![]()
Prev Page--Fossil vertebrates and mollusks || Next Page--Stratigraphy
Paleontology, continued
Fossil Plants
For stratigraphic correlations fossil plants are the most useful of the several kinds of organic remains preserved in Ogallala rocks. Hulls of grass seeds and fruits of various forbs are numerously represented in Ogallala sediments in the region here considered; these plant remains are generally well preserved and can be collected, properly preserved, and stored without expensive equipment, and without excessive expenditure of time and effort; diagnostic characters are generally so well shown that accurate determinations can be made; vertical ranges of most species are limited and generally well known; and finally, seeds are so numerous, both vertically in the succession of Ogallala rocks and geographically over the area of Kansas covered by Ogallala deposition, that these plant fossils assume primary importance as a tool for local as well as for regional stratigraphic correlations. It is rare indeed that an exposure of Ogallala rocks does not reveal at least one kind of fossil seed, and more frequently than not, general stratigraphic placement of even isolated small exposures can be made on the evidence presented by fossil seeds.
Those interested in stratigraphic problems of late Cenozoic deposits in the plains region of Kansas and Oklahoma are indebted to Dr. Maxim K. Elias, whose notable studies of fossil seeds (Elias, 1942) provided stratigraphers with a new and highly usable tool for correlation studies in this segment of the rock column. We wish to express our gratitude to Dr. Elias, who has provided us with much valuable information, and who generously confirmed the identifications of many of our seed collections.
Without exception, so far as known, seeds in the Ogallala formation of northwestern Kansas are opalized. Seeds of Biorbia fossilia were analyzed spectrochemically by John Schleicher of the Geochemical Laboratories of the Kansas Geological Survey; his analysis is given below:
"The fossil seeds from the Ogallala are mostly silica . . . However, there is some calcium and aluminum present . . . about 2 to 3 percent calcium and the same amount for aluminum, both calculated as the metal. There is about 0.5 percent iron, 0.2 percent magnesium and a trace of copper. The remainder is silica."
The fact that the predominant mineral present in the fossilized seeds is opal makes it feasible to use dilute hydrochloric acid to remove seeds from sediments cemented by calcite.
Fossil seeds in Ogallala elastics can be found merely by examining surfaces of rocks that have been exposed to weathering, although close attention to detail is necessary because seeds may be discolored or even covered by growths of moss, lichens, or algae. Seeds of many species are small, being no more than 2 mm in diameter, and some experience is required to find them, even where they may be numerous. Strangely, fossil seeds, including the seemingly delicate hulls of grass seeds, are found in all varieties of sediments, and they occur in large numbers in some coarse angular sands or gravels. Obviously, they could not.have been transported any significant distance with these clastics, a fact that increases their value for stratigraphic purposes.
Seeds commonly occur in local dense aggregations of twenty to several hundred individuals in rocks where single seeds are rare. Not infrequently, such a local aggregation is the only collection from an exposure, and more than once we have carefully searched a suitable exposure and decided it was barren of fossil seeds, only to return later and find a rich collection at some small, previously overlooked spot on the outcrop. The reason for this unusual distribution within the rocks is not understood, but Elias has suggested that some of these aggregations result from passage through animals, that is, they are the remains of fecal deposits; others may be the result of minor eddy currents in sluggish water. Such erratic distribution is not universal; in many beds seeds may be found almost uniformly dispersed. It is not usual to find seeds uniformly distributed vertically through great thicknesses of section; generally speaking, worth-while collections can be made only from certain strata, others being almost or totally devoid of fossil seeds. There seems to be no correlation between the nature of the sediments and the numbers of contained fossil seeds; at one locality seeds may be most abundant in fine sand, at another in coarse sands and gravels, or seeds may occur in both types of rock at the same locality. It is our judgment that this phenomenon reflects primarily the nature of Ogallala deposition in a complex series of channel-ways, so that at the same time seeds might become entombed in fine sands and silts, or in coarse sands and gravels, depending upon local conditions in the depositing channel.
