particulate plant decomposition products) are forming occur here and there along the Gulf Coastal Plain of North America (and no doubt elsewhere). There are small ponds, lakes and bayous amid cypress swamps, often covered by a floating mat of vegetation (flotant).
Similar conditions are suggested for the development of the black shale in the early deposits of the Pennsylvanian in Illinois, Indiana, and Kentucky basin, where deltaic sediments have been widely deposited (Potter and Glass, 1958). The intricate system of shallow sandstone-filled channels in this basin is different from the above discussed, much deeper channels, in the eastern slopes of Kansas Sea; and extremely shallow hydrography suggested by Imbrie and associates for it in Beattie time, would fit much better the Illinois-Indiana-Kentucky region of marine invasions, represented by the local Pennsylvanian sandstones and black shales. On the other hand, the historical changes of the water level in the Caspian Sea are of a magnitude comparable to that suggested by me for the depth of deposition of the Big Blue (Lyonian) sediments in Kansas. The disappearance of the black shale in the cyclothems of the Wabaunsee and later marine deposits appears to indicate that post-Shawnee transgressions were no longer capable to reach the source area of the black muds.
Impartially speaking, it appears that we have now two opposing hypotheses regarding a maximum depth of the late Paleozoic Kansas Sea: one suggests its maximum depth of not more than 80 feet for the Beattie, and by implication for other Lyonian (Big Blue) marine transgressions (Imbrie and others, 1959). The other, earlier hypothesis, allows as much as 180 feet for these same transgressions, based on a belief that the fusulinid phase of cyclic sedimentation, wherever developed, indicates culmination of the transgression (Elias, 1937). In contrast, the first mentioned hypothesis considers the fusulinid facies to indicate nearshore turbulent waters, less than 30 feet deep.
It appears, furthermore, that the strongest argument in favor of the shallower water conditions is the supposition that the abundance of organisms believed to be blue-green and green algae, indicate their development in the photic zone, whose lower limit is set at some 80 to 90 feet below the water surface. Corroboration is seen in the evidence of agitated water, such as broken organic skeletons and distribution of terrigenous sedimentary material.
The deeper water hypothesis appears to have its greatest support in some geological evidence, such as comparative hydrology of the modern epicontinental seas; and particularly in the direct evidence of the maximal depth of the channels as much as 180 feet, that cut through the marine sediments of the Kansas Sea.
Even if the general interior relief of the North American continent in late Paleozoic time was so subdued that almost a "wading depth" of the Kansas Sea, in spite of all its rich biota, can be postulated, the observed deep channeling requires special explanation. The seemingly strongest argument in favor of the shallower water hypothesis, is the postulated lower limit of the photic zone commonly supposed to control the development of abundant algae. However, certain green and brown, and particularly red algae, are known to occur abundantly to the depth of about 175 feet in the bay of Naples. Furthermore, there is no certainty that the filamentous forms classified as Girvanella, if truly algae, belong to blue-green or green algae; nor is it known whether the platy Ivanovia is definitely a green, brown, or red alga.
The lower boundary of the supposed near-shore turbulence of water depends to a considerable extent on the postulated maximal depth of a basin, the shallower the basin, the smaller the amplitude of the waves, and the less the depth of disturbance. In any event, it is well known that hurricanes agitate water to much greater depths than do normal waves; and it seems hardly reasonable to assume that there were no hurricanes in late Paleozoic time.
The problem of the history of the Kansas Sea is far from being simple and its fully satisfactory solution undoubtedly requires all embracing additional research. Every facet of the problem needs to be re-explored and re-evaluated. The paleoecological approach should be properly meshed with the "cycleological." It is odd for the present state of the paleoecological doctrine that cyclic and rhythmic sedimentations are left out of consideration (Gekker, 1957; Ager, 1963). Perhaps the great American late Paleozoic Mediterranean is destined to become a classical region where the two equally important research approaches, the paleoecological and cycleological will be integrated.
Ager, D. Y., 1963, Principles of paleoecology: McGraw-Hill Book Co., New York, 371 p.
Bandy, Orville, 1960, General correlation of foraminiferal structure with environment: 21st. Internat. Geol. Cong. Rept., Norden, pt. 22, p. 7-19.