Grass hulls commonly occur in loose sand, in an elongate pattern of distribution, and mixed with various kinds of organic debris, suggesting drift along the border of a stream. Seeds commonly occur in sediments that have undergone some degree of induration, and from these rocks the seeds must be removed by knife or awl, or pieces of the rock may be taken to the laboratory and the seeds removed, or at least loosened from the rock, by treatment with dilute hydrochloric acid. At least part of the cementing mineral that binds the "mortar beds" in the Ogallala is acid-soluble. Calcite is not the predominant cementing substance everywhere, however; at many localities, especially in the northern tier of counties in Kansas, the predominant cementing mineral is not acid-soluble and is presumed to be opal or fine-grained chert. Hulls of Stipidium illustrated in Plate 5A occur in medium to coarse indurated sands in which only a small part of the cementing material is acid-soluble, so of course removal of the fossils from the matrix is difficult. The source of the silica that forms the cementing substance is not known; it could come from weathering of feldspathic sands in the rocks themselves, or from thin, widespread volcanic ash falls that have subsequently disappeared through weathering. The latter hypothesis seems more tenable, as the sands show few signs of weathering whereas volcanic ash lentils are common, and bentonitic clays, although not conspicuous, are widespread.
The fossil seeds from the Ogallala formation (Pl. 8, 9; Fig. 4) may be divided into three major, stratigraphically significant groups: (1) seeds that occur in the Valentine member, represented in Kansas by a single species of grass, Stipidium commune; (2) a large assortment of grasses and other herbs characteristic of the Ash Hollow member, among which Biorbia fossilia is most diagnostic, most widespread, most numerous, and usually most easily found; and (3) a small group diagnostic of the Kimball member, including Prolithospermum johnsoni and Berrichloa minima. In addition, floral subzones permit a rough subdivision of sediments in the Ash Hollow into upper, middle, and lower zones (Fig. 5).
The fruits of the hackberry tree, Celtis willistoni, are widespread and in many places abundant, but are found at all stratigraphic levels of the Ogallala formation, hence are of no stratigraphic significance.
Plate 8--Fossil seeds from Ogallala formation. a. Stipidium commune Elias; b. Stipidium elongatum Elias; c. Stipidium variegatum Elias; d. Stipidium grande Elias; e. Stipidium nebraskense Elias; f. Stipidium aristatum (Berry) Elias; g. Panicum elegans Elias; h. Prolithospermum johnsoni Elias. All figures X 9 [images enlarged for web viewing].
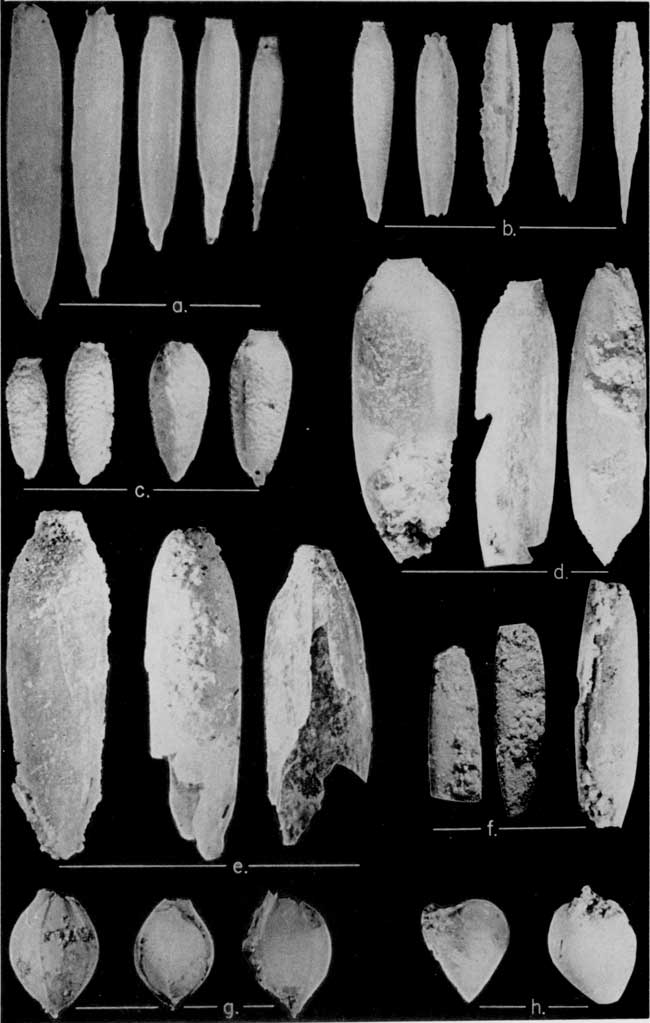
Plate 9--Fossil seeds from Ogallala formation. i. Berrichloa maxima Elias; j. Berrichloa tuberculata Elias; k. Berrichloa conica Elias; l. Berrichloa amphoralis Elias; m. Krynitzkia auriculata Elias; n. Krynitzkia coroniformis Elias; o. Biorbia fossilia (Berry) Elias; p. Celtis willistoni (Cockerell) Berry. All figures X 9 [images enlarged for web viewing].

Figure 4--Check list of fossil seeds. Occurrences in Valentine, Ash Hollow, and Kimball members are indicated by symbols V, AH, and K.
| Plant Fossil Species |
NE NE sec. 3,T. 12 S., R. 42 W., Wallace Co. | SE sec. 11, T. 13 S., R. 42 W., Wallace Co. | C W line NW sec. 35, T. 14 S., R. 42 W., Wallace Co. | NW SE sec. 16, T. 14 S., R. 40 W., Wallace Co. | W2 SW sec. 7, T. 14 S, R. 38 W., Wallace Co. | SE SE sec. 20, T. 5 S., R. 36 W., Rawlins Co. | SE SE sec.. 13, T. 3 S., R. 35 W., Rawlins Co. | C W line SW sec. 4, T. 4 S., R. 34 W., Rawlins Co. | SW SE sec. 2, T. 3 S., R. 33 W., Rawlins Co. | SW NW sec. 24, T. 3 S, R. 30 W., Decatur Co. | SE SW sec. 8, T. 4 S., R. 29 W., Decatur Co. | NW NE sec. 34, T. 1 S., R. 27 W., Decatur Co. | C N line sec. 36, T. 2 S., R. 25 W., Norton Co. | C E line sec. 34, T. 5 S., R. 25 W., Norton Co. (New Almelo section) | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | |
| Prolithospermum johnsoni | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Berrichloa maxima | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Panicum elegans | • | |||||||||||||||||||||||||||||||||||||||||
| Berrichloa glabra | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Berrichloa conica | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Stipidium tubus | • | • | ||||||||||||||||||||||||||||||||||||||||
| Celtis willistoni | • | • | • | • | • | • | • | • | • | • | • | • | • | |||||||||||||||||||||||||||||
| Stipidium elongatum | • | • | • | • | ||||||||||||||||||||||||||||||||||||||
| Stipidium grande | • | |||||||||||||||||||||||||||||||||||||||||
| Biorbia fossilia | • | • | • | • | • | • | • | • | • | • | • | |||||||||||||||||||||||||||||||
| Stipidium aristatum | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Stipidium nebraskense | • | • | ||||||||||||||||||||||||||||||||||||||||
| Stipidium kansasense | ||||||||||||||||||||||||||||||||||||||||||
| Stipidium variegatum | • | • | • | • | • | |||||||||||||||||||||||||||||||||||||
| Berrichloa amphoralis | • | • | • | • | • | • | • | |||||||||||||||||||||||||||||||||||
| Berrichloa tuberculata | • | |||||||||||||||||||||||||||||||||||||||||
| Krynitzkia auriculata | • | • | • | |||||||||||||||||||||||||||||||||||||||
| Krynitzkia coroniformis | • | |||||||||||||||||||||||||||||||||||||||||
| Stipidium commune | ||||||||||||||||||||||||||||||||||||||||||
| Total number of species |
4 | 6 | 3 | 2 | 6 | 3 | 3 | 3 | 7 | 5 | 4 | 3 | 9 | 5 | 3 | |||||||||||||||||||||||||||
| Plant Fossil Species |
NW NW sec. 2, T. 4 S., R. 24 W., Norton Co. | sec. 35, T. 5 S., R. 24 W., Norton Co. (Lenora section) | C W line sec. 29, T. 2 S., R. 22 W., Norton Co. (Calvert section) | W2 sec. 30, T. 5 S., R. 22 W., Norton Co. (Edmond section) | W2 sec. 16, T. 2 S., R. 21 W., Norton Co. (Almena section) | sec. 17, T. 5 S., R. 20 W., Phillips Co. | NW NE sec. 30, T. 1 S., R. 19 W., Phillips Co. | SW SW sec. 31 T. 1 S., R. 19 W., Phillips Co. | C W line SW sec. 32, T. 1 S., R. 14 W., Smith Co. | SE SE sec. 2, T. 16 S., R. 33 W., Scott Co. (Scott County State Lake section) | NE NE sec. 23, T. 12 S., R. 32 W., Logan Co. | SE sec. 16, T. 11 S., R. 25 W., Trego Co. | C N line SW sec. 7, T. 8 S., R. 26 W., Sheridan Co. | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | V | AH | K | |