Baranov, A. N. (editor), 1954, Atlas Mira, Part 1, Moscow.
Black, Maurice, 1933, The algal sediments of Andros Island, Bahamas: Phil. Trans., Royal Soc. London, B, v. 22, p. 165-192.
Cloud, P. E., 1942, Notes on stromatolites: Am. Jour. Sci., v. 240, no. 5, p. 363-379.
Dunbar, C. O., 1957, Fusuline Foraminifera-annotated bibliography, in Treatise on Marine Ecology and Paleoecology, V. 2, Paleoecology: Geol. Soc. America Mem. 67, p. 753-754.
Eardley, A. J., 1951, Structural geology of North America: Harper & Bros., New York, 624 p.
Elias, M. K., 1937, Depth of deposition of the Big Blue (late Paleozoic) sediments in Kansas: Geol. Soc. America Bull., v. 48, p. 403-432.
Elias, M. K., 1959, Review of "Facies of Upper Carboniferous and Artinskian deposits in the SterlitamakIshimbaevo region. . . .": Internat. Geol. Review, v. 1, no. 2, p. 39-88.
Elias, M. K., 1962, Comments on recent paleoecological studies of Late Paleozoic rocks in Kansas: Kansas Geol. Soc. 27th Field Conf. Guidebook, p. 106-115.
Gekker (Hekker), R. F., 1957, Yvedenie v paleoekologiyu, Nauch.-Tekh, Izd. Lit. Geol. i, Izdat, Akad. Nauk SSSR. Okhrane Nedr, Moscow. (French translation by J. Roger: Hekker, R. F., Basses de la Paleoecologic, 1960, Paris, Bur. Recherches Geol. Min.).
Gubler, J., 1935, Les fusulinides du Permien de l'Indochine: Soc. Geol. de France, Mem. Nou. Ser., t. 11, fasc. 4, Mem. 26, 69 p.
Ginsburg, R. N., and others, 1954, Laminated algal sediments of south Florida and their recognition in the fossil record: Unpub. rept., Marine Lab., Miami Univ.
Ginsburg, R. N., and Lowenstam, H., 1958, The influence of marine bottom communities on the depositional environments of sediments: Jour. Geology, v. 66, p. 310-318.
Henbest, L. G., 1963, Biology, mineralogy, and diagenesis of some typical Late Paleozoic sedimentary Formaminifera and algal-foraminiferal colonies: Cushman Found. Foram. Research, Spec. Publ. no. 6, 441 p.
Hinds, H., and Greene, F. C., 1915, The stratigraphy of the Pennsylvanian Series in Missouri: Missouri Bureau Geol. and Mines, v. 13, 2nd ser., 407 p.
Illing, L. Y., 1954, Bahaman calcareous sands: Am. Assoc. Petroleum Geologists Bull., v. 38, p. 1-95.
Imbrie, John, 1955, Quantitative lithofacies and biofacies study of Florena Shale (Permian) of Kansas: Am. Assoc. Petroleum Geologists Bull., v. 39, p. 649-670.
Imbrie, John, Laporte, L. F., and Merriam, D. F., 1959, Beattie Limestone facies and their bearing on cyclical sedimentation theory: Kansas Geol. Soc. 24th Field Conf. Guidebook, p. 69-78.
Ivanova, E. A., and Khvorova, I. Y., 1955, Stratigrafiya srednego i verkhnego karbona zapadnoi chasti Moskovskoi sineklizy (Razvitie fauny sredne- i verkhnekamennougolnogo morya zapadnoi chasti Moskovskoi sineklizy v svyazi s ego istoriei, kniga 1): Akad. Nauk SSSR, Paleont. lnst., Tr. t. 53, 282 p.
Ivanova, E. A., 1958, Razvitie fauny v svyazi s usloviyami sushchestvovaniya: Akad. Nauk. SSSR. Paleont. lnst., Tr., t. 69, 303 p.
Johnson, J. H., 1946, Lime-secreting algae from the Pennsylvanian and Permian of Kansas: Geol. Soc. America Bull., v. 57, p. 1087-1120.
Kahler, F. I., and Kahler, G., 1947, Die Pseudoschwagerinen der Granzlandbanke und des Oberen Schwagerinakalkes: Palaeontographica, Bd. 87, pt. A, 43 p.
Khvorova, I. Y., 1946, On a new genus of algae from the middle Carboniferous deposits of the Moscow Basin: Acad. Sci. URSS, C. R. (Doklady) , v. 53, no. 8, p. 737-739.
King, R. E., and others, 1942, Resume of the geology of the south Permian Basin, Texas and New Mexico: Geol. Soc. America Bull., v. 53, p. 539-560.
Konishi, K., and Wray, J., 1961, Eugonophyllum, a new Pennsylvanian and Permian algal genus: Jour. Paleontology, v. 35, p. 659-665.
Kovalevsky, S. A., 1933, Lik Kaspiya: Azneft, Baku, SSSR, 129 p.
Lane, N. G., 1958, Environment of deposition of the Grenola Limestone (Lower Permian} in southern Kansas: Kansas Geol. Survey Bull. 130, pt. 3, p. 117-164.
Laporte, L. F., 1962, Paleoecology of the Cottonwood Limestone (Permian), northern Mid-continent: Geol. Soc. America Bull., v. 73, p. 521-524.
Moore, R. C., 1950, Late Paleozoic cyclic sedimentation in central United States: 18th Internat. Geol. Cong., Great Britain, 1948, Rept., pt. 4, p. 5-16.
Moore, R. C., and Jewett, J. M., 1942, Oil and gas fields of Kansas: The Mines Magazine, October, 1942, p. 1-16.
Mudge, M. R., 1956, Sandstones and channels of the Upper Pennsylvanian and Lower Permian in Kansas: Am. Assoc. Petroleum Geologists Bull., v. 44, no. 4, p. 654-678.
Mudge, M. R., and Yochelson, E. L., 1962, Stratigraphy and paleontology of the uppermost Pennsylvanian and lowermost Permian rocks in Kansas: U. S. Geol. Survey Prof. Paper 323, 213 p.
Potter, P. E., and Glass, H. D., 1958, Petrology and sedimentation of the Pennsylvanian sediments in southern Illinois: a vertical profile: Illinois Geol. Survey, Rept. Inv. 204, 601 p.
Rauser-Chernousova, D. M., 1950, Fatsii Verkhnekamennougolnykh i Artinskikh otlozhenii Sterlitamaksko-Ishimbaiskogo Priuralya: Akad. Nauk SSSR, Inst. Geol. Nauk, Tr., t. 119, Geol. Ser., no. 43, 108 p.
Reitlinger, S. A., 1950, Foraminifery Srednekamennougolnykh otlozhenii tsentralnoi chasti Russkoi Platformy: Akad. Nauk SSSR, Inst. Geol. Nauk, Tr., t. 126, Geol. Ser. no. 47, 111 p.
Ross, C. A., 1963, Standard Wolfcampian Series (Permian), Glass Mountains, Texas: Geol. Soc. America, Mem. 88, 205 p.
Tanner, W. J., 1959, Permo-Pennsylvanian paleogeography of part of Oklahoma: Jour. Sed. Pet., v. 29, p. 326-335.
Thompson, M. L., 1948, Studies of American fusulinids: Univ. Kansas, Paleont. Contr., Protozoa, Art. 1, 184 p. [available online]
Thompson, M. L., 1951, New genera of fusulinid Foraminifera: Cushman Found. Foram. Research, Contr., v. 2, p. 115-119.
Thompson, M. L.. 1954, American Wolfcampian fusulinids: Univ. Kansas, Paleont. Contr., Protozoa, Art. 5, 225 p. [available online]
Twenhofel, W. H., 1919, Precambrian algae and Carboniferous algal deposits: Am. Jour. Sci., 4th Ser., v. 38, p. 339.352.
Wood, Alan, 1963, The British Carboniferous species of Girvanella (calcareous algae): Paleontology, v. 6, pt. 2, p. 264.273.
Zavoico, B. B., 1935, Review of "The Face of the Caspian Sea": Am. Assoc. Petroleum Geologists Bull., v. 19, p. 120-125.
Zangerl, Rainer, and Richardson, E. S., Jr., 1963, The paleoecological history of two Pennsylvanian black shales: Chicago Natural History Museum, Fieldiana, Geology Memoires, v. 4, 353 p.