| Prolithospermum johnsoni | |||||||||||||||||||||||||||||||||||||||
| Berrichloa maxima | |||||||||||||||||||||||||||||||||||||||
| Panicum elegans | • | ||||||||||||||||||||||||||||||||||||||
| Berrichloa glabra | • | • | |||||||||||||||||||||||||||||||||||||
| Berrichloa conica | • | • | • | • | |||||||||||||||||||||||||||||||||||
| Stipidium tubus | • | ||||||||||||||||||||||||||||||||||||||
| Celtis willistoni | • | • | • | • | • | • | • | • | • | • | • | • | |||||||||||||||||||||||||||
| Stipidium elongatum | • | • | |||||||||||||||||||||||||||||||||||||
| Stipidium grande | • | ||||||||||||||||||||||||||||||||||||||
| Biorbia fossilia | • | • | • | • | • | • | • | • | • | • | • | • | |||||||||||||||||||||||||||
| Stipidium aristatum | • | ||||||||||||||||||||||||||||||||||||||
| Stipidium nebraskense | |||||||||||||||||||||||||||||||||||||||
| Stipidium kansasense | |||||||||||||||||||||||||||||||||||||||
| Stipidium variegatum | • | • | • | • | • | ||||||||||||||||||||||||||||||||||
| Berrichloa amphoralis | • | • | • | • | • | • | |||||||||||||||||||||||||||||||||
| Berrichloa tuberculata | • | • | • | • | • | ||||||||||||||||||||||||||||||||||
| Krynitzkia auriculata | |||||||||||||||||||||||||||||||||||||||
| Krynitzkia coroniformis | • | • | • | • | • | ||||||||||||||||||||||||||||||||||
| Stipidium commune | • | • | • | • | |||||||||||||||||||||||||||||||||||
| Total number of species |
3 | 4 | 1 | 6 | 1 | 5 | 1 | 7 | 1 | 5 | 5 | 7 | 3 | 2 | 5 | 5 | |||||||||||||||||||||||
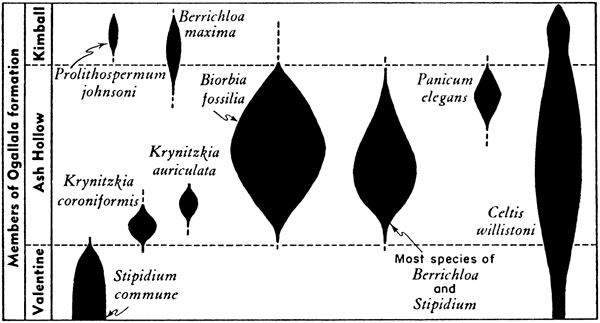
Figure 5--Diagrammatic representation of stratigraphic range of fossil plants in the Ogallala formation of Kansas.
Seeds from the Valentine member
The single diagnostic fossil seed found within sediments of the Valentine member is that of the grass Stipidium commune, related to the Stipa grasses now growing in northwestern Kansas and elsewhere on the plains. According to Elias (1942, p. 80) Stipidium commune ". . . extends also into the succeeding Krynitzkia coroniformis zone of the Ash Hollow formation [member of Kansas classification,] but in higher horizons is replaced by S. kansasense and other larger species of Stipidium." We have found S. commune in northwestern Kansas in a few localities (Fig. 4), but never in direct association with K. coroniformis or other species of Stipidium, and it is our judgment that in this region S. commune may be taken to be diagnostic of the Valentine member. The species undoubtedly ranges through great thicknesses of Valentine beds, but in northwestern Kansas only the upper part of the Valentine member is exposed, and that at relatively few localities.
Stipidium commune exists in the Ogallala formation as the opalized hulls of the seeds of this grass, but fortunately the characters on the hull (palea and lemma) are those most valuable for taxonomic classification. Hulls of S. commune are illustrated on Plate 8a. The length of the hull ranges from 3 to 8 mm, the diameter from 0.7 to 1.3 mm. The smallest individuals are densely tuberculate over their entire length, but most large individuals are smooth over their posterior one-third to one-half. The slender form, general size, and finely tuberculate sculpture are the characters most usable for recognition of this fossil plant. S. commune differs from S. dawsense from the Miocene Sheep Creek formation by its larger size and general tuberculation; S. kansasense, which succeeds S. commune in the lower part of the Ash Hollow member, is larger and more coarsely tuberculate.
Seeds from the Ash Hollow member
Fifteen species of fossil seeds are here reported from Ash Hollow deposits in northwestern Kansas; most of these are illustrated on Plates 8 and 9, and the known distribution at 27 stratigraphically significant localities is shown on Figure 4; lesser collections from other localities judged to have minor stratigraphic significance are not reported here. Elias (1942) has reported other fossil seeds from the Ash Hollow member in Nebraska, so that the entire flora is somewhat larger than that reported by us.
By far the most widely dispersed, most numerous, and most characteristic fossil seed in the Ash Hollow member is Biorbia fossilia; we have treated it (as did Elias) as a fossil diagnostic of the Ash Hollow member.
Biorbia fossilia is a borrage, represented by small ovoid nutlets; these range from 2 to 3 mm in length and 1.75 to 2.5 mm in diameter (Pl. 9o). The nutlet is asymmetrically compressed, the dorsal side being more convex than the ventral. There is a keel over the cuspidate apex or tip of the seed to the scar of attachment, where the keel assumes a lateral position to one side of the scar. The scar of attachment is round and elevated, consisting of two raised rings, one inside the other. The rims, especially that of the inner ring, are dentate or slightly fluted. The surface of most seeds is rugose, the sculpture being formed of an irregular pattern of winding elevated ridges, which coalesce here and there to form a sort of honeycombed or reticulate pattern. Many seeds, presumably of the same species, are quite smooth and some of these have a polished appearance. The ratio of rugose to smooth forms differs widely from place to place. At some exposures of Ash Hollow rocks only rugose forms are found, even in large collections comprising several hundred individuals; at other localities, not obviously different lithologically or otherwise, a few smooth seeds may be found interspersed among rugose seeds to the extent of ten percent or even more. At a few localities smooth seeds have been found exclusively. Whether this situation represents the occurrence of a mutation that produced smooth seeds in various proportions according to the genetic constitution of the local population, or whether the smooth seeds represent another species of Biorbia has not been determined.
Elias (1942, pl. 17) divided the Ash Hollow member into three floral subzones; we have found these subzones generally valid and have utilized local floras of fossil seeds in conjunction with other types of data, including assemblages of fossil mollusks and the distinctive properties of volcanic ash falls, to determine the approximate placement of sediments in the lower, middle, or upper part of the Ash Hollow member.
Characteristic seeds from the lower third of the Ash Hollow member, which Elias (1942, pl. 17) referred to as the Krynitzkia coroniformis subzone, include the tiny nutlets of K. coroniformis (Pl. 9n), Stipidium kansasense, and K. auriculata; others less characteristic of the lower strata of the Ash Hollow member, include Berrichloa conica, B. amphoralis, Stipidium variegatum, and S. elongatum. Stipidium aristatum probably belongs at the top of the lower zone of the Ash Hollow member. There is no question of Krynitzkia coroniformis and K. auriculatum being diagnostic of the lower third of the Ash Hollow; other species associated with these, including Biorbia fossilia, which is not numerous at this level, are not diagnostic. S. kansasense probably is nearly as diagnostic of this lower seed zone as the two species of Krynitzkia, but its usefulness is less because it is less widely dispersed.
The middle third of the Ash Hollow member is characterized by large populations of Biorbia fossilia, which reach their peak here after feeble beginnings in the lower Ash Hollow; populations of B. fossilia decline above the middle of the Ash Hollow member, and are virtually absent in the Kimball member. Other seeds associated with B. fossilia in middle Ash Hollow rocks include Stipidium grande, S. nebraskense, S. elongatum, S. intermedium, Berrichloa conica, B. tuberculata, and locally B. amphoralis. These associated seeds are typically present in middle Ash Hollow rocks, but cannot be said to be diagnostic of this portion of the member, because all occur less commonly at other stratigraphic positions within the member. An additional characteristic of the floral assemblages present in the middle portion of the Ash Hollow member is the complete absence here of Krynitzkia coroniformis and K. auriculata, which are restricted to the lower part of the Ash Hollow, and the absence or rare occurrence of Panicum elegans and Stipidium tubus, which are characteristic of the upper third of the Ash Hollow.
The upper part of the Ash Hollow member is characterized by Panicum elegans, Stipidium tubus, and Berrichloa maxima (Panicum elegans subzone of Elias), although it is our observation that B. maxima is almost restricted to sediments of the Kimball member.
Seeds from the Kimball member
The fossil seed flora in deposits of the Kimball member is sparse; we have found Prolithospermum johnsoni to be restricted to this member, although it is not numerous in it.
The nutlets of Prolithospermum johnsoni (Pl. 8h) superficially resemble in size and shape those of the smooth variety of Biorbia fossilia, but differ from them in several important details. The ventral keel of the nutlet of P. johnsoni extends over the cuspidate apex as does that of Biorbia fossilia, but whereas the keel on the seed of B. fossilia extends completely to the scar of attachment, that on the seed of P. johnsoni dwindles in size as it approaches the scar of attachment, and disappears well in front of the scar. The scar of attachment on the seed of P. johnsoni is relatively much larger than that on the seed of B. fossilia, the rim rises higher, and the scar is buttressed anteriorly by a swelling or ridge that extends toward the apex. This ridge is lacking on the nutlets of B. fossilia. P. johnsoni can be distinguished easily from the rugose nutlets of B. fossilia, but can be confused with the smooth variety of the seeds of B. fossilia unless strict attention is paid to the details mentioned.
Berrichloa maxima, which is known to occur also in the uppermost part of the Ash Hollow member, is a common associate of P. johnsoni. Elias (1942, p. 97) reports Berrichloa minuta in association with P. johnsoni and Berrichloa maxima at a locality 4 miles south of Sharon Springs, Wallace County. We failed to find B. minuta at these exposures; the "small variety of Biorbia" that Elias mentions as another associate of P. johnsoni at this locality is likewise missing from our collections.
The sparseness of the flora in deposits of the Kimball member may be due in part to local unfavorable ecological conditions prevalent at the time of deposition of these sediments, but we judge that another, equally tenable explanation may be found. The heavy accumulations of caliche, common in the upper portions of the Kimball member, may render the seeds more difficult to find, or the seeds may have been destroyed in the processes that produced the accumulations of caliche, or both.
In summary, it must be emphasized that for the field stratigrapher interested in local and regional correlations, fossil seeds are the most useful stratigraphically of all the kinds of organic remains found in the Ogallala sediments. Furthermore, distinctive floral assemblages are known as far down the geologic column as the Harrison formation of Miocene age, so that stratigraphic framing of the Ogallala sediments is possible in regions where older Cenozoic strata exist. A thorough knowledge of fossil seeds and an awareness of the distinctive qualities of the several ash falls in Neogene strata in the western plains region are necessities for anyone who pursues detailed stratigraphic studies in this part of the geologic column.
Prev Page--Fossil vertebrates and mollusks || Next Page--Stratigraphy
Kansas Geological Survey, Geology
Placed on web Aug. 4, 2011; originally published March 1956.
Comments to webadmin@kgs.ku.edu
The URL for this page is http://www.kgs.ku.edu/Publications/Bulletins/118/05_paleo.